
SARS-CoV-2 Delta Variant (AY.3) in the Feces of a Domestic Cat


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Human Subjects
2.2. SARS-CoV-2 Clinical Testing
2.3. SARS-CoV-2 Whole Genome Sequencing
2.4. SARS-CoV-2 Variant Analysis
2.5. Phylogenetic Tree Construction
3. Results
3.1. Case Description
3.2. Molecular Detection and Sequencing
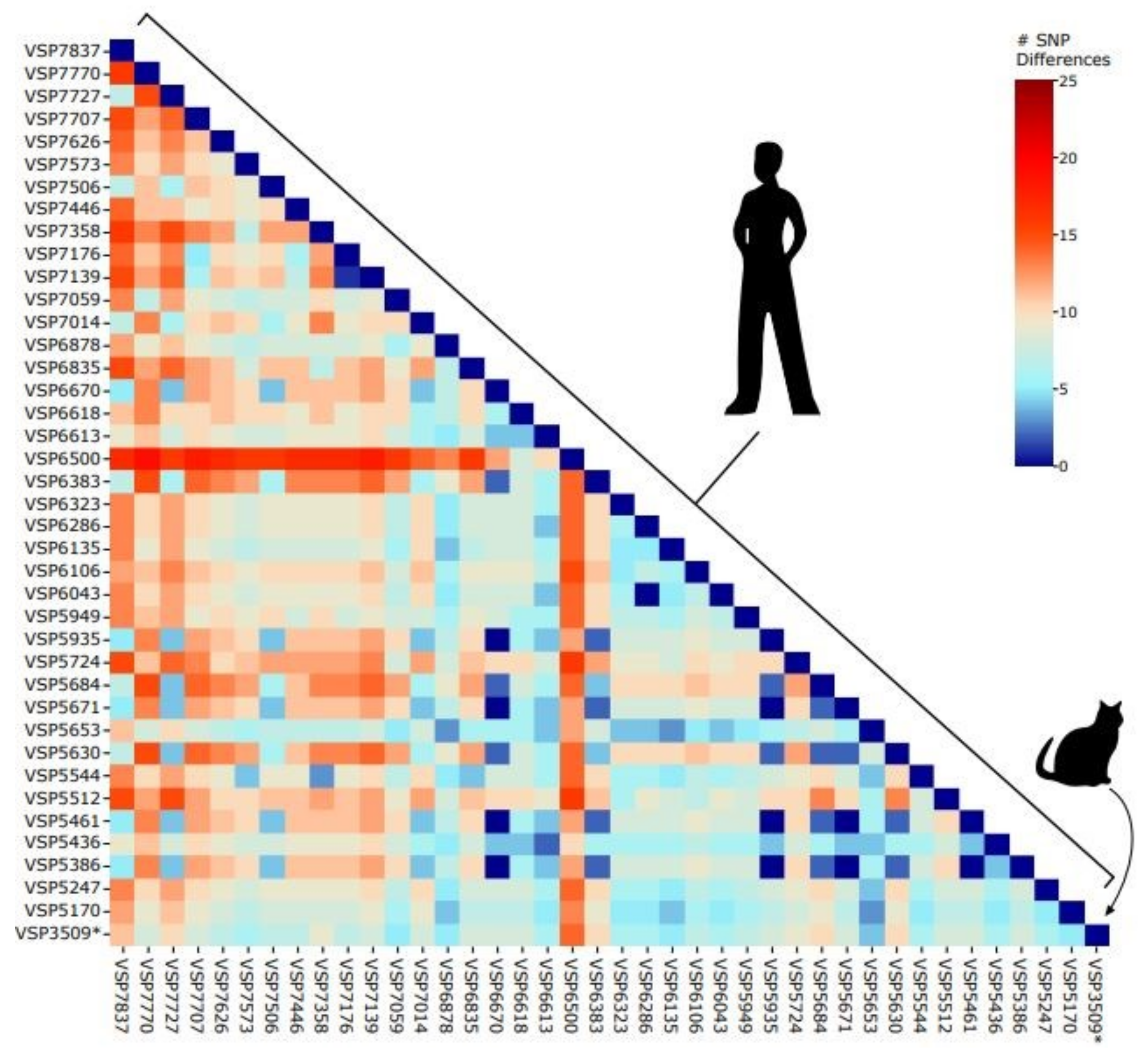
3.3. Comparison to Known Sequences in the Delaware Valley
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Kuchipudi, S.V.; Surendran-Nair, M.; Ruden, R.M.; Yon, M.; Nissly, R.H.; Nelli, R.K.; Li, L.; Jayarao, B.M.; Vandegrift, K.J.; Maranas, C.D.; et al. Multiple spillovers and onward transmission of SARS-CoV-2 in free-living and captive white-tailed deer. bioRxiv 2021. preprint. [Google Scholar] [CrossRef]
- Hamer, S.A.; Pauvolid-Corrêa, A.; Zecca, I.B.; Davila, E.; Auckland, L.D.; Roundy, C.M.; Tang, W.; Torchetti, M.K.; Killian, M.L.; Jenkins-Moore, M.; et al. SARS-CoV-2 Infections and Viral Isolations among Serially Tested Cats and Dogs in Households with Infected Owners in Texas, USA. Viruses 2021, 13, 938. [Google Scholar] [CrossRef] [PubMed]
- Oude Munnink, B.B.; Sikkema, R.S.; Nieuwenhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; van der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M.; et al. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science 2021, 371, 172–177. [Google Scholar] [CrossRef]
- Halfmann, P.J.; Hatta, M.; Chiba, S.; Maemura, T.; Fan, S.; Takeda, M.; Kinoshita, N.; Hattori, S.I.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; et al. Transmission of SARS-CoV-2 in Domestic Cats. N. Engl. J. Med. 2020, 383, 592–594. [Google Scholar] [CrossRef]
- Palmer, M.V.; Martins, M.; Falkenberg, S.; Buckley, A.; Caserta, L.C.; Mitchell, P.K.; Cassmann, E.D.; Rollins, A.; Zylich, N.C.; Renshaw, R.W.; et al. Susceptibility of white-tailed deer (Odocoileus virginianus) to SARS-CoV-2. J. Virol. 2021, 95, e00083-21. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.C.; Bevins, S.N.; Ellis, J.W.; Linder, T.J.; Tell, R.M.; Jenkins-Moore, M.; Root, J.J.; Lenoch, J.B.; Robbe-Austerman, S.; DeLiberto, T.J.; et al. SARS-CoV-2 exposure in wild white-tailed deer (Odocoileus virginianus). Proc. Natl. Acad. Sci. USA 2021, 118, e2114828118. [Google Scholar] [CrossRef]
- Fagre, A.; Lewis, J.; Eckley, M.; Zhan, S.; Rocha, S.M.; Sexton, N.R.; Burke, B.; Geiss, B.; Peersen, O.; Bass, T.; et al. SARS-CoV-2 infection, neuropathogenesis and transmission among deer mice: Implications for spillback to New World rodents. PLoS Pathog. 2021, 17, e1009585. [Google Scholar] [CrossRef]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.L.; Whitfield, M.G.; Varro, R.; et al. Community transmission and viral load kinetics of the SARS-CoV-2 delta (B.1.617.2) variant in vaccinated and unvaccinated individuals in the UK: A prospective, longitudinal, cohort study. Lancet Infect. Dis. 2021, 22, 183–195. [Google Scholar] [CrossRef]
- Arora, P.; Sidarovich, A.; Krüger, N.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.S.; Winkler, M.S.; Schulz, S.; Jäck, H.M.; et al. B.1.617.2 enters and fuses lung cells with increased efficiency and evades antibodies induced by infection and vaccination. Cell Rep. 2021, 37, 109825. [Google Scholar] [CrossRef]
- Luo, C.H.; Morris, C.P.; Sachithanandham, J.; Amadi, A.; Gaston, D.; Li, M.; Swanson, N.J.; Schwartz, M.; Klein, E.Y.; Pekosz, A.; et al. Infection with the SARS-CoV-2 Delta Variant is Associated with Higher Infectious Virus Loads Compared to the Alpha Variant in both Unvaccinated and Vaccinated Individuals. medRxiv 2021. preprint. [Google Scholar] [CrossRef]
- What We Know About the Science Center for Disease Control and Prevention. 2021. Available online: https://www.cdc.gov/coronavirus/2019-ncov/variants/delta-variant.html (accessed on 25 January 2022).
- Áine O’Toole, E.S.; Rambaut, A. PANGO Lineages: Latest Epidemiological Lineages of SARS-CoV-2: Lineage AY.3. 2021. Available online: https://cov-lineages.org/lineage.html?lineage=AY.3 (accessed on 25 January 2022).
- Doerksen, T.; Lu, A.; Noll, L.; Almes, K.; Bai, J.; Upchurch, D.; Palinski, R. Near-Complete Genome of SARS-CoV-2 Delta (AY.3) Variant Identified in a Dog in Kansas, USA. Viruses 2021, 13, 2104. [Google Scholar] [CrossRef] [PubMed]
- Karikalan, M.; Chander, V.; Mahajan, S.; Deol, P.; Agrawal, R.K.; Nandi, S.; Rai, S.K.; Mathur, A.; Pawde, A.; Singh, K.P.; et al. Natural infection of Delta mutant of SARS-CoV-2 in Asiatic lions of India. Transbound. Emerg. Dis. 2021; online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Kumar, N.; Bhatia, S.; Aasdev, A.; Kanniappan, S.; Sekhar, A.T.; Gopinadhan, A.; Silambarasan, R.; Sreekumar, C.; Dubey, C.K.; et al. SARS-CoV-2 Delta Variant among Asiatic Lions, India. Emerg. Infect. Dis. 2021, 27, 2723–2725. [Google Scholar] [CrossRef]
- Kang, K.; Chen, Q.; Gao, Y.; Yu, K.J. Detection of SARS-CoV-2 B.1.617.2 (Delta) variant in three cats owned by a confirmed COVID-19 patient in Harbin, China. Vet. Med. Sci. 2021. preprint. [Google Scholar] [CrossRef]
- Marques, A.D.; Sherrill-Mix, S.; Everett, J.; Reddy, S.; Hokama, P.; Roche, A.M.; Hwang, Y.; Glascock, A.; Whiteside, S.A.; Graham-Wooten, J.; et al. SARS-CoV-2 variants associated with vaccine breakthrough in the Delaware Valley through summer 2021. medRxiv 2021. preprint. [Google Scholar] [CrossRef] [PubMed]
- St Hilaire, B.G.; Durand, N.C.; Mitra, N.; Pulido, S.G.; Mahajan, R.; Blackburn, A.; Colaric, Z.L.; Theisen, J.W.M.; Weisz, D.; Dudchenko, O.; et al. A rapid, low cost, and highly sensitive SARS-CoV-2 diagnostic based on whole genome sequencing. bioRxiv 2020. preprint. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Li, H. A statistical framework for SNP calling.; mutation discovery.; association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Everett, J.; Hokama, P.; Roche, A.M.; Reddy, S.; Hwang, Y.; Kessler, L.; Glascock, A.; Li, Y.; Whelan, J.N.; Weiss, S.R.; et al. SARS-CoV-2 Genomic Variation in Space and Time in Hospitalized Patients in Philadelphia. mBio 2021, 12, e03456-20. [Google Scholar] [CrossRef] [PubMed]
- Ivan Aksamentov, C.R.; Hodcroft, E.B.; Neher, R.A. Nextclade: Clade assignment, mutation calling and quality control for viral genomes. J. Open Res. Softw. 2021, 6, 3773. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.J.; Long, S.W.; Christensen, P.A.; Olsen, R.J.; Olson, R.; Shukla, M.; Subedi, S.; Stevens, R.; Musser, J.M.; Pride, D.T. Analysis of the ARTIC Version 3 and Version 4 SARS-CoV-2 Primers and Their Impact on the Detection of the G142D Amino Acid Substitution in the Spike Protein. Microbiol. Spectr. 2021, 9, e01803-21. [Google Scholar] [CrossRef]
- Garigliany, M.; Van Laere, A.S.; Clercx, C.; Giet, D.; Escriou, N.; Huon, C.; van der Werf, S.; Eloit, M.; Desmecht, D. SARS-CoV-2 Natural Transmission from Human to Cat, Belgium, March 2020. Emerg. Infect. Dis. 2020, 26, 3069–3071. [Google Scholar] [CrossRef]
- Yaglom, H.D.; Hecht, G.; Goedderz, A.; Jasso-Selles, D.; Ely, J.L.; Ruberto, I.; Bowers, J.R.; Engelthaler, D.M.; Venkat, H. Genomic investigation of a household SARS-CoV-2 disease cluster in Arizona involving a cat, dog, and pet owner. One Health 2021, 13, 100333. [Google Scholar] [CrossRef]
- Wu, Y.; Guo, C.; Tang, L.; Hong, Z.; Zhou, J.; Dong, X.; Yin, H.; Xiao, Q.; Tang, Y.; Qu, X.; et al. Prolonged presence of SARS-CoV-2 viral RNA in faecal samples. Lancet Gastroenterol. Hepatol. 2020, 5, 434–435. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genomic Position | Gene | Affected Protein | Protein Mutation | Nucleotide Mutation | Reference Nucleotide | Percent in Human Dataset |
|---|---|---|---|---|---|---|
| 210 | intergenic | T | G | 40.00% | ||
| 241 | intergenic | T | C | 99.60% | ||
| 3037 | ORF1ab | nsp3 | silent | T | C | 99.95% |
| 4181 | ORF1ab | nsp3 | A1306S | T | G | 34.64% |
| 6402 | ORF1ab | nsp3 | P2046L | T | C | 34.73% |
| 7124 | ORF1ab | nsp3 | P2287S | T | C | 34.64% |
| 8140 | ORF1ab | nsp3 | silent | T | C | 1.46% |
| 8986 | ORF1ab | nsp4 | silent | T | C | 34.68% |
| 9053 | ORF1ab | nsp4 | V2930L | T | G | 34.66% |
| 9080 | ORF1ab | nsp4 | silent | T | C | 0.02% |
| 10029 | ORF1ab | nsp4 | T3255I | T | C | 38.16% |
| 11201 | ORF1ab | nsp6 | T3646A | G | A | 34.68% |
| 11332 | ORF1ab | nsp6 | silent | G | A | 34.71% |
| 11456 | ORF1ab | nsp6 | I3731V | G | A | 4.66% |
| 14408 | ORF1ab | nsp12 (RdRp) | P314L | T | C | 99.53% |
| 14520 | ORF1ab | nsp12 (RdRp) | silent | T | C | 0.00% |
| 15451 | ORF1ab | nsp12 (RdRp) | G662S | A | G | 39.36% |
| 16466 | ORF1ab | nsp13 (Hel) | P1000L | T | C | 39.32% |
| 19220 | ORF1ab | nsp14 (ExoN) | A1918V | T | C | 34.59% |
| 20744 | ORF1ab | nsp16 (2′-O-MT) | N2426T | C | A | 0.00% |
| 21618 | S | spike | T19R | G | C | 39.86% |
| 21800 | S | spike | D80N | A | G | 0.00% |
| 21987 | S | spike | G142D | A | G | 13.04% |
| 22029 | S | spike | del 6 | delAGTTCA | GAGTTCA | 39.13% |
| 22917 | S | spike | L452R | G | T | 41.58% |
| 22995 | S | spike | T478K | A | C | 40.45% |
| 23284 | S | spike | silent | C | T | 2.47% |
| 23403 | S | spike | D614G | G | A | 99.98% |
| 23604 | S | spike | P681R | G | C | 40.07% |
| 24410 | S | spike | D950N | A | G | 40.02% |
| 25339 | S | spike | silent | T | C | 2.64% |
| 25469 | ORF3a | ORF3a | S26L | T | C | 40.05% |
| 26767 | M | membrane | I82T | C | T | 41.55% |
| 27638 | ORF7a | ORF7a | V82A | C | T | 39.29% |
| 27752 | ORF7a | ORF7a | T120I | T | C | 39.60% |
| 27874 | ORF7b | ORF7b | T40I | T | C | 34.42% |
| 28248 | ORF8 | ORF8 | del 6 | delGATTTC | AGATTTC | 38.87% |
| 28271 | intergenic | del 1 | delA | TAAAA | 62.19% | |
| 28461 | N | nucleocapsid | D63G | G | A | 39.41% |
| 28881 | N | nucleocapsid | R203M | T | G | 39.91% |
| 28916 | N | nucleocapsid | G215C | T | G | 34.49% |
| 29050 | N | nucleocapsid | silent | A | G | 4.64% |
| 29402 | N | nucleocapsid | D377Y | T | G | 42.21% |
| 29509 | N | nucleocapsid | silent | T | C | 4.78% |
| 29742 | intergenic | T | G | 37.01% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lenz, O.C.; Marques, A.D.; Kelly, B.J.; Rodino, K.G.; Cole, S.D.; Perera, R.A.P.M.; Weiss, S.R.; Bushman, F.D.; Lennon, E.M. SARS-CoV-2 Delta Variant (AY.3) in the Feces of a Domestic Cat. Viruses 2022, 14, 421. https://doi.org/10.3390/v14020421
Lenz OC, Marques AD, Kelly BJ, Rodino KG, Cole SD, Perera RAPM, Weiss SR, Bushman FD, Lennon EM. SARS-CoV-2 Delta Variant (AY.3) in the Feces of a Domestic Cat. Viruses. 2022; 14(2):421. https://doi.org/10.3390/v14020421
Chicago/Turabian StyleLenz, Olivia C., Andrew D. Marques, Brendan J. Kelly, Kyle G. Rodino, Stephen D. Cole, Ranawaka A. P. M. Perera, Susan R. Weiss, Frederic D. Bushman, and Elizabeth M. Lennon. 2022. "SARS-CoV-2 Delta Variant (AY.3) in the Feces of a Domestic Cat" Viruses 14, no. 2: 421. https://doi.org/10.3390/v14020421
APA StyleLenz, O. C., Marques, A. D., Kelly, B. J., Rodino, K. G., Cole, S. D., Perera, R. A. P. M., Weiss, S. R., Bushman, F. D., & Lennon, E. M. (2022). SARS-CoV-2 Delta Variant (AY.3) in the Feces of a Domestic Cat. Viruses, 14(2), 421. https://doi.org/10.3390/v14020421