
Antibodies to Crucial Epitopes on HSV-2 Glycoprotein D as a Guide to Dosing an mRNA Genital Herpes Vaccine

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
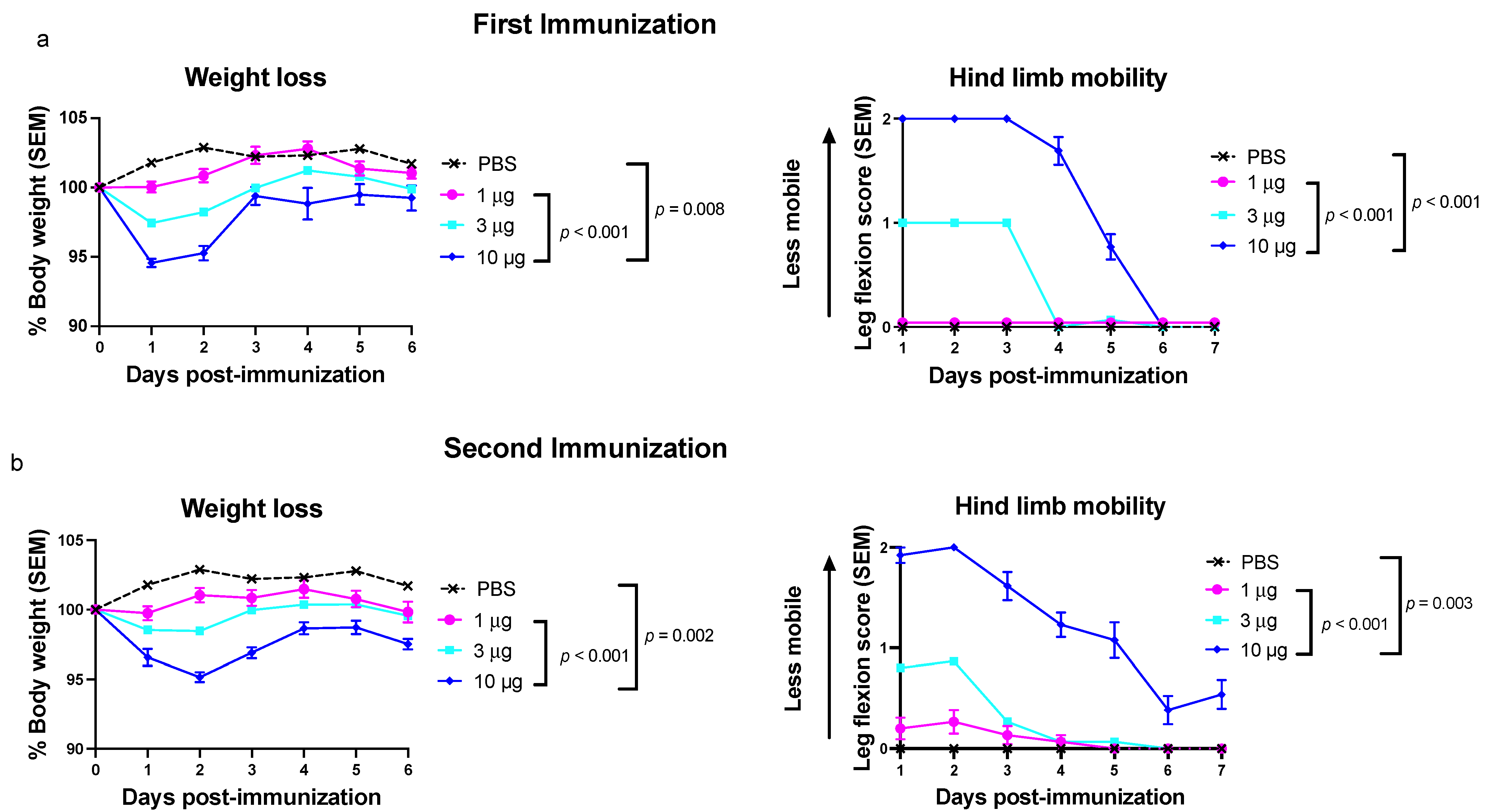
2.1. Evaluating Trivalent mRNA-LNP Vaccine Toxicity
2.2. Immunizing with gD2 mRNA-LNP
2.3. Serum IgG ELISA and Neutralizing Antibody Titers
2.4. Biosensor-Based Antibody Competition Assay
2.5. HSV-2 Vaginal Infection and Vaccine Efficacy
2.6. Vaginal Virus Titers
2.7. qPCR for HSV-2 DNA Present in the DRG
2.8. Statistical Analysis
2.9. Study Approval
3. Results
3.1. Experimental Design and Rationale
3.2. Experiment 1: Toxicity Is Dose Dependent for the Trivalent mRNA-LNP Vaccine
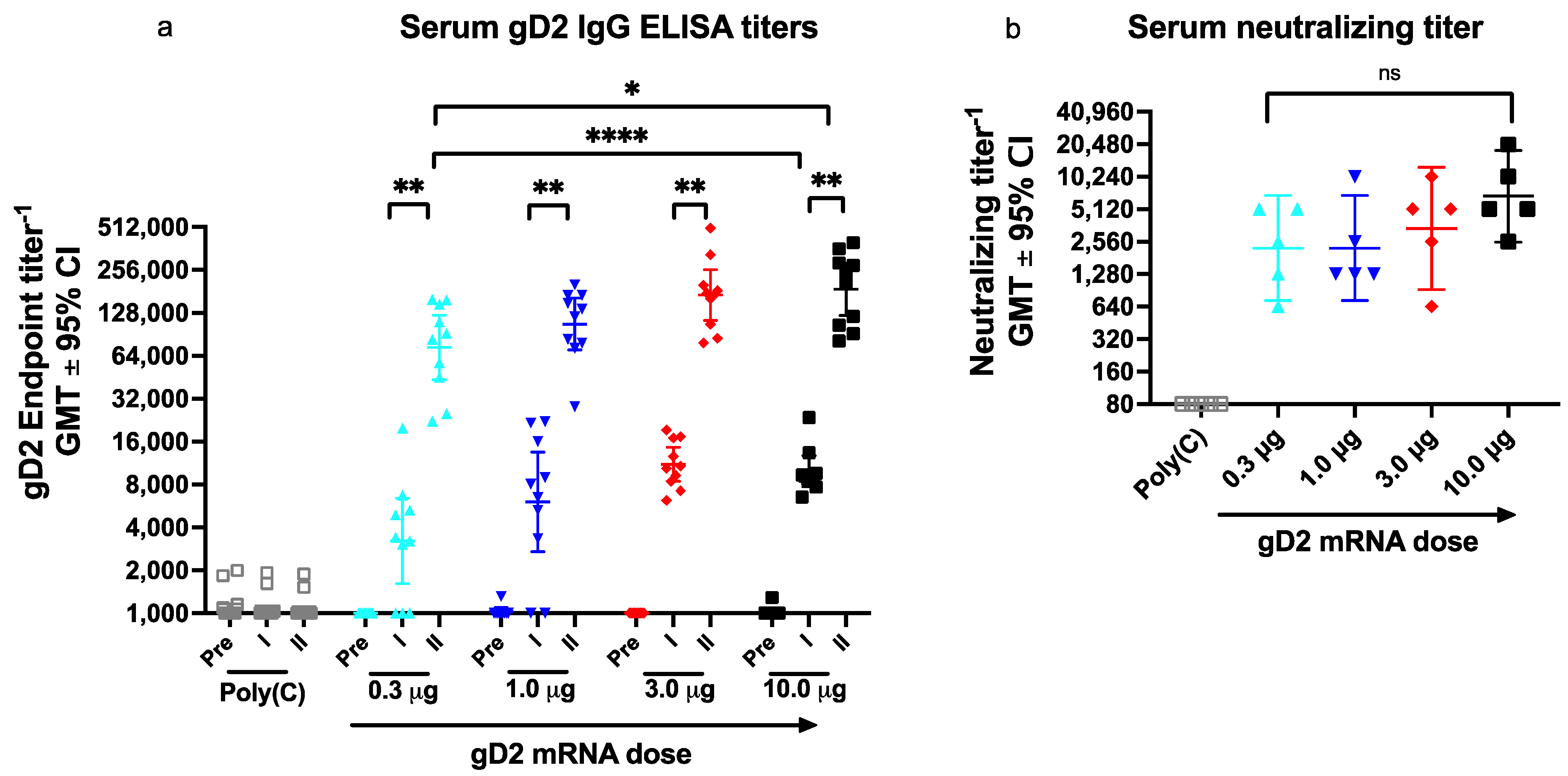
3.3. Experiment 2: Dose Escalation Study of gD2 mRNA to Evaluate IgG ELISA Titers, Neutralizing Antibodies, and Antibodies to Crucial gD2 Epitopes
3.3.1. Serum IgG ELISA Titers
3.3.2. Serum Neutralizing Antibody Titers
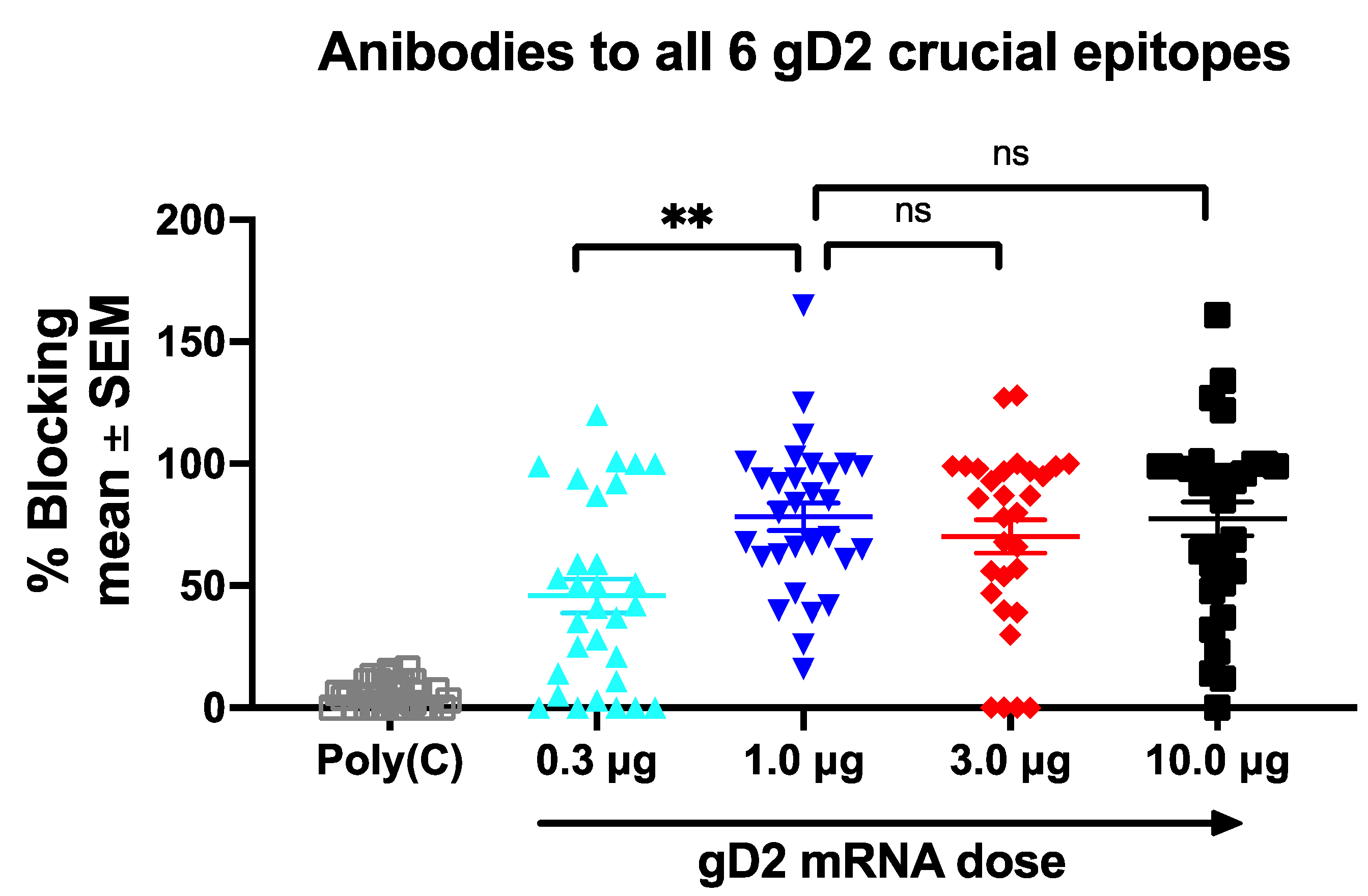
3.3.3. Antibodies to Crucial gD2 Epitopes
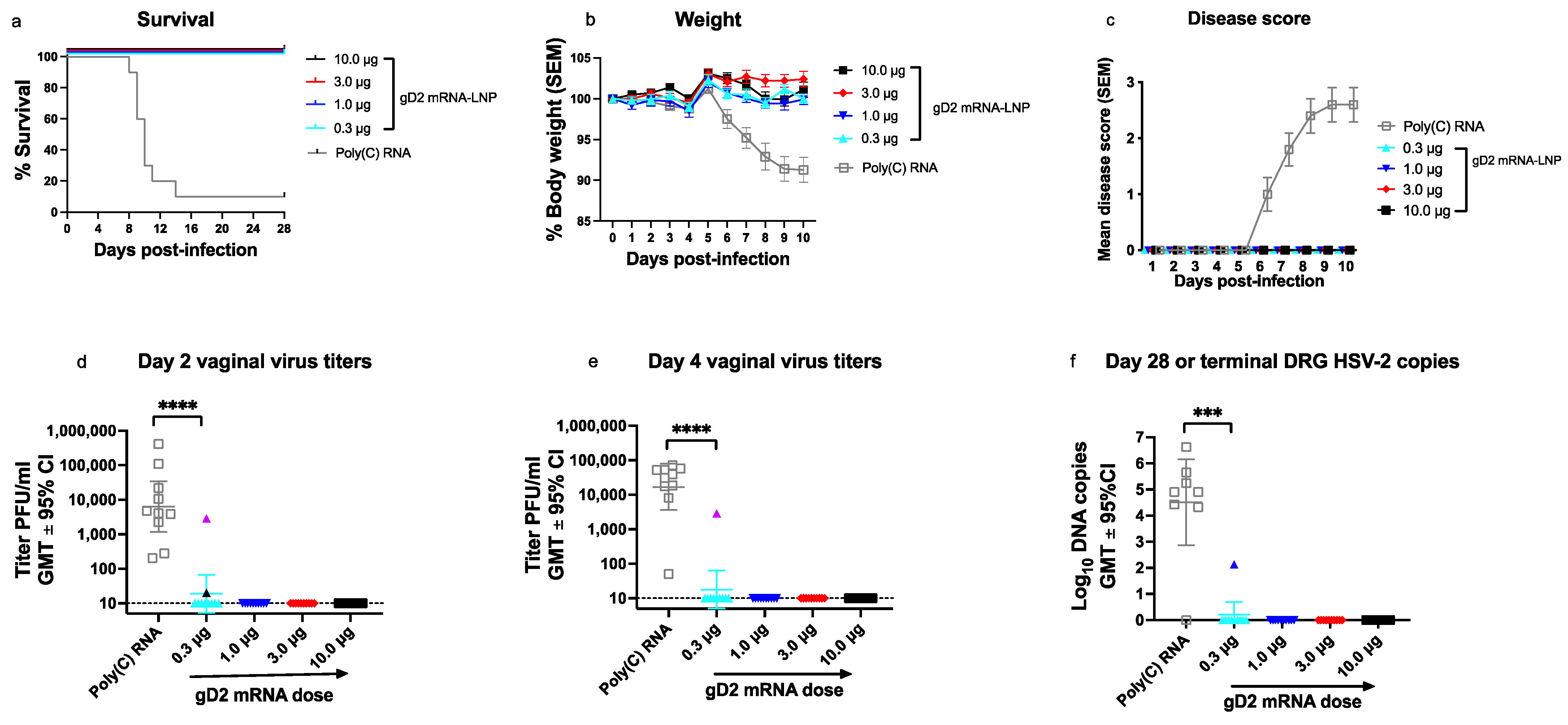
3.4. Experiment 3: Vaccine Efficacy in gD2 mRNA Immunized Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, M.G.; Burgess, J.L.; Naleway, A.L.; Tyner, H.; Yoon, S.K.; Meece, J.; Olsho, L.E.W.; Caban-Martinez, A.J.; Fowlkes, A.L.; Lutrick, K.; et al. Prevention and Attenuation of Covid-19 with the BNT162b2 and mRNA-1273 Vaccines. N. Engl. J. Med. 2021, 385, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.A.; Fuhr, R.; Smolenov, I.; Ribeiro, A.; Panther, L.; Watson, M.; Senn, J.J.; Smith, M.; Almarsson, Ö.; Pujar, H.S.; et al. mRNA Vaccines against H10N8 and H7N9 Influenza Viruses of Pandemic Potential Are Immunogenic and Well Tolerated in Healthy Adults in Phase 1 Randomized Clinical Trials. Vaccine 2019, 37, 3326–3334. [Google Scholar] [CrossRef] [PubMed]
- Alberer, M.; Gnad-Vogt, U.; Hong, H.S.; Mehr, K.T.; Backert, L.; Finak, G.; Gottardo, R.; Bica, M.A.; Garofano, A.; Koch, S.D.; et al. Safety Immunogenicity of a mRNA Rabies Vaccine in Healthy Adults: An Open-Label, Non-Randomised, Prospective, First-in-Human Phase 1 Clinical Trial. Lancet 2017, 390, 1511–1520. [Google Scholar] [CrossRef]
- Awasthi, S.; Knox, J.J.; Desmond, A.; Alameh, M.G.; Gaudette, B.T.; Lubinski, J.M.; Naughton, A.; Hook, L.M.; Egan, K.P.; Tam, Y.K.; et al. Trivalent Nucleoside-Modified mRNA Vaccine Yields Durable Memory B Cell Protection against Genital Herpes in Preclinical Models. J. Clin. Investig. 2021, 131, e152310. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Hook, L.M.; Pardi, N.; Wang, F.; Myles, A.; Cancro, M.P.; Cohen, G.H.; Weissman, D.; Friedman, H.M. Nucleoside-Modified mRNA Encoding HSV-2 Glycoproteins C, D, and E Prevents Clinical and Subclinical Genital Herpes. Sci. Immunol. 2019, 4, eaaw7083. [Google Scholar] [CrossRef] [PubMed]
- Egan, K.P.; Hook, L.M.; Naughton, A.; Pardi, N.; Awasthi, S.; Cohen, G.H.; Weissman, D.; Friedman, H.M. An HSV-2 Nucleoside-Modified mRNA Genital Herpes Vaccine Containing Glycoproteins gC, gD, and gE Protects Mice against HSV-1 Genital Lesions and Latent Infection. PLoS Pathog. 2020, 16, e1008795. [Google Scholar] [CrossRef] [PubMed]
- LaTourette, P.C., 2nd; Awasthi, S.; Desmond, A.; Pardi, N.; Cohen, G.H.; Weissman, D.; Friedman, H.M. Protection against Herpes Simplex Virus Type 2 Infection in a Neonatal Murine Model Using a Trivalent Nucleoside-Modified mRNA in Lipid Nanoparticle Vaccine. Vaccine 2020, 38, 7409–7413. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II Study of COVID-19 RNA Vaccine BNT162b1 in Adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Estanbouli, H.; Bouteau, A.; Igyártó, B.Z. The mRNA-LNP Platform’s Lipid Nanoparticle Component Used in Preclinical Vaccine Studies Is Highly Inflammatory. Iscience 2021, 24, 103479. [Google Scholar] [CrossRef] [PubMed]
- Hook, L.M.; Cairns, T.M.; Awasthi, S.; Brooks, B.D.; Ditto, N.T.; Eisenberg, R.J.; Cohen, G.H.; Friedman, H.M. Vaccine-Induced Antibodies to Herpes Simplex Virus Glycoprotein D Epitopes Involved in Virus Entry and Cell-to-Cell Spread Correlate with Protection against Genital Disease in Guinea Pigs. PLoS Pathog 2018, 14, e1007095. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Hook, L.M.; Swaminathan, G.; Cairns, T.M.; Brooks, B.; Smith, J.S.; Ditto, N.T.; Gindy, M.E.; Bett, A.J.; Espeseth, A.S.; et al. Antibody Responses to Crucial Functional Epitopes as a Novel Approach to Assess Immunogenicity of Vaccine Adjuvants. Vaccine 2019, 37, 3770–3778. [Google Scholar] [CrossRef] [PubMed]
- Whitbeck, J.C.; Huang, Z.Y.; Cairns, T.M.; Gallagher, J.R.; Lou, H.; Ponce-de-Leon, M.; Belshe, R.B.; Eisenberg, R.J.; Cohen, G.H. Repertoire of Epitopes Recognized by Serum IgG from Humans Vaccinated with Herpes Simplex Virus 2 Glycoprotein D. J. Virol. 2014, 88, 7786–7795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awasthi, S.; Hook, L.M.; Shaw, C.E.; Friedman, H.M. A Trivalent Subunit Antigen Glycoprotein Vaccine as Immunotherapy for Genital Herpes in the Guinea Pig Genital Infection Model. Hum. Vaccines Immunother. 2017, 13, 2785–2793. [Google Scholar] [CrossRef] [PubMed]
- Cairns, T.M.; Ditto, N.T.; Lou, H.; Brooks, B.D.; Atanasiu, D.; Eisenberg, R.J.; Cohen, G.H. Global Sensing of the Antigenic Structure of Herpes Simplex Virus gD Using High-Throughput Array-Based SPR Imaging. PLoS Pathog 2017, 13, e1006430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushic, C.; Ashkar, A.A.; Reid, L.A.; Rosenthal, K.L. Progesterone Increases Susceptibility and Decreases Immune Responses to Genital Herpes Infection. J. Virol. 2003, 77, 4558–4565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, T.M.; Ditto, N.T.; Atanasiu, D.; Lou, H.; Brooks, B.D.; Saw, W.T.; Eisenberg, R.J.; Cohen, G.H. Surface Plasmon Resonance (SPR) Reveals Direct Binding of HSV Glycoproteins gH/gL to gD and Locates a gH/gL Binding Site on gD. J. Virol. 2019, 93, e00289-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanasiu, D.; Saw, W.T.; Cairns, T.M.; Eisenberg, R.J.; Cohen, G.H. Using Split Luciferase Assay and Anti-HSV Glycoprotein Monoclonal Antibodies to Predict a Functional Binding Site between gD and gH/gL. J. Virol. 2021, 95, e00053-21. [Google Scholar] [CrossRef] [PubMed]
- Lazear, E.; Whitbeck, J.C.; Ponce-de-Leon, M.; Cairns, T.M.; Willis, S.H.; Zuo, Y.; Krummenacher, C.; Cohen, G.H.; Eisenberg, R.J. Antibody-Induced Conformational Changes in Herpes Simplex Virus Glycoprotein gD Reveal New Targets for Virus Neutralization. J. Virol. 2012, 86, 1563–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toh, Z.Q.; Kosasih, J.; Russell, F.M.; Garland, S.M.; Mulholland, E.K.; Licciardi, P.V. Recombinant Human Papillomavirus Nonavalent Vaccine in the Prevention of Cancers Caused by Human Papillomavirus. Infect. Drug Resist. 2019, 12, 1951–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hook, L.M.; Awasthi, S.; Cairns, T.M.; Alameh, M.-G.; Fowler, B.T.; Egan, K.P.; Sung, M.M.H.; Weissman, D.; Cohen, G.H.; Friedman, H.M. Antibodies to Crucial Epitopes on HSV-2 Glycoprotein D as a Guide to Dosing an mRNA Genital Herpes Vaccine. Viruses 2022, 14, 540. https://doi.org/10.3390/v14030540
Hook LM, Awasthi S, Cairns TM, Alameh M-G, Fowler BT, Egan KP, Sung MMH, Weissman D, Cohen GH, Friedman HM. Antibodies to Crucial Epitopes on HSV-2 Glycoprotein D as a Guide to Dosing an mRNA Genital Herpes Vaccine. Viruses. 2022; 14(3):540. https://doi.org/10.3390/v14030540
Chicago/Turabian StyleHook, Lauren M., Sita Awasthi, Tina M. Cairns, Mohamad-Gabriel Alameh, Bernard T. Fowler, Kevin P. Egan, Molly M. H. Sung, Drew Weissman, Gary H. Cohen, and Harvey M. Friedman. 2022. "Antibodies to Crucial Epitopes on HSV-2 Glycoprotein D as a Guide to Dosing an mRNA Genital Herpes Vaccine" Viruses 14, no. 3: 540. https://doi.org/10.3390/v14030540
APA StyleHook, L. M., Awasthi, S., Cairns, T. M., Alameh, M. -G., Fowler, B. T., Egan, K. P., Sung, M. M. H., Weissman, D., Cohen, G. H., & Friedman, H. M. (2022). Antibodies to Crucial Epitopes on HSV-2 Glycoprotein D as a Guide to Dosing an mRNA Genital Herpes Vaccine. Viruses, 14(3), 540. https://doi.org/10.3390/v14030540