
Two Novel Lytic Bacteriophages Infecting Enterococcus spp. Are Promising Candidates for Targeted Antibacterial Therapy

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteriophage Isolation and Preparation
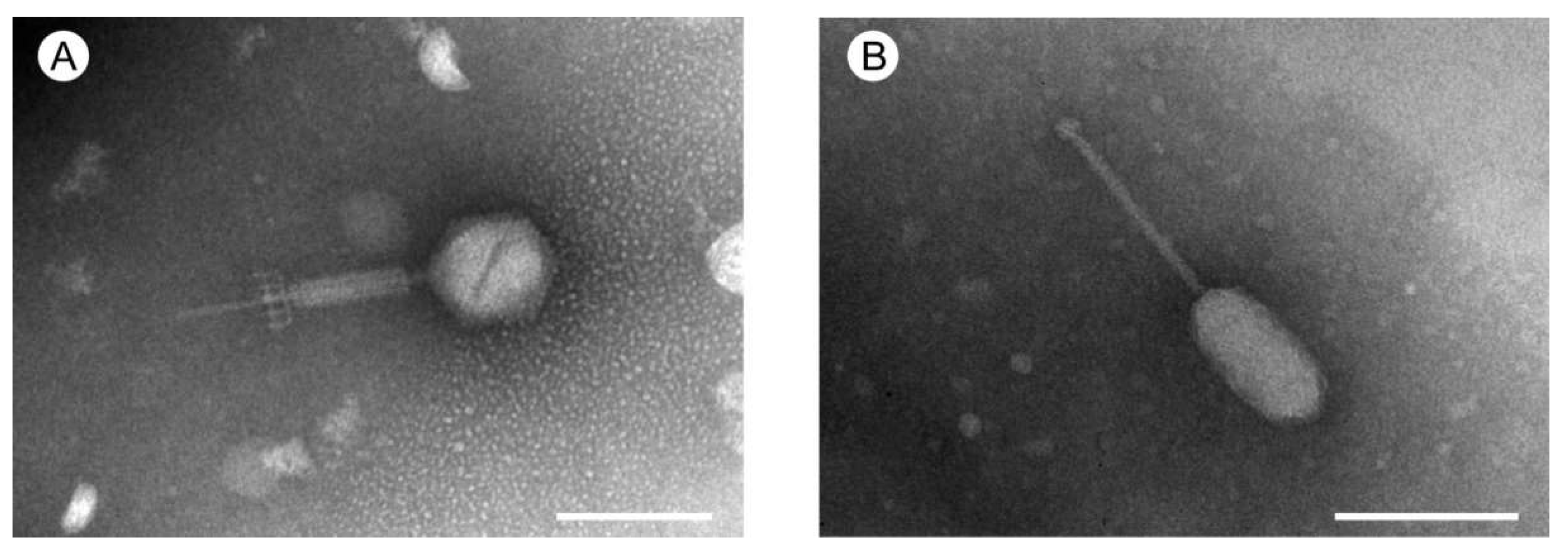
2.2. Transmission Electron Microscopy
2.3. In Vitro Efficacy
2.4. Determining the Host Range of Viruses
2.5. DNA Isolation and Whole-Genome Sequencing
2.6. Bioinformatic Analysis
2.7. Peritoneal Infection Model in Mice
3. Results and Discussion
3.1. Isolation and Identification of Bacteriophages
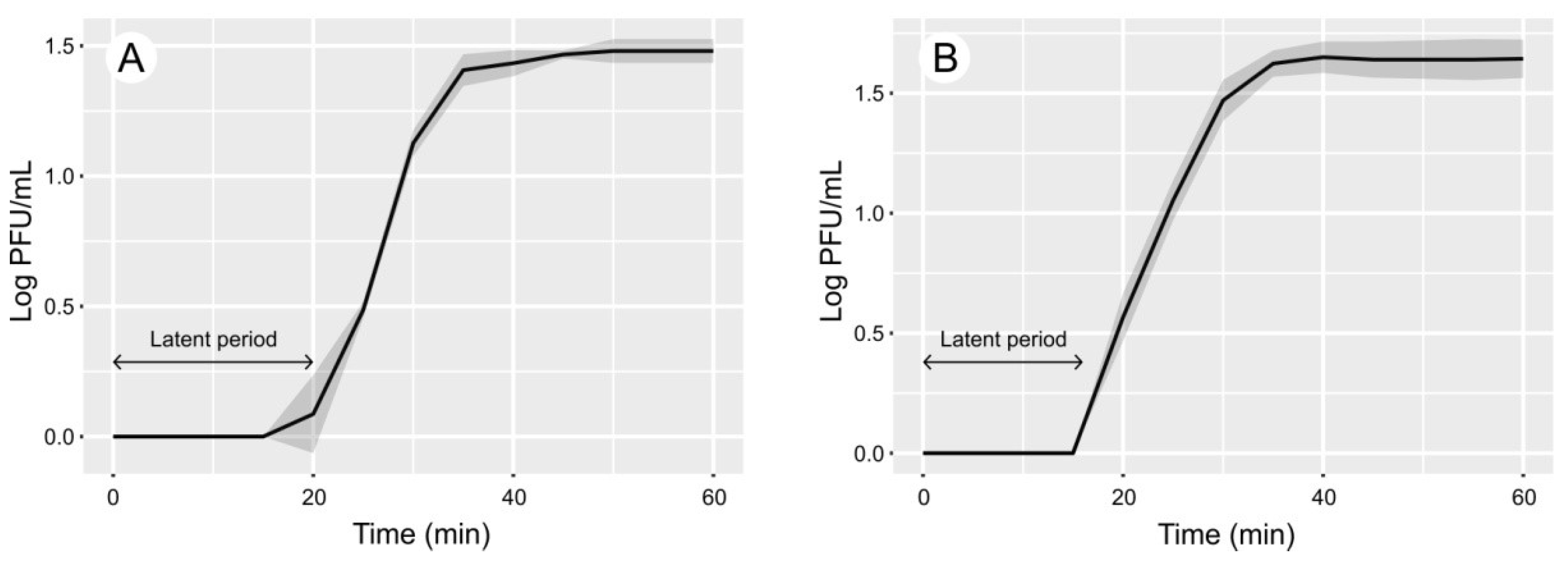
3.2. Phage Life Cycle and Morphology
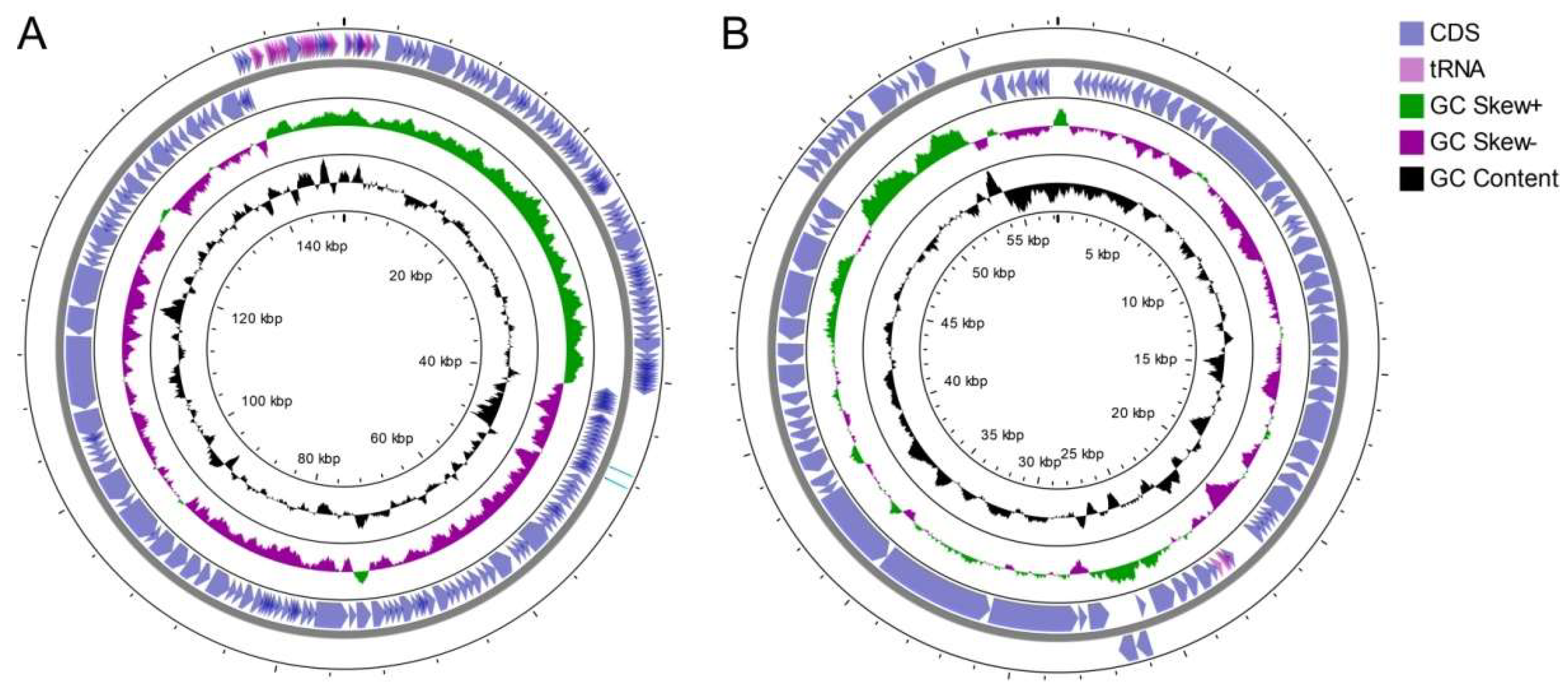
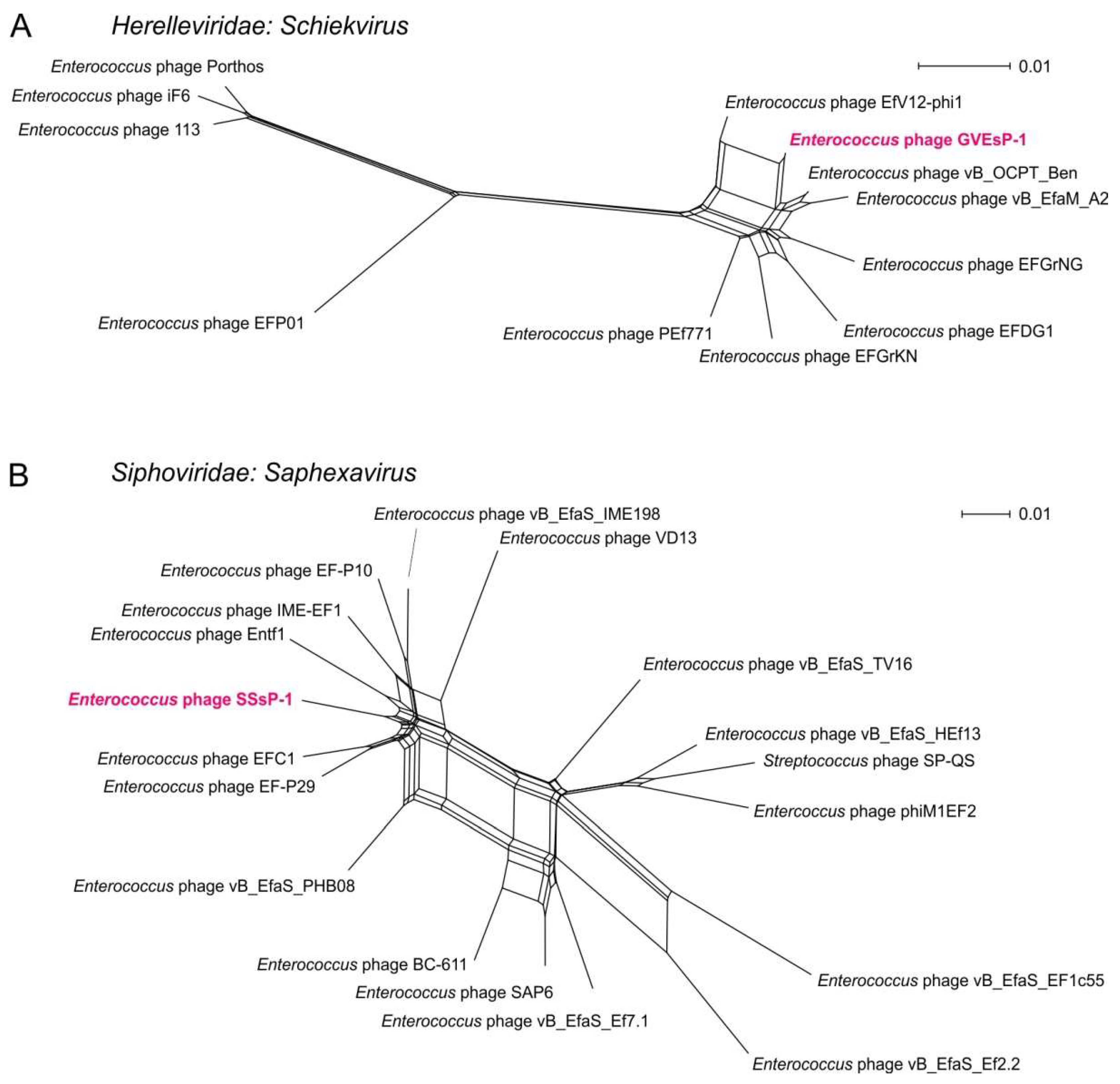
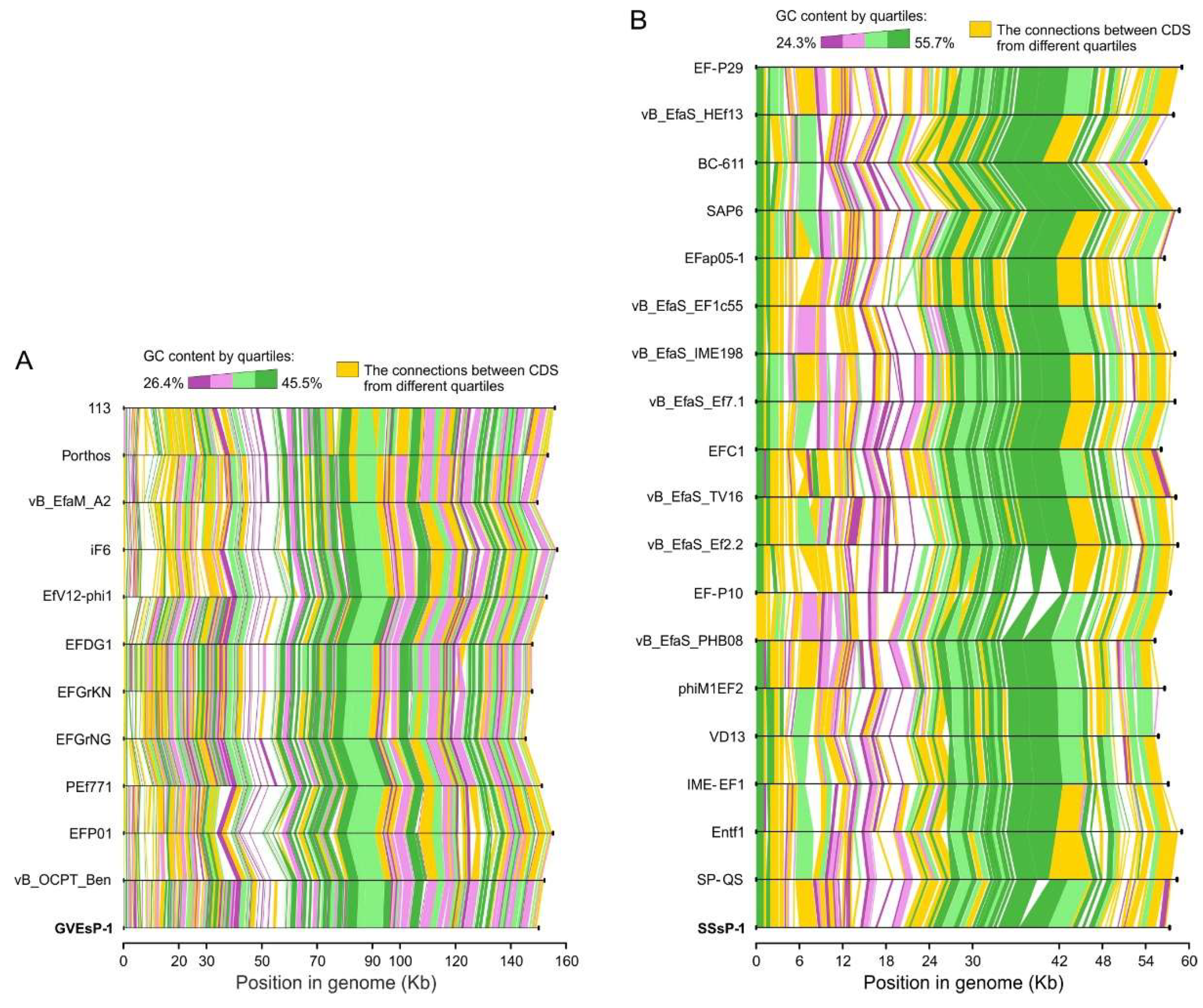
3.3. Genome Structure and Conservation
3.4. Potential Anti-CRISPR Loci
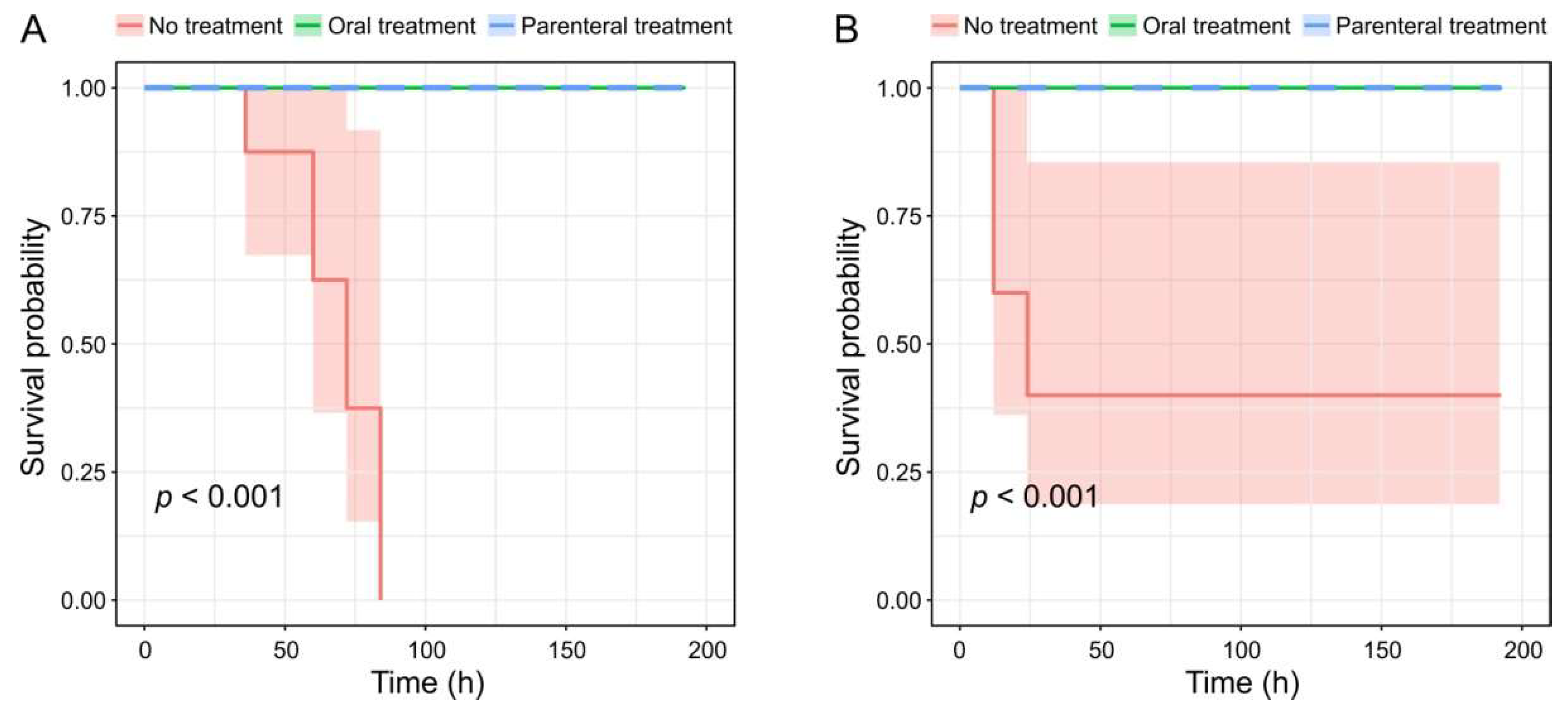
3.5. Mouse Infection Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial resistance in ESKAPE pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Gao, W.; Howden, B.P.; Stinear, T.P. Evolution of virulence in Enterococcus faecium, a hospital-adapted opportunistic pathogen. Curr. Opin. Microbiol. 2018, 41, 76–82. [Google Scholar] [CrossRef]
- Fraser, J.L.; Mwatondo, A.; Alimi, Y.H.; Varma, J.K.; Vilas, V.J.D.R. Healthcare-associated outbreaks of bacterial infections in Africa, 2009-2018: A review. Int. J. Infect. Dis. 2021, 103, 469–477. [Google Scholar] [CrossRef]
- Kirkizlar, T.A.; Akalin, H.; Kirkizlar, O.; Ozkalemkas, F.; Ozkocaman, V.; Kazak, E.; Ozakin, C.; Bulbul, E.N.; Ozboz, E.S.; Ali, R. Vancomycin-resistant enterococci infection and predisposing factors for infection and mortality in patients with acute leukaemia and febrile neutropenia. Leuk. Res. 2020, 99, 106463. [Google Scholar] [CrossRef]
- Serra-Burriel, M.; Keys, M.; Campillo-Artero, C.; Agodi, A.; Barchitta, M.; Gikas, A.; Palos, C.; López-Casasnovas, G. Impact of multi-drug resistant bacteria on economic and clinical outcomes of healthcare-associated infections in adults: Systematic review and meta-analysis. PLoS ONE. 2020, 15, e0227139. [Google Scholar] [CrossRef]
- Gilmore, M.S.; Lebreton, F.; van Schaik, W. Genomic transition of enterococci from gut commensals to leading causes of multidrug-resistant hospital infection in the antibiotic era. Curr. Opin. Microbiol. 2013, 16, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Zhen, X.; Lundborg, C.S.; Sun, X.; Hu, X.; Dong, H. Economic burden of antibiotic resistance in ESKAPE organisms: A systematic review. Antimicrob. Resist. Infect. Control. 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet. Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Baëtz, B.; Boudrioua, A.; Hartke, A.; Giraud, C. Alternatives to fight vancomycin-resistant staphylococci and enterococci. Antibiotics 2021, 10, 1116. [Google Scholar] [CrossRef]
- McCallin, S.; Brüssow, H. Clinical Trials of Bacteriophage Therapeutics. In Bacteriophages: Biology, Technology, Therapy; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1099–1127. [Google Scholar] [CrossRef]
- Neuts, A.S.; Berkhout, H.J.; Hartog, A.; Goosen, J.H.M. Bacteriophage therapy cures a recurrent Enterococcus faecalis infected total hip arthroplasty? A case report. Acta Orthop. 2021, 20, 678–680. [Google Scholar] [CrossRef]
- Paul, K.; Merabishvili, M.; Hazan, R.; Christner, M.; Herden, U.; Gelman, D.; Khalifa, L.; Yerushalmy, O.; Coppenhagen-Glazer, S.; Harbauer, T.; et al. Bacteriophage rescue therapy of a vancomycin-resistant Enterococcus faecium infection in a one-year-old child following a third liver transplantation. Viruses 2021, 13, 1785. [Google Scholar] [CrossRef]
- Rubalskii, E.; Ruemke, S.; Salmoukas, C.; Boyle, E.C.; Warnecke, G.; Tudorache, I.; Shrestha, M.; Schmitto, J.D.; Martens, A.; Rojas, S.V.; et al. Bacteriophage therapy for critical infections related to cardiothoracic surgery. Antibiotics 2020, 9, 232. [Google Scholar] [CrossRef]
- Khalifa, L.; Shlezinger, M.; Beyth, S.; Houri-Haddad, Y.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Phage therapy against Enterococcus faecalis in dental root canals. J. Oral Microbiol. 2016, 8, 32157. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Johnson, C.N.; Luong, P.; Hullahalli, K.; McBride, S.W.; Schubert, A.M.; Palmer, K.L.; Carlson, P.E., Jr.; Duerkop, B.A. Bacteriophage resistance alters antibiotic-mediated intestinal expansion of enterococci. Infect. Immun. 2019, 87, e00085-19. [Google Scholar] [CrossRef] [Green Version]
- Bhandare, S.; Goodridge, L. Bacteriophages as bio-sanitizers in food production and healthcare settings. In Bacteriophages: Biology, Technology, Therapy; Springer: Berlin/Heidelberg, Germany, 2021; pp. 769–788. [Google Scholar] [CrossRef]
- Taati Moghadam, M.; Amirmozafari, N.; Shariati, A.; Hallajzadeh, M.; Mirkalantari, S.; Khoshbayan, A.; Masjedian Jazi, F. How phages overcome the challenges of drug resistant bacteria in clinical infections. Infect. Drug Resist. 2020, 13, 45–61. [Google Scholar] [CrossRef] [Green Version]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.H.; Siak, J.S.; Gray, R.H. Characteristics of PRD1, a plasmid-dependent broad host range DNA bacteriophage. J. Virol. 1974, 14, 689–699. [Google Scholar] [CrossRef] [Green Version]
- Pirnay, J.P.; Blasdel, B.G.; Bretaudeau, L.; Buckling, A.; Chanishvili, N.; Clark, J.R.; Corte-Real, S.; Debarbieux, L.; Dublanchet, A.; De Vos, D.; et al. Quality and safety requirements for sustainable phage therapy products. Pharm. Res. 2015, 32, 2173–2179. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.H. Bacteriophages; Interscience Publishers: New York, NY, USA; London, UK, 1959. [Google Scholar]
- Kutter, E.; Sulakvelidze, A. Bacteriophages: Biology and Applications; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Sullivan, M. Calculating Burst Size. Available online: https://www.protocols.io/view/Calculating-burst-size-3byl47erlo5d/v1.dx.doi.org/10.17504/protocols.io.ddm245 (accessed on 8 April 2022).
- Maniatis, T.; Fritsch, E.F.; Sambrook, J. Molecular Cloning. A Laboratory Manual; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1982. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Farrer, R.A. Synima: A synteny imaging tool for annotated genome assemblies. BMC Bioinformatics 2017, 18, 507. [Google Scholar] [CrossRef]
- Yi, H.; Huang, L.; Yang, B.; Gomez, J.; Zhang, H.; Yin, Y. AcrFinder: Genome mining anti-CRISPR operons in prokaryotes and their viruses. Nucleic Acids Res. 2020, 48, W358–W365. [Google Scholar] [CrossRef]
- Seemann, T. ABRicate. Available online: https://github.com/tseemann/abricate (accessed on 27 December 2021).
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 8 November 2021).
- Therneau, T. A Package for Survival Analysis in R. R Package Version 3.3-1. 2022. Available online: https://CRAN.R-project.org/package=survival (accessed on 7 April 2022).
- Kassambara, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using ‘ggplot2’. R Package Version 0.4.9. 2021. Available online: https://CRAN.R-project.org/package=survminer (accessed on 7 April 2022).
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A roadmap for genome-based phage taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef]
- Rigvava, S.; Tchgkonia, I.; Jgenti, D.; Dvalidze, T.; Carpino, J.; Goderdzishvili, M. Comparative analysis of the biological and physical properties of Enterococcus faecalis bacteriophage vB_EfaS_GEC-EfS_3 and Streptococcus mitis bacteriophage vB_SmM_GEC-SmitisM_2. Can. J. Microbiol. 2013, 59, 18–21. [Google Scholar] [CrossRef]
- Zhang, W.; Mi, Z.; Yin, X.; Fan, H.; An, X.; Zhang, Z.; Chen, J.; Tong, Y. Characterization of Enterococcus faecalis phage IME-EF1 and its endolysin. PLoS ONE 2013, 8, e80435. [Google Scholar] [CrossRef]
- Eladero, V.; Egomez-Sordo, C.; Esanchez-Llana, E.; del Rio, B.; Redruello, B.; Efernandez, M.; Martin, M.C.; Alvarez, M.A. Q69 (an E. faecalis-infecting bacteriophage) as a biocontrol agent for reducing tyramine in dairy products. Front. Microbiol. 2016, 7, 445. [Google Scholar] [CrossRef]
- Cheng, M.; Liang, J.; Zhang, Y.; Hu, L.; Gong, P.; Cai, R.; Zhang, L.; Zhang, H.; Ge, J.; Ji, Y.; et al. The Bacteriophage EF-P29 efficiently protects against lethal vancomycin-resistant Enterococcus faecalis and alleviates gut microbiota imbalance in a murine bacteremia model. Front. Microbiol. 2017, 8, 837. [Google Scholar] [CrossRef]
- Rahmat Ullah, S.; Andleeb, S.; Raza, T.; Jamal, M.; Mehmood, K. Effectiveness of a lytic phage SRG1 against vancomycin-resistant Enterococcus faecalis in compost and soil. Biomed. Res. Int. 2017, 2017, 9351017. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Xing, S.; Zhao, F.; Li, P.; Mi, Z.; Shi, T.; Liu, H.; Tong, Y. Characterization and genome analysis of novel phage vB_EfaP_IME195 infecting Enterococcus faecalis. Virus Genes 2018, 54, 804–811. [Google Scholar] [CrossRef]
- Lee, D.; Im, J.; Na, H.; Ryu, S.; Yun, C.H.; Han, S.H. The novel Enterococcus phage vB_EfaS_HEf13 has broad lytic activity against clinical isolates of Enterococcus faecalis. Front Microbiol. 2019, 10, 2877. [Google Scholar] [CrossRef]
- Lancefield, R.C. A serological differentiation of human and other groups of hemolytic streptococci. J. Exp. Med. 1933, 57, 571–595. [Google Scholar] [CrossRef]
- Fiore, E.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of enterococci. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Barylski, J.; Kropinski, A.M.; Alikhan, N.F.; Adriaenssens, E.M.; Ictv Report Consortium. ICTV virus taxonomy profile: Herelleviridae. J. Gen. Virol. 2020, 101, 362–363. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Y.; Sun, E.; Hua, L.; Peng, Z.; Wu, B. Characterization of a lytic bacteriophage vB_EfaS_PHB08 harboring endolysin Lys08 against Enterococcus faecalis biofilms. Microorganisms 2020, 8, 1332. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, N. Complete genome sequence of bacteriophage EFC1 infecting Enterococcus faecalis from chicken. 2021. preprint. Available online: https://doi.org/10.21203/rs.3.rs-1118970/v1 (accessed on 22 December 2021).
- Xiang, Y.; Li, W.; Song, F.; Yang, X.; Zhou, J.; Yu, H.; Ji, X.; Wei, Y. Biological characteristics and whole-genome analysis of the Enterococcus faecalis phage PEf771. Can. J. Microbiol. 2020, 66, 505–520. [Google Scholar] [CrossRef]
- Khalifa, L.; Brosh, Y.; Gelman, D.; Coppenhagen-Glazer, S.; Beyth, S.; Poradosu-Cohen, R.; Que, Y.A.; Beyth, N.; Hazan, R. Targeting Enterococcus faecalis biofilms with phage therapy. Appl. Environ. Microbiol. 2015, 81, 2696–2705. [Google Scholar] [CrossRef] [Green Version]
- Lima-Mendez, G.; Van Helden, J.; Toussaint, A.; Leplae, R. Reticulate representation of evolutionary and functional relationships between phage genomes. Mol. Biol. Evol. 2008, 25, 762–777. [Google Scholar] [CrossRef] [Green Version]
- Casjens, S.R. Comparative genomics and evolution of the tailed-bacteriophages. Curr. Opin. Microbiol. 2005, 8, 451–458. [Google Scholar] [CrossRef]
- Maxwell, K.L. The anti-CRISPR story: A battle for survival. Mol. Cell. 2017, 68, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Bondy-Denomy, J.; Pawluk, A.; Maxwell, K.L.; Davidson, A.R. Bacteriophage genes that inactivate the CRISPR/Cas bacterial immune system. Nature 2013, 493, 429–432. [Google Scholar] [CrossRef] [Green Version]
- Malone, L.M.; Warring, S.L.; Jackson, S.A.; Warnecke, C.; Gardner, P.P.; Gumy, L.F.; Fineran, P.C. A jumbo phage that forms a nucleus-like structure evades CRISPR–Cas DNA targeting but is vulnerable to type III RNA-based immunity. Nat. Microbiol. 2020, 5, 48–55. [Google Scholar] [CrossRef]
- Harrington, L.B.; Doxzen, K.W.; Ma, E.; Liu, J.J.; Knott, G.J.; Edraki, A.; Garcia, B.; Amrani, N.; Chen, J.S.; Cofsky, J.C.; et al. A broad-spectrum inhibitor of CRISPR-Cas9. Cell 2017, 170, 1224–1233.e15. [Google Scholar] [CrossRef] [Green Version]
- Lyons, C.; Raustad, N.; Bustos, M.A.; Shiaris, M. Incidence of Type II CRISPR1-Cas systems in Enterococcus is species-dependent. PLoS ONE 2015, 10, e0143544. [Google Scholar] [CrossRef] [Green Version]
- Goh, H.M.S.; Yong, M.H.A.; Chong, K.K.L.; Kline, K.A. Model systems for the study of enterococcal colonization and infection. Virulence. 2017, 8, 1525–1562. [Google Scholar] [CrossRef] [Green Version]
- Olsen, N.S.; Nielsen, K.J.; Plöger, M.; Kot, W.; Hansen, L.H. Enterococcus phage Nonaheksakonda infecting clinical isolates of Enterococcus faecalis represents a new lineage in the family Siphoviridae. Arch. Virol. 2021, 166, 593–599. [Google Scholar] [CrossRef]
- Dąbrowska, K. Phage therapy: What factors shape phage pharmacokinetics and bioavailability? Systematic and critical review. Med. Res. Rev. 2019, 39, 2000–2025. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Source |
|---|---|
| Serg | Urine of patient with urinary tract infection, Saint Petersburg, Russia |
| 5arctic | Ornithogenic soil associated with Rissa tridactyla, Svalbard |
| ATCC 29212 | Swedish Institute for Infectious Disease Control (SMI) |
| CCUG 52538 | Swedish Institute for Infectious Disease Control (SMI) |
| Phage | Putative Anti-CRISPR Locus | Strand | Number of ORFs | Start | End | Known ORF Annotations |
|---|---|---|---|---|---|---|
| SSsP-1 | pAcrS1 | - | 10 | 529 | 3163 | DNA-binding protein |
| SSsP-1 | pAcrS2 | - | 9 | 5611 | 11,590 | DNA-binding protein |
| SSsP-1 | pAcrS3 | - | 2 | 14,071 | 14,808 | HNH endonuclease |
| SSsP-1 | pAcrS4 | - | 4 | 22,880 | 24,454 | HNH homing endonuclease |
| SSsP-1 | pAcrS5 | + | 2 | 25,842 | 26,812 | HNH homing endonuclease |
| SSsP-1 | pAcrS6 | - | 7 | 54,585 | 56,973 | None |
| GVEsP-1 | pAcrG1 | + | 2 | 4947 | 5568 | DNA-binding protein |
| GVEsP-1 | pAcrG2 | + | 11 | 22,501 | 26,731 | Phosphoesterase |
| GVEsP-1 | pAcrG3 | + | 6 | 30,688 | 32,712 | None |
| GVEsP-1 | pAcrG4 | + | 11 | 37,918 | 40,949 | None |
| GVEsP-1 | pAcrG5 | - | 3 | 52,269 | 53,288 | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tkachev, P.V.; Pchelin, I.M.; Azarov, D.V.; Gorshkov, A.N.; Shamova, O.V.; Dmitriev, A.V.; Goncharov, A.E. Two Novel Lytic Bacteriophages Infecting Enterococcus spp. Are Promising Candidates for Targeted Antibacterial Therapy. Viruses 2022, 14, 831. https://doi.org/10.3390/v14040831
Tkachev PV, Pchelin IM, Azarov DV, Gorshkov AN, Shamova OV, Dmitriev AV, Goncharov AE. Two Novel Lytic Bacteriophages Infecting Enterococcus spp. Are Promising Candidates for Targeted Antibacterial Therapy. Viruses. 2022; 14(4):831. https://doi.org/10.3390/v14040831
Chicago/Turabian StyleTkachev, Pavel V., Ivan M. Pchelin, Daniil V. Azarov, Andrey N. Gorshkov, Olga V. Shamova, Alexander V. Dmitriev, and Artemiy E. Goncharov. 2022. "Two Novel Lytic Bacteriophages Infecting Enterococcus spp. Are Promising Candidates for Targeted Antibacterial Therapy" Viruses 14, no. 4: 831. https://doi.org/10.3390/v14040831
APA StyleTkachev, P. V., Pchelin, I. M., Azarov, D. V., Gorshkov, A. N., Shamova, O. V., Dmitriev, A. V., & Goncharov, A. E. (2022). Two Novel Lytic Bacteriophages Infecting Enterococcus spp. Are Promising Candidates for Targeted Antibacterial Therapy. Viruses, 14(4), 831. https://doi.org/10.3390/v14040831