
A Single-Round Infection Fluorescent SARS-CoV-2 Neutralization Test for COVID-19 Serological Testing at a Biosafety Level-2 Laboratory


Abstract
:1. Introduction
2. Materials and Methods
2.1. Serum/Plasma Specimens
2.2. Cells
2.3. Preparation of SFV Stocks
2.4. SFV Neutralization Assay (SFVNT)
2.5. Plaque Reduction Neutralization Test (PRNT)
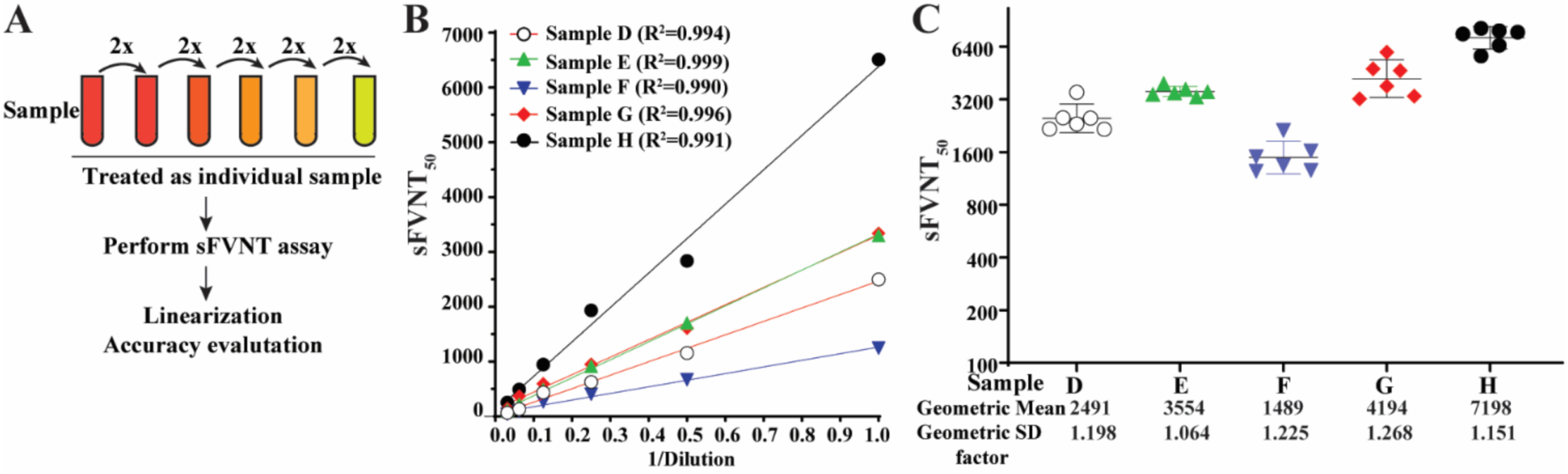
2.6. Linearity Analysis
2.7. Reproducibility
2.8. Ethics Statement
3. Results
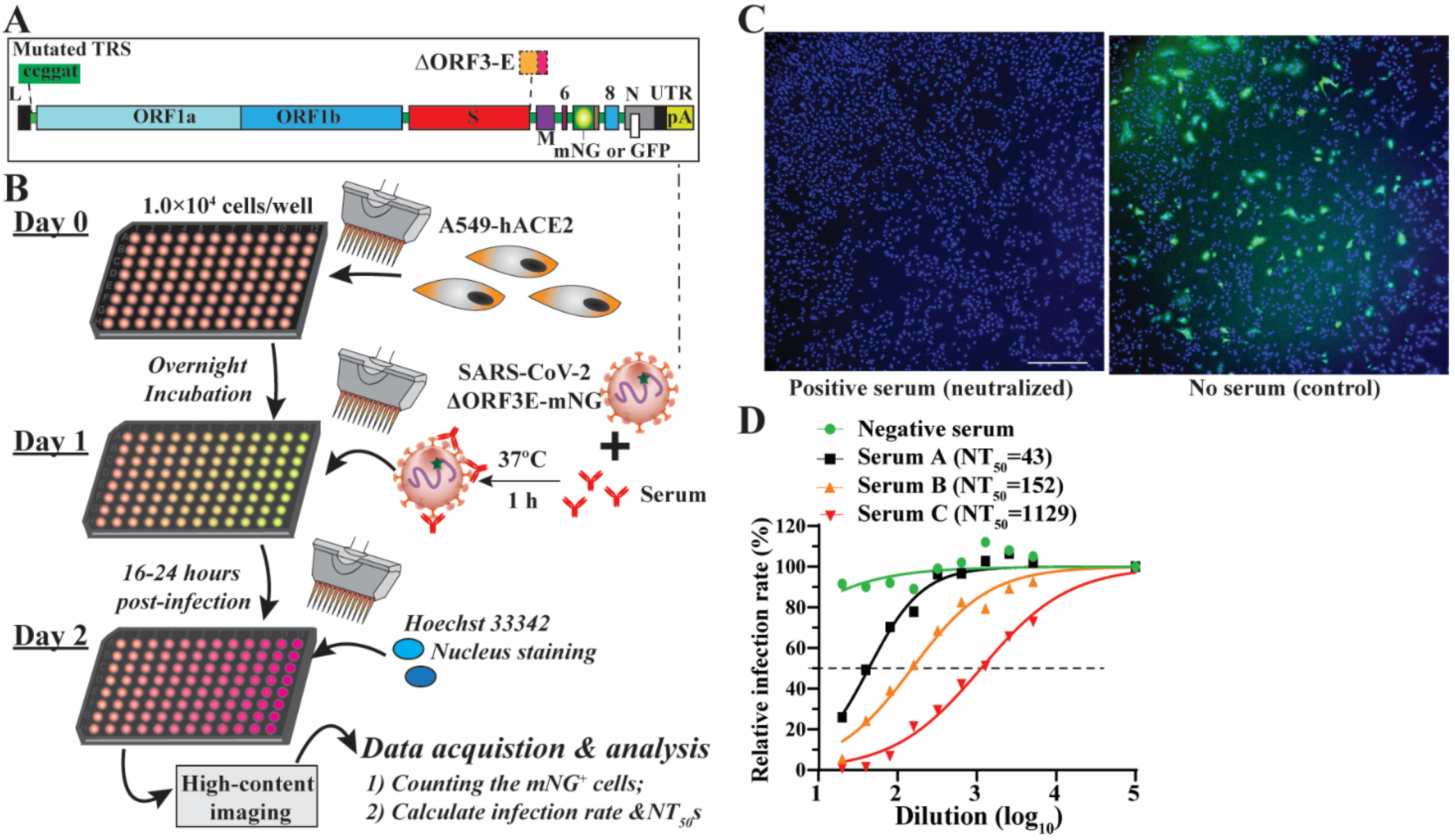
3.1. Rationale and Workflow
3.2. Optimization of the SFVNT
3.3. Accuracy of the SFVNT
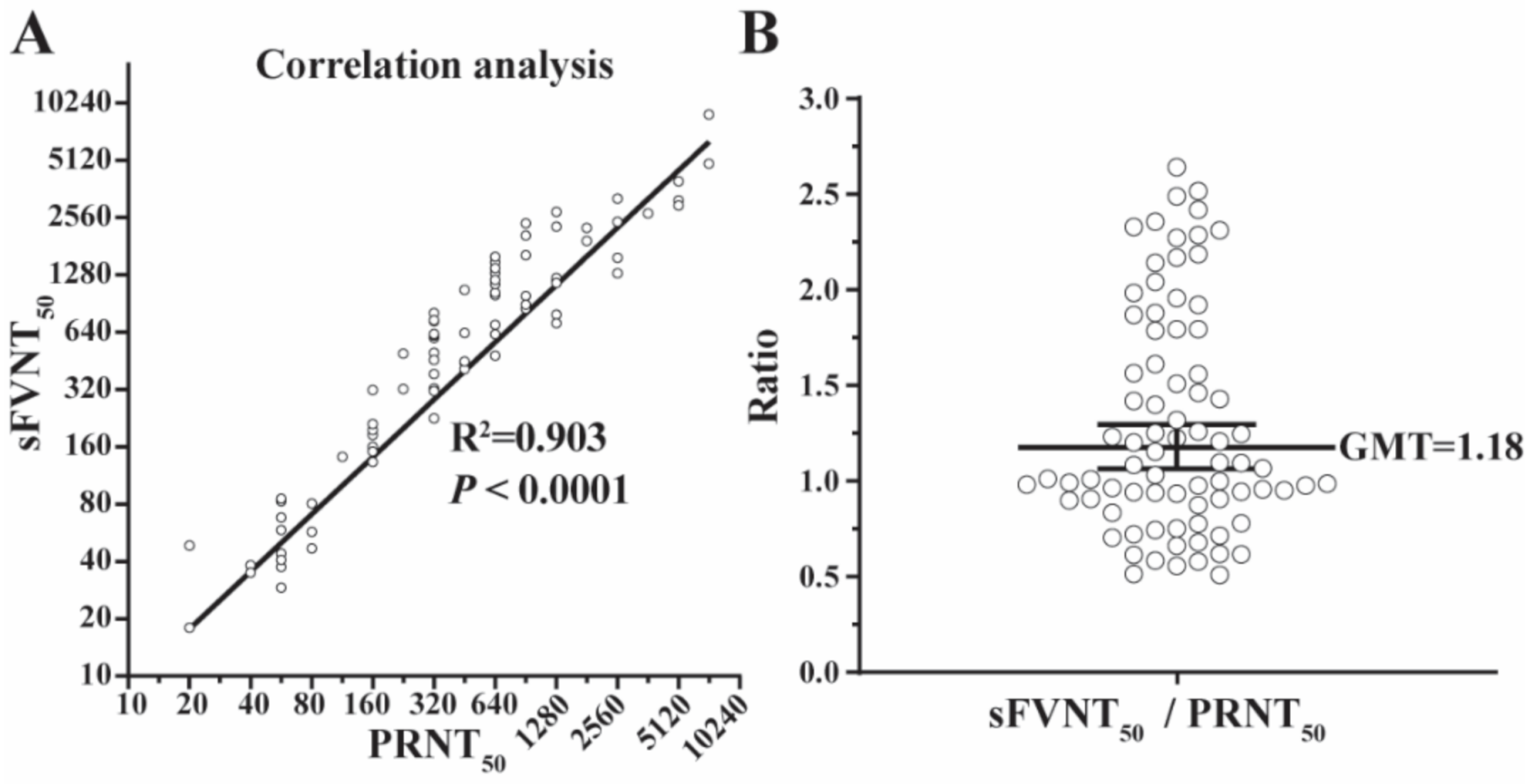
3.4. Correlation between the SFVNT and PRNT
3.5. Linearity
3.6. Analytical Specificity
3.7. Reproducibility
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, X.; Muruato, A.; Lokugamage, K.G.; Narayanan, K.; Zhang, X.; Zou, J.; Liu, J.; Schindewolf, C.; Bopp, N.E.; Aguilar, P.V.; et al. An Infectious cDNA Clone of SARS-CoV-2. Cell Host Microbe 2020, 27, 841–848.e3. [Google Scholar] [CrossRef]
- Sims, A.C.; Baric, R.S.; Yount, B.; Burkett, S.E.; Collins, P.L.; Pickles, R.J. Severe acute respiratory syndrome coronavirus infection of human ciliated airway epithelia: Role of ciliated cells in viral spread in the conducting airways of the lungs. J. Virol. 2005, 79, 15511–15524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muruato, A.E.; Fontes-Garfias, C.R.; Ren, P.; Garcia-Blanco, M.A.; Menachery, V.D.; Xie, X.; Shi, P.-Y. A high-throughput neutralizing antibody assay for COVID-19 diagnosis and vaccine evaluation. Nat. Commun. 2020, 11, 4059. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Y.; Liu, J.; Bailey, A.L.; Plante, K.S.; Plante, J.A.; Zou, J.; Xia, H.; Bopp, N.E.; Aguilar, P.V.; et al. A trans-complementation system for SARS-CoV-2 recapitulates authentic viral replication without virulence. Cell 2021, 184, 2229–2238.e13. [Google Scholar] [CrossRef] [PubMed]
- Almazán, F.; DeDiego, M.L.; Sola, I.; Zuñiga, S.; Nieto-Torres, J.L.; Marquez-Jurado, S.; Andrés, G.; Enjuanes, L. Engineering a replication-competent, propagation-defective Middle East respiratory syndrome coronavirus as a vaccine candidate. MBio 2013, 4, e00650-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, N.; Mossel, E.C.; Narayanan, K.; Popov, V.L.; Huang, C.; Inoue, T.; Peters, C.J.; Makino, S. Severe acute respiratory syndrome coronavirus 3a protein is a viral structural protein. J. Virol. 2005, 79, 3182–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Ku, Z.; Xie, X.; Davidson, E.; Ye, X.; Su, H.; Menachery, V.D.; Li, Y.; Yuan, Z.; Zhang, X.; Muruato, A.E.; et al. Molecular determinants and mechanism for antibody cocktail preventing SARS-CoV-2 escape. Nat. Commun. 2021, 12, 469. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Li, Q.; Wu, J.; Zhao, C.; Hao, H.; Liu, H.; Zhang, L.; Nie, L.; Qin, H.; Wang, M.; et al. Establishment and validation of a pseudovirus neutralization assay for SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 680–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Huang, B.; Ar, L.W.; Wang, W.; Deng, Y.; Tan, W. Development and effectiveness of pseudotyped SARS-CoV-2 system as determined by neutralizing efficiency and entry inhibition test in vitro. Biosaf. Health 2020, 2, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Case, J.B.; Rothlauf, P.W.; Chen, R.E.; Liu, Z.; Zhao, H.; Kim, A.S.; Bloyet, L.-M.; Zeng, Q.; Tahan, S.; Droit, L.; et al. Neutralizing Antibody and Soluble ACE2 Inhibition of a Replication-Competent VSV-SARS-CoV-2 and a Clinical Isolate of SARS-CoV-2. Cell Host Microbe 2020, 28, 475–485.e5. [Google Scholar] [CrossRef] [PubMed]
- Crawford, K.H.D.; Eguia, R.; Dingens, A.S.; Loes, A.N.; Malone, K.D.; Wolf, C.R.; Chu, H.Y.; Tortorici, M.A.; Veesler, D.; Murphy, M.; et al. Protocol and Reagents for Pseudotyping Lentiviral Particles with SARS-CoV-2 Spike Protein for Neutralization Assays. Viruses 2020, 12, 513. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Larragoite, E.T.; Williams, E.S.C.P.; Lama, J.; Cisneros, I.; Delgado, J.C.; Slev, P.; Rychert, J.; Innis, E.A.; Coiras, M.; et al. Neutralization assay with SARS-CoV-1 and SARS-CoV-2 spike pseudotyped murine leukemia virions. Virol. J. 2021, 18, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zettl, F.; Meister, T.L.; Vollmer, T.; Fischer, B.; Steinmann, J.; Krawczyk, A.; V’kovski, P.; Todt, D.; Steinmann, E.; Pfaender, S.; et al. Rapid Quantification of SARS-CoV-2-Neutralizing Antibodies Using Propagation-Defective Vesicular Stomatitis Virus Pseudotypes. Vaccines 2020, 8, 386. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.C.; Ren, P. SARS-CoV-2 serology testing: Progress and challenges. J. Immunol. Methods 2021, 494, 113060. [Google Scholar] [CrossRef] [PubMed]
- Kruglova, N.; Siniavin, A.; Gushchin, V.; Mazurov, D. Different Neutralization Sensitivity of SARS-CoV-2 Cell-to-Cell and Cell-Free Modes of Infection to Convalescent Sera. Viruses 2021, 13, 1133. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Evans, J.P.; King, T.; Zheng, Y.-M.; Oltz, E.M.; Whelan, S.P.J.; Saif, L.J.; Peeples, M.E.; Liu, S.-L. SARS-CoV-2 spreads through cell-to-cell transmission. Proc. Natl. Acad. Sci. USA 2022, 119, e2111400119. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.E.; Zhang, X.; Case, J.B.; Winkler, E.S.; Liu, Y.; VanBlargan, L.A.; Liu, J.; Errico, J.M.; Xie, X.; Suryadevara, N.; et al. Resistance of SARS-CoV-2 variants to neutralization by monoclonal and serum-derived polyclonal antibodies. Nat. Med. 2021, 27, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, J.; Xia, H.; Zhang, X.; Fontes-Garfias, C.R.; Swanson, K.A.; Cai, H.; Sarkar, R.; Chen, W.; Cutler, M.; et al. Neutralizing Activity of BNT162b2-Elicited Serum. N. Engl. J. Med. 2021, 384, 1466–1468. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, Y.; Xia, H.; Zou, J.; Weaver, S.C.; Swanson, K.A.; Cai, H.; Cutler, M.; Cooper, D.; Muik, A.; et al. BNT162b2-elicited neutralization of B.1.617 and other SARS-CoV-2 variants. Nature 2021, 596, 273–275. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serum ID | a PRNT50 | b SFVNT50 | Serum ID | a PRNT50 | b SFVNT50 |
|---|---|---|---|---|---|
| 1 | 20 | 18 | 41 | 453 | 1067 |
| 2 | 20 | 48 | 42 | 453 | 633 |
| 3 | 40 | 38 | 43 | 453 | 449 |
| 4 | 40 | 35 | 44 | 640 | 481 |
| 5 | 57 | 83 | 45 | 640 | 624 |
| 6 | 57 | 85 | 46 | 640 | 1000 |
| 7 | 57 | 37 | 47 | 640 | 700 |
| 8 | 57 | 58 | 48 | 640 | 1307 |
| 9 | 57 | 29 | 49 | 640 | 1491 |
| 10 | 57 | 44 | 50 | 640 | 1031 |
| 11 | 57 | 68 | 51 | 640 | 1143 |
| 12 | 57 | 41 | 52 | 640 | 1203 |
| 13 | 80 | 47 | 53 | 640 | 1389 |
| 14 | 80 | 57 | 54 | 640 | 693 |
| 15 | 80 | 81 | 55 | 640 | 1592 |
| 16 | 113 | 141 | 56 | 905 | 1623 |
| 17 | 160 | 185 | 57 | 905 | 993 |
| 18 | 160 | 318 | 58 | 905 | 850 |
| 19 | 160 | 196 | 59 | 905 | 2057 |
| 20 | 160 | 160 | 60 | 905 | 2391 |
| 21 | 160 | 133 | 61 | 905 | 892 |
| 22 | 160 | 151 | 62 | 1280 | 793 |
| 23 | 160 | 150 | 63 | 1280 | 1205 |
| 24 | 160 | 211 | 64 | 1280 | 1233 |
| 25 | 226 | 321 | 65 | 1280 | 1162 |
| 26 | 226 | 495 | 66 | 1280 | 713 |
| 27 | 320 | 499 | 67 | 1280 | 2296 |
| 28 | 320 | 731 | 68 | 1280 | 2741 |
| 29 | 320 | 457 | 69 | 1810 | 2255 |
| 30 | 320 | 323 | 70 | 1810 | 1929 |
| 31 | 320 | 598 | 71 | 2560 | 2434 |
| 32 | 320 | 225 | 72 | 2560 | 1303 |
| 33 | 320 | 314 | 73 | 2560 | 1573 |
| 34 | 320 | 615 | 74 | 2560 | 3222 |
| 35 | 320 | 806 | 75 | 3620 | 2695 |
| 36 | 320 | 386 | 76 | 5120 | 3975 |
| 37 | 320 | 740 | 77 | 5120 | 3151 |
| 38 | 320 | 626 | 78 | 5120 | 2962 |
| 39 | 453 | 441 | 79 | 7241 | 4911 |
| 40 | 453 | 410 | 80 | 7241 | 8913 |
| a Immune Sera and b Interfering Substances | Sample Number | Number of SFVNT-Positive |
|---|---|---|
| Adenovirus | 1 | 0 |
| Cryptococcus neoformans antigen | 1 | 0 |
| Anti-cytomegalovirus | 10 | 0 |
| Anti-Epstein–Barr virus capsid or nuclear antigen | 15 | 0 |
| Anti-hepatitis A virus | 9 | 0 |
| Anti-hepatitis B virus surface or core antigen | 21 | 0 |
| Anti-hepatitis C virus | 7 | 0 |
| Anti-herpes simplex virus 1 | 8 | 0 |
| Anti-herpes simplex virus 2 | 3 | 0 |
| Human coronavirus 229E | 1 | 0 |
| Human coronavirus HKU1 | 2 | 0 |
| Human coronavirus NL63 | 1 | 0 |
| Human coronavirus OC43 | 3 | 0 |
| Anti-human immunodeficiency virus 1 | 4 | 0 |
| Human rhinovirus | 2 | 0 |
| Influenza B virus | 1 | 0 |
| Anti-measles virus | 2 | 0 |
| Anti-mumps virus | 1 | 0 |
| Respiratory syncytial virus | 1 | 0 |
| Anti-rubella virus | 16 | 0 |
| Anti-varicella zoster virus | 20 | 0 |
| Anti-West Nile virus | 5 | 0 |
| Anti-yellow fever virus (vaccination) | 1 | 0 |
| b Albumin (4.5–4.9 g/dL) | 7 | 0 |
| b Elevated total bilirubin (1.1 mg/dL) | 5 | 0 |
| b Elevated cholesterol (>200 mg/dL) | 3 | 0 |
| b Elevated rheumatoid factor (>100 IU/mL) | 4 | 0 |
| b Anti-cardiolipin | 2 | 0 |
| b Anti-mitochondrial M2 | 1 | 0 |
| b Anti-nuclear antibodies | 10 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, J.; Xia, H.; Shi, P.-Y.; Xie, X.; Ren, P. A Single-Round Infection Fluorescent SARS-CoV-2 Neutralization Test for COVID-19 Serological Testing at a Biosafety Level-2 Laboratory. Viruses 2022, 14, 1211. https://doi.org/10.3390/v14061211
Zou J, Xia H, Shi P-Y, Xie X, Ren P. A Single-Round Infection Fluorescent SARS-CoV-2 Neutralization Test for COVID-19 Serological Testing at a Biosafety Level-2 Laboratory. Viruses. 2022; 14(6):1211. https://doi.org/10.3390/v14061211
Chicago/Turabian StyleZou, Jing, Hongjie Xia, Pei-Yong Shi, Xuping Xie, and Ping Ren. 2022. "A Single-Round Infection Fluorescent SARS-CoV-2 Neutralization Test for COVID-19 Serological Testing at a Biosafety Level-2 Laboratory" Viruses 14, no. 6: 1211. https://doi.org/10.3390/v14061211
APA StyleZou, J., Xia, H., Shi, P. -Y., Xie, X., & Ren, P. (2022). A Single-Round Infection Fluorescent SARS-CoV-2 Neutralization Test for COVID-19 Serological Testing at a Biosafety Level-2 Laboratory. Viruses, 14(6), 1211. https://doi.org/10.3390/v14061211