
High Incidence of SARS-CoV-2 Variant of Concern Breakthrough Infections Despite Residual Humoral and Cellular Immunity Induced by BNT162b2 Vaccination in Healthcare Workers: A Long-Term Follow-Up Study in Belgium

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
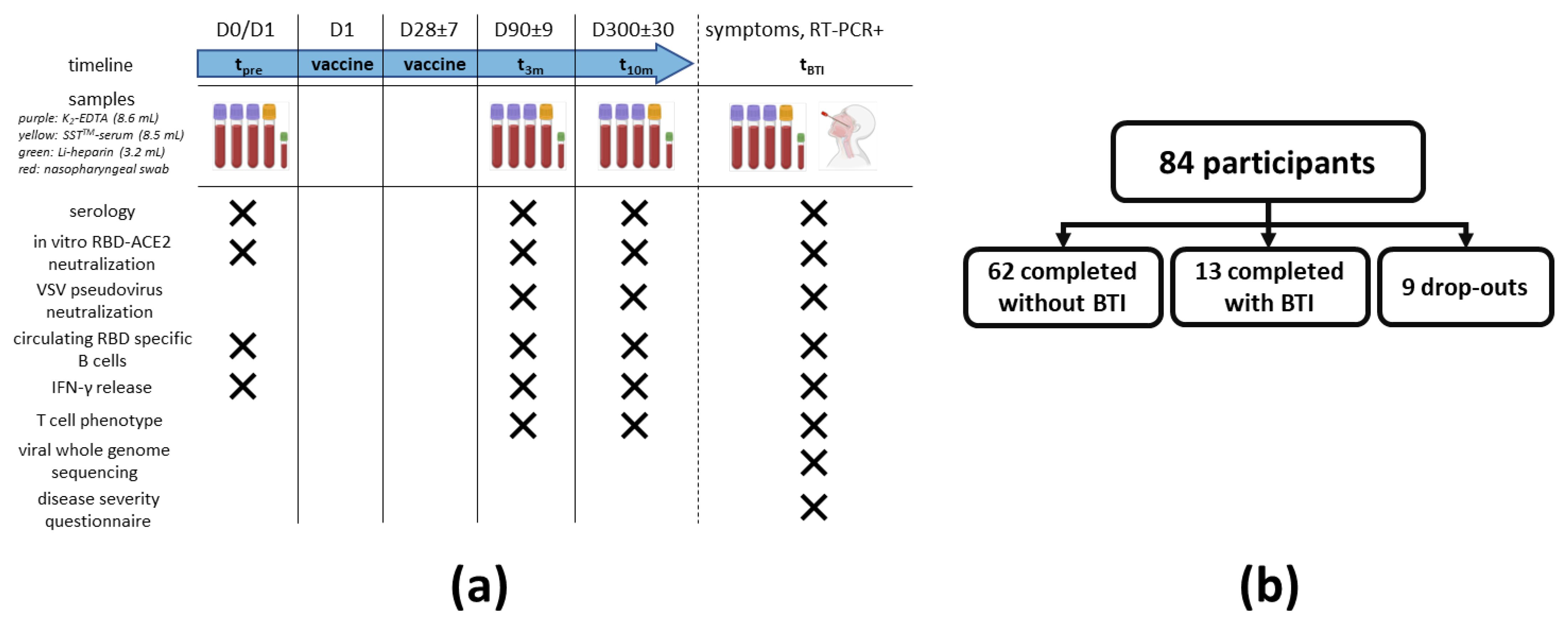
2.1. Study Design
2.1.1. Recruitment
2.1.2. Sampling
2.1.3. Peripheral Blood Mononuclear Cells (PBMC) Isolation and Serum Collection
2.2. Serological Parameters
2.2.1. Anti-S IgA and IgG Assay
2.2.2. Anti-RBD IgG Assay
2.2.3. Anti-N IgG Assay
2.3. Functional Assessment of Vaccine-Induced Antibodies
2.3.1. In Vitro RBD-ACE2 Binding Inhibition Assay
2.3.2. Vesicular Stomatitis Virus (VSV) Pseudovirus Neutralization Assay
2.4. Analysis of SARS-CoV-2-Specific T Cells
2.4.1. Interferon γ (IFN-γ) ELISA
2.4.2. T Cell Phenotyping
2.5. Quantification of Circulating RBD-Specific B Cells
2.6. Viral Whole Genome Sequencing
2.7. Statistical Analyses
3. Results
3.1. Trial Characteristics and Exclusions
3.2. Preservation of Functional In Vitro Neutralization over Time despite Waning Antibody Levels
3.3. A Minority of Unaffected Individuals Display Circulating RBD-Specific B Cells with Different Kinetics and Correlation to Serology over Time
3.4. Residual SARS-CoV-2-Specific T Cell Activity Is More Retained in CD8+ Than CD4+ T Cells
3.5. Humoral and Cellular Responses upon BNT162b2 Vaccination Are Not Correlated
3.6. Real-World Incidence of Symptomatic BNT162b2 Breakthrough Infections
3.7. Participants with BTI Show Unaltered Cellular Responses but Compromised Humoral Immunity to Vaccination at Mid-Term Follow-Up
3.8. Vaccine-Induced SARS-CoV-2 Neutralizing Antibodies Are Not Sufficient to Prevent BTI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kreye, J.; Reincke, S.M.; Kornau, H.C.; Sánchez-Sendin, E.; Corman, V.M.; Liu, H.; Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.C.D.; et al. A Therapeutic Non-self-reactive SARS-CoV-2 Antibody Protects from Lung Pathology in a COVID-19 Hamster Model. Cell 2020, 183, 1058–1069.e19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Z. Novel 2019 Coronavirus Genome—SARS-CoV-2 Coronavirus—Virological. 2020, pp. 1–7. Available online: Virological.org (accessed on 28 February 2022).
- Zhong, N.S.; Zheng, B.J.; Li, Y.M.; Poon, L.L.M.; Xie, Z.H.; Chan, K.H.; Li, P.H.; Tan, S.Y.; Chang, Q.; Xie, J.P.; et al. Epidemiology and cause of severe acute respiratory syndrome (SARS) in Guangdong, People’s Republic of China, in February, 2003. Lancet 2003, 362, 1353–1358. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, P.; Wang, J.; Feng, J.; Zhou, H.; Li, X.; Zhong, W.; Hao, P. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Sci. China Life Sci. 2020, 63, 457–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Yang, X.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. World Health Organization: Coronavirus; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- OMS. IHR Emergency Committee on Novel Coronavirus (2019-nCoV); OMS: Geneva, Switzerland, 2020; pp. 1–4. [Google Scholar]
- Romani, G.; Dal Mas, F.; Massaro, M.; Cobianchi, L.; Modenese, M.; Barcellini, A.; Ricciardi, W.; Barach, P.; Lucà, R.; Ferrara, M. Population Health Strategies to Support Hospital and Intensive Care Unit Resiliency during the COVID-19 Pandemic: The Italian Experience. Popul. Health Manag. 2021, 24, 174–181. [Google Scholar] [CrossRef]
- FDA. Pfizer-BioNTech COVID-19 Vaccine; FDA: Silver Spring, MD, USA, 2021. [Google Scholar]
- EMA—Committee for Medicinal Products for Human Use. Pfizer-BioNTech COVID-19 Vaccine EMA Public Assesment Report; EMA: Amsterdam, The Netherlands, 2021. [Google Scholar]
- EMA—Committee for Medicinal Products for Human Use. Moderna COVID-19 Vaccine EMA Public Assessment Report; EMA: Amsterdam, The Netherlands, 2021. [Google Scholar]
- FDA. Moderna COVID-19 Vaccine; FDA: Silver Spring, MD, USA, 2021. [Google Scholar]
- EMA—Committee for Medicinal Products for Human Use. AstraZeneca EMA Public Assessment Report; EMA: Amsterdam, The Netherlands, 2021; Volume 31. [Google Scholar]
- EMA—Committee for Medicinal Products for Human. COVID-19 Vaccine Janssen EMA Public Assessment Report; EMA: Amsterdam, The Netherlands, 2021; Volume 31. [Google Scholar]
- FDA. Janssen COVID-19 Vaccine; FDA: Silver Spring, MD, USA, 2021. [Google Scholar]
- Bettini, E.; Locci, M. SARS-CoV-2 mRNA Vaccines: Immunological mechanism and beyond. Vaccines 2021, 9, 147. [Google Scholar] [CrossRef]
- Ciabattini, A.; Pastore, G.; Fiorino, F.; Polvere, J.; Lucchesi, S.; Pettini, E.; Auddino, S.; Rancan, I.; Durante, M.; Miscia, M.; et al. Evidence of SARS-CoV-2-Specific Memory B Cells Six Months After Vaccination with the BNT162b2 mRNA Vaccine. Front. Immunol. 2021, 12, 3751. [Google Scholar] [CrossRef]
- Gil-Manso, S.; Carbonell, D.; López-Fernández, L.; Miguens, I.; Alonso, R.; Buño, I.; Muñoz, P.; Ochando, J.; Pion, M.; Correa-Rocha, R. Induction of High Levels of Specific Humoral and Cellular Responses to SARS-CoV-2 After the Administration of COVID-19 mRNA Vaccines Requires Several Days. Front. Immunol. 2021, 12, 3970. [Google Scholar] [CrossRef]
- Lombardi, A.; Bozzi, G.; Ungaro, R.; Villa, S.; Castelli, V.; Mangioni, D.; Muscatello, A.; Gori, A.; Bandera, A. Mini Review Immunological Consequences of Immunization With COVID-19 mRNA Vaccines: Preliminary Results. Front. Immunol. 2021, 12, 677. [Google Scholar] [CrossRef]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Zavaglio, F.; Frangipane, V.; Morosini, M.; Gabanti, E.; Zelini, P.; Sammartino, J.C.; Ferrari, A.; Gregorini, M.; Rampino, T.; Asti, A.; et al. Robust and persistent b-and t-cell responses after covid-19 in immunocompetent and solid organ transplant recipient patients. Viruses 2021, 13, 2261. [Google Scholar] [CrossRef] [PubMed]
- Calcoen, B.; Callebaut, K.; Vandenbulcke, A.; Callewaert, N.; Bossuyt, X.; Van Weyenbergh, J.; Maes, P.; Imbrechts, M.; Vercruysse, T.; Thibaut, H.J.; et al. Real-world monitoring of BNT162b2 vaccine-induced SARS-CoV-2 B and T cell immunity in naive healthcare workers: A prospective single center study. medRxiv 2022. [Google Scholar] [CrossRef]
- WHO. Tracking SARS-CoV-2 Variants. WHO. 2021. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 1 April 2022).
- WHO. Tracking SARS-CoV-2 Variants; WHO: Geneva, Switzerland, 2021; pp. 1–13. Available online: https://www.who.int/docs/default-source/coronaviruse/2022-01-07-global-technical-brief-and-priority-action-on-omicron---corr2.pdf?sfvrsn=918b09d_20 (accessed on 1 April 2022).
- Otto, S.P.; Day, T.; Arino, J.; Colijn, C.; Dushoff, J.; Li, M.; Mechai, S.; Van Domselaar, G.; Wu, J.; Earn, D.J.D.; et al. The origins and potential future of SARS-CoV-2 variants of concern in the evolving COVID-19 pandemic. Curr. Biol. 2021, 31, R918–R929. [Google Scholar] [CrossRef] [PubMed]
- Hacisuleyman, E.; Hale, C.; Saito, Y.; Blachere, N.E.; Bergh, M.; Conlon, E.G.; Schaefer-Babajew, D.J.; DaSilva, J.; Muecksch, F.; Gaebler, C.; et al. Vaccine Breakthrough Infections with SARS-CoV-2 Variants. N. Engl. J. Med. 2021, 384, 2212–2218. [Google Scholar] [CrossRef] [PubMed]
- Glatman-Freedman, A.; Bromberg, M.; Dichtiar, R.; Hershkovitz, Y.; Keinan-Boker, L. The BNT162b2 vaccine effectiveness against new COVID-19 cases and complications of breakthrough cases: A nation-wide retrospective longitudinal multiple cohort analysis using individualised data. EBioMedicine 2021, 72, 103574. [Google Scholar] [CrossRef] [PubMed]
- Blanquart, F.; Abad, C.; Ambroise, J.; Bernard, M.; Cosentino, G.; Giannoli, J.M.; Debarre, F. Characterisation of vaccine breakthrough infections of sars-cov-2 delta and alpha variants and within-host viral load dynamics in the community, France, June to July 2021. Eurosurveillance 2021, 26, 2100824. [Google Scholar] [CrossRef]
- Tene, Y.; Levytskyi, K.; Adler, A.; Halutz, O.; Paran, Y.; Goldshmidt, H.; Itzhaki, A.; Halperin, T.; Stefansky, S.; Ben-Ami, R.; et al. An outbreak of SARS-CoV-2 infections among hospital personnel with high mRNA vaccine uptake. In Infection Control Hospital Epidemiology; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Bahl, A.; Johnson, S.; Maine, G.; Garcia, M.H.; Nimmagadda, S.; Qu, L.; Chen, N.-W. Vaccination reduces need for emergency care in breakthrough COVID-19 infections: A multicenter cohort study. Lancet Reg. Health Am. 2021, 4, 100065. [Google Scholar] [CrossRef]
- BioRender. Available online: www.biorender.com (accessed on 1 January 2022).
- EMA—Committee for Medicinal Products for Human Use. Guideline on Bioanalytical Method Validation; EMEA/CHMP/EWP/192217/2009 Rev 1 Corr 2**; EMA: Amsterdam, The Netherlands, 2012; pp. 1–23. [Google Scholar]
- NIBSC NI for BS and Controls. WHO International Standard First WHO International Standard for Anti-SARS-CoV-2 Immunoglobulin (Human) NIBSC Code: 20/136 Instructions for Use (Version 2.0, Dated 17/12/2020); NIBSC: Ridge, UK, 2020. [Google Scholar]
- Sanchez-Felipe, L.; Vercruysse, T.; Sharma, S.; Ma, J.; Lemmens, V.; Van Looveren, D.; Arkalagud Javarappa, M.P.; Boudewijns, R.; Malengier-Devlies, B.; Liesenborghs, L.; et al. A single-dose live-attenuated YF17D-vectored SARS-CoV-2 vaccine candidate. Nature 2021, 590, 320–325. [Google Scholar] [CrossRef]
- Whitt, M.A. Generation of VSV pseudotypes using recombinant ΔG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. J. Virol. Methods 2010, 169, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Imbrechts, M.; Maes, W.; Ampofo, L.; Van Den Berghe, N.; Calcoen, B.; Van Looveren, D.; Noppen, S.; Hollevoet, K.; Vercruysse, T.; Zhang, X.; et al. Potent neutralizing anti-SARS-CoV-2 human antibodies cure infection with SARS-CoV-2 variants in hamster model. bioRxiv 2021. [Google Scholar] [CrossRef]
- Van Cleemput, J.; Van Snippenberg, W.; Lambrechts, L.; Dendooven, A.; D’Onofrio, V.; Couck, L.; Trypsteen, W.; Vanrusselt, J.; Theuns, S.; Vereecke, N.; et al. Organ-specific genome diversity of replication-competent SARS-CoV-2. Nat. Commun. 2021, 12, 6612. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2010; ISBN 9780131008465. [Google Scholar]
- Diaz, J.; Appiah, J.; Askie, L.; Baller, A.; Banerjee, A.; Barkley, S.; Bertagnolio, S.; Hemmingsen, B.; Bonet, M.; Cunningham, J. COVID-19: Clinical Management Living Guidance; World Health Organization: Geneva, Switzerland, 2021; p. 81. [Google Scholar]
- COVID-19 Wekelijks Epidemiologisch Bulletin. Available online: https://covid-19.sciensano.be/sites/default/files/Covid19/Meest%20recente%20update.pdf (accessed on 1 April 2022).
- Despres, H.W.; Mills, M.G.; Shirley, D.J.; Schmidt, M.M.; Huang, M.-L.; Jerome, K.R.; Greninger, A.L.; Bruce, E.A. Quantitative measurement of infectious virus in SARS-CoV-2 Alpha, Delta and Epsilon variants reveals higher infectivity (viral titer:RNA ratio) in clinical samples containing the Delta and Epsilon variants. medRxiv Prepr. Serv. Health Sci. 2021. [Google Scholar] [CrossRef]
- Baele, G.; Cuypers, L.; Maes, P.; Dellicour, S.; Keyaerts, E.; Wollants, E.; Ranst, M. Van, André, E. Genomic Surveillance of SARS-CoV-2 in Belgium; UZ Leuven: Leuven, Belgium, 2021; Volume 2021. [Google Scholar]
- Sciensano. COVID-19 Surveillance Frequently Asked Questions; Sciensano: Brussels, Belgium, 2021. [Google Scholar]
- Amodio, D.; Ruggiero, A.; Sgrulletti, M.; Pighi, C.; Cotugno, N.; Medri, C.; Morrocchi, E.; Colagrossi, L.; Russo, C.; Zaffina, S.; et al. Humoral and Cellular Response Following Vaccination with the BNT162b2 mRNA COVID-19 Vaccine in Patients Affected by Primary Immunodeficiencies. Front. Immunol. 2021, 12, 3947. [Google Scholar] [CrossRef] [PubMed]
- Braun-Moscovici, Y.; Kaplan, M.; Braun, M.; Markovits, D.; Giryes, S.; Toledano, K.; Tavor, Y.; Dolnikov, K.; Balbir-Gurman, A. Disease activity and humoral response in patients with inflammatory rheumatic diseases after two doses of the Pfizer mRNA vaccine against SARS-CoV-2. Ann. Rheum. Dis. 2021, 80, 1317–1321. [Google Scholar] [CrossRef]
- Achiron, A.; Mandel, M.; Dreyer-Alster, S.; Harari, G.; Magalashvili, D.; Sonis, P.; Dolev, M.; Menascu, S.; Flechter, S.; Falb, R.; et al. Humoral immune response to COVID-19 mRNA vaccine in patients with multiple sclerosis treated with high-efficacy disease-modifying therapies. Ther. Adv. Neurol. Disord. 2021, 14, 17562864211012835. [Google Scholar] [CrossRef]
- Grupper, A.; Sharon, N.; Finn, T.; Cohen, R.; Israel, M.; Agbaria, A.; Rechavi, Y.; Schwartz, I.F.; Schwartz, D.; Lellouch, Y.; et al. Humoral response to the pfizer bnt162b2 vaccine in patients undergoing maintenance hemodialysis. Clin. J. Am. Soc. Nephrol. 2021, 16, 1037–1042. [Google Scholar] [CrossRef]
- Zitt, E.; Davidovic, T.; Schimpf, J.; Abbassi-Nik, A.; Mutschlechner, B.; Ulmer, H.; Benda, M.A.; Sprenger-Mähr, H.; Winder, T.; Lhotta, K. The Safety and Immunogenicity of the mRNA-BNT162b2 SARS-CoV-2 Vaccine in Hemodialysis Patients. Front. Immunol. 2021, 12, 2390. [Google Scholar] [CrossRef]
- Lustig, Y.; Sapir, E.; Regev-Yochay, G.; Cohen, C.; Fluss, R.; Olmer, L.; Indenbaum, V.; Mandelboim, M.; Doolman, R.; Amit, S.; et al. BNT162b2 COVID-19 vaccine and correlates of humoral immune responses and dynamics: A prospective, single-centre, longitudinal cohort study in health-care workers. Lancet Respir. Med. 2021, 9, 999–1009. [Google Scholar] [CrossRef]
- Segundo, D.S.; Comins-Boo, A.; Irure-Ventura, J.; Renuncio-García, M.; Roa-Bautista, A.; González-Lípez, E.; Merino-Fernández, D.; Lamadrid-Perojo, P.; Alonso-Peña, M.; Ocejo-Vinyals, J.; et al. Immune assessment of BNT162b2 m-RNA-spike based vaccine response in adults. Biomedicines 2021, 9, 868. [Google Scholar] [CrossRef]
- Wang, Z.; Schmidt, F.; Weisblum, Y.; Muecksch, F.; Barnes, C.O.; Finkin, S.; Schaefer-Babajew, D.; Cipolla, M.; Gaebler, C.; Lieberman, J.A.; et al. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. Nature 2021, 592, 616–622. [Google Scholar] [CrossRef]
- Müller-Hilke, B.; Mai, F.; Müller, M.; Volzke, J.; Reisinger, E.C. Higher SARS-CoV-2 Spike Binding Antibody Levels and Neutralization Capacity 6 Months after Heterologous Vaccination with AZD1222 and BNT162b2. Vaccines 2022, 10, 322. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Espinosa, D.; Montagud-Marrahi, E.; Cacho, J.; Arana, C.; Taurizano, N.; Hermida, E.; Del Risco-Zevallos, J.; Casals, J.; Rosario, A.; Cuadrado-Payán, E.; et al. Incidence of severe breakthrough SARS-CoV-2 infections in vaccinated kidney transplant and haemodialysis patients. J. Nephrol. 2022, 35, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Boedecker-Lips, S.C.; Lautem, A.; Runkel, S.; Klimpke, P.; Kraus, D.; Keil, P.; Holtz, S.; Tomalla, V.; Marczynski, P.; Boedecker, C.B.; et al. Six-Month Follow-Up after Vaccination with BNT162b2: SARS-CoV-2 Antigen-Specific Cellular and Humoral Immune Responses in Hemodialysis Patients and Kidney Transplant Recipients. Pathogens 2022, 11, 67. [Google Scholar] [CrossRef] [PubMed]
- Casado, J.L.; Haemmerle, J.; Vizcarra, P.; Rodriguez-Dominguez, M.; Velasco, T.; Velasco, H.; Velasco, H.; Centenera, E.; Romero-Hernandez, B.; Fernandez-Escribano, M.; et al. T-cell response after first dose of BNT162b2 SARS-CoV-2 vaccine among healthcare workers with previous infection or cross-reactive immunity. Clin. Transl. Immunol. 2021, 10, e1341. [Google Scholar] [CrossRef]
- Hirotsu, Y.; Amemiya, K.; Sugiura, H.; Shinohara, M.; Takatori, M.; Mochizuki, H.; Omata, M. Robust Antibody Responses to the BNT162b2 mRNA Vaccine Occur Within a Week After the First Dose in Previously Infected Individuals and After the Second Dose in Uninfected Individuals. Front. Immunol. 2021, 12, 3457. [Google Scholar] [CrossRef]
- Chivu-Economescu, M.; Bleotu, C.; Grancea, C.; Chiriac, D.; Botezatu, A.; Iancu, I.V.; Pitica, I.; Necula, L.G.; Neagu, A.; Matei, L.; et al. Kinetics and persistence of cellular and humoral immune responses to SARS-CoV-2 vaccine in healthcare workers with or without prior COVID-19. J. Cell Mol. Med. 2022, 26, 1293–1305. [Google Scholar] [CrossRef]
- Spitzer, A.; Angel, Y.; Marudi, O.; Zeltser, D.; Saiag, E.; Goldshmidt, H.; Goldiner, I.; Stark, M.; Halutz, O.; Gamzu, R.; et al. Association of a Third Dose of BNT162b2 Vaccine with Incidence of SARS-CoV-2 Infection among Health Care Workers in Israel. JAMA J. Am. Med. Assoc. 2022, 327, 341–349. [Google Scholar] [CrossRef]
- Wisnewski, A.V.; Luna, J.C.; Redlich, C.A. Human IgG and IgA responses to COVID-19 mRNA vaccines. PLoS ONE 2021, 16, e0249499. [Google Scholar] [CrossRef]
- Pratesi, F.; Caruso, T.; Testa, D.; Tarpanelli, T.; Gentili, A.; Gioè, D.; Migliorini, P. Bnt162b2 mrna sars-cov-2 vaccine elicits high avidity and neutralizing antibodies in healthcare workers. Vaccines 2021, 9, 672. [Google Scholar] [CrossRef]
- Salvagno, G.L.; Henry, B.M.; Di Piazza, G.; Pighi, L.; De Nitto, S.; Bragantini, D.; Gianfilippi, G.; Lippi, G. Anti-spike s1 iga, anti-spike trimeric igg, and anti-spike rbd igg response after bnt162b2 covid-19 mrna vaccination in healthcare workers. J. Med. Biochem. 2021, 40, 327–334. [Google Scholar] [CrossRef]
- Naaber, P.; Tserel, L.; Kangro, K.; Sepp, E.; Jürjenson, V.; Adamson, A.; Haljasmägi, L.; Rumm, A.P.; Maruste, R.; Kärner, J.; et al. Dynamics of antibody response to BNT162b2 vaccine after six months: A longitudinal prospective study. Lancet Reg. Health Eur. 2021, 10, 100208. [Google Scholar] [CrossRef] [PubMed]
- Hatzakis, A.; Karabinis, A.; Roussos, S.; Pantazis, N.; Degiannis, D.; Chaidaroglou, A.; Petsios, K.; Pavlopoulou, I.; Tsiodras, S.; Paraskevis, D.; et al. Modelling SARS-CoV-2 Binding Antibody Waning 8 Months after BNT162b2 Vaccination. Vaccines 2022, 10, 285. [Google Scholar] [CrossRef] [PubMed]
- Xiaojie, S.; Yu, L.; Lei, Y.; Guang, Y.; Min, Q. Neutralizing antibodies targeting SARS-CoV-2 spike protein. Stem Cell Res. 2021, 50, 102125. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, P.J.M.; Caniels, T.G.; van der Straten, K.; Snitselaar, J.L.; Aldon, Y.; Bangaru, S.; Torres, J.L.; Okba, N.M.A.; Claireaux, M.; Kerster, G.; et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science 2020, 369, 643–650. [Google Scholar] [CrossRef]
- Byazrova, M.; Yusubalieva, G.; Spiridonova, A.; Efimov, G.; Mazurov, D.; Baranov, K.; Baklaushev, V.; Filatov, A. Pattern of circulating SARS-CoV-2-specific antibody-secreting and memory B-cell generation in patients with acute COVID-19. Clin. Transl. Immunol. 2021, 10, e1245. [Google Scholar] [CrossRef]
- Turner, J.S.; O’Halloran, J.A.; Kalaidina, E.; Kim, W.; Schmitz, A.J.; Zhou, J.Q.; Lei, T.; Thapa, M.; Chen, R.E.; Case, J.B.; et al. SARS-CoV-2 mRNA vaccines induce persistent human germinal centre responses. Nature 2021, 596, 109–113. [Google Scholar] [CrossRef]
- Terreri, S.; Piano Mortari, E.; Vinci, M.R.; Russo, C.; Alteri, C.; Albano, C.; Colavita, F.; Gramigna, G.; Agrati, C.; Linardos, G.; et al. Persistent B cell memory after SARS-CoV-2 vaccination is functional during breakthrough infections. Cell Host Microbe. 2022, 30, 400–408.e4. [Google Scholar] [CrossRef]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Moyon, Q.; Sterlin, D.; Miyara, M.; Anna, F.; Mathian, A.; Lhote, R.; Ghillani-Dalbin, P.; Breillat, P.; Mudumba, S.; de Alba, S.; et al. BNT162b2 vaccine-induced humoral and cellular responses against SARS-CoV-2 variants in systemic lupus erythematosus. Ann. Rheum. Dis. 2021, 81, 575–583. [Google Scholar] [CrossRef]
- Lasagna, A.; Agustoni, F.; Percivalle, E.; Borgetto, S.; Paulet, A.; Comolli, G.; Sarasini, A.; Bergami, F.; Sammartino, J.C.; Ferrari, A.; et al. A snapshot of the immunogenicity, efficacy and safety of a full course of BNT162b2 anti-SARS-CoV-2 vaccine in cancer patients treated with PD-1/PD-L1 inhibitors: A longitudinal cohort study. ESMO Open 2021, 6, 100272. [Google Scholar] [CrossRef]
- Van Praet, J.; Reynders, M.; De Bacquer, D.; Viaene, L.; Schoutteten, M.; Caluwé, R.; Doubel, P.; Heylen, L.; De Bel, A.; Steensels, D.; et al. Predictors and Dynamics of the Humoral and Cellular Immune Response to SARS-CoV-2 mRNA Vaccines in Hemodialysis Patients: A Multicenter Observational Study. J. Am. Soc. Nephrol. 2021, 32, 3208–3220. [Google Scholar] [CrossRef] [PubMed]
- Painter, M.M.; Mathew, D.; Goel, R.R.; Apostolidis, S.A.; Pattekar, A.; Kuthuru, O.; Baxter, A.E.; Herati, R.S.; Oldridge, D.A.; Gouma, S.; et al. Rapid induction of antigen-specific CD4+ T cells is associated with coordinated humoral and cellular immunity to SARS-CoV-2 mRNA vaccination. Immunity 2021, 54, 2133–2142.e3. [Google Scholar] [CrossRef] [PubMed]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Stouten, V.; Hubin, P.; Haarhuis, F.; van Loenhout, J.; Billuart, M.; Brondeel, R.; Braeye, T.; Van Oyen, H.; Wyndham-Thomas, C.; Catteau, L. Incidence and Risk Factors of COVID-19 Vaccine Breakthrough Infections: A Prospective Cohort Study in Belgium. Viruses 2022, 14, 802. [Google Scholar] [CrossRef]
- Heftdal, L.D.; Schultz, M.; Lange, T.; Knudsen, A.D.; Fogh, K.; Hasselbalch, R.B.; Linander, C.B.; Kallemose, T.; Bundgaard, H.; Grønbæk, K.; et al. Incidence of positive SARS-CoV-2 PCR after COVID-19 vaccination with up to eight months of follow-up: Real life data from the Capital Region of Denmark. Clin. Infect. Dis. 2022, ciac012. [Google Scholar] [CrossRef]
- Ponticelli, D.; Madotto, F.; Conti, S.; Antonazzo, I.C.; Vitale, A.; Della Ragione, G.; Romano, M.L.; Borrelli, M.; Schiavone, B.; Polosa, R.; et al. Response to BNT162b2 mRNA COVID-19 vaccine among healthcare workers in Italy: A 3-month follow-up. Intern. Emerg. Med. 2021, 17, 481–486. [Google Scholar] [CrossRef]
- Porru, S.; Spiteri, G.; Monaco, M.G.L.; Valotti, A.; Carta, A.; Lotti, V.; Diani, E.; Lippi, G.; Gibellini, D.; Verlato, G. Post-Vaccination SARS-CoV-2 Infections among Health Workers at the University Hospital of Verona, Italy: A Retrospective Cohort Survey. Vaccines 2022, 10, 272. [Google Scholar] [CrossRef]
- Dieter, G.; Pierre, V.D.; Walter, V.; Peggy, B.; Bea, J.; Patrick, S.; Reinout, N. SARS-CoV-2 vaccine breakthrough infections in health care workers of a large Belgian hospital network. In Infection Control and Hospital Epidemiology; Cambridge University Press: Cambridge, UK, 2021; p. 1. [Google Scholar]
- Oster, Y.; Benenson, S.; Nir-Paz, R.; Buda, I.; Cohen, M.J. The effect of a third BNT162b2 vaccine on breakthrough infections in health care workers: A cohort analysis. Clin. Microbiol. Infect. 2022, 28, 735.e1–735.e3. [Google Scholar] [CrossRef]
- Katz, M.A.; Harlev, E.B.; Chazan, B.; Chowers, M.; Greenberg, D.; Peretz, A.; Tshori, S.; Levy, J.; Yacobi, M.; Hirsch, A.; et al. Early effectiveness of BNT162b2 COVID-19 vaccine in preventing SARS-CoV-2 infection in healthcare personnel in six Israeli hospitals (CoVEHPI). Vaccine 2022, 40, 512–520. [Google Scholar] [CrossRef]
- Naito, T.; Yan, Y.; Tabe, Y.; Seyama, K.; Deshpande, G.A. Real-world evidence for the effectiveness and breakthrough of BNT162b2 mRNA COVID-19 vaccine at a medical center in Japan. Hum. Vaccines Immunother. 2021, 18, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Bassi, J.; Giannini, O.; Silacci-Fregni, C.; Pertusini, L.; Hitz, P.; Terrot, T.; Franzosi, Y.; Muoio, F.; Saliba, C.; Meury, M.; et al. Poor neutralization and rapid decay of antibodies to SARS-CoV-2 variants in vaccinated dialysis patients. PLoS ONE 2022, 17, e0263328. [Google Scholar] [CrossRef] [PubMed]
- Colavita, F.; Meschi, S.; Gruber, C.E.M.; Rueca, M.; Vairo, F.; Matusali, G.; Lapa, D.; Giombini, E.; De Carli, G.; Spaziante, M.; et al. Virological and Serological Characterisation of SARS-CoV-2 Infections Diagnosed After mRNA BNT162b2 Vaccination Between December 2020 and March 2021. Front. Med. 2022, 8, 815870. [Google Scholar] [CrossRef] [PubMed]
- Rovida, F.; Cassaniti, I.; Paolucci, S.; Percivalle, E.; Sarasini, A.; Piralla, A.; Giardina, F.; Sammartino, J.C.; Ferrari, A.; Bergami, F.; et al. SARS-CoV-2 vaccine breakthrough infections with the alpha variant are asymptomatic or mildly symptomatic among health care workers. Nat. Commun. 2021, 12, 6032. [Google Scholar] [CrossRef] [PubMed]
- Benning, L.; Morath, C.; Bartenschlager, M.; Reineke, M.; Töllner, M.; Nusshag, C.; Kälble, F.; Reichel, P.; Schaier, M.; Klein, K.; et al. Neutralizing antibody activity against the B.1.617.2 (delta) variant 8 months after two-dose vaccination with BNT162b2 in health care workers. Clin. Microbiol. Infect. 2022. [Google Scholar] [CrossRef]
- Evans, J.P.; Zeng, C.; Carlin, C.; Lozanski, G.; Saif, L.J.; Oltz, E.M.; Gumina, R.J.; Liu, S.-L. Neutralizing antibody responses elicited by SARS-CoV-2 mRNA vaccination wane over time and are boosted by breakthrough infection. Sci. Transl. Med. 2022, 14, eabn8057. [Google Scholar] [CrossRef]
- Rümke, L.W.; Groenveld, F.C.; Van Os, Y.M.G.; Praest, P.; Tanja, A.A.N.; De Jong, D.T.C.M.; Symons, J.; Schuurman, R.; Reinders, T.; Hofstra, L.M.; et al. In-depth Characterization of Vaccine Breakthrough Infections with SARS-CoV-2 among Health Care Workers in a Dutch Academic Medical Center. Open Forum Infect. Dis. 2022, 9, ofab553. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter VS. Parameter | t3m | t10m | ||
|---|---|---|---|---|
| Spearman r (p-Value) | Pearson R² | Spearman r (p-Value) | Pearson R² | |
| Anti-S IgG vs. anti-S IgA | 0.6218 (p < 0.0001) | 0.3438 | 0.4803 (p < 0.0001) | 0.1854 |
| Anti-S IgG vs. anti-RBD IgG | 0.8988 (p < 0.0001) | 0.6325 | 0.5074 (p = 0.005) | 0.5295 |
| Anti-S IgA vs. anti-RBD IgG | 0.5502 (p < 0.0001) | 0.2162 | 0.3728 (p = 0.0464) | 0.0067 |
| Anti-S IgG vs. RBD-ACE2% IH | 0.8421 (p < 0.0001) | 0.5941 | 0.9162 (p < 0.0001) | 0.7336 |
| Anti-S IgG vs. qAC50 D614G | 0.8903 (p < 0.0001) | 0.6358 | 0.8486 (p < 0.0001) | 0.6496 |
| Anti-S IgG vs. qAC50 delta | 0.7462 (p < 0.0001) | 0.3699 | 0.6357 (p = 0.0046) | 0.1802 |
| Anti-RBD IgG vs. RBD-ACE2% IH | 0.8194 (p < 0.0001) | 0.3997 | 0.4793 (p = 0.0085) | 0.1352 |
| Anti-RBD IgG vs. qAC50 D614G | 0.8127 (p < 0.0001) | 0.5336 | 0.3210 (p = 0.1025) | 0.1893 |
| Anti-RBD IgG vs. qAC50 delta | 0.6784 (p < 0.0001) | 0.3757 | 0.4259 (p = 0.1007) | 0.0178 |
| RBD-ACE2% IH vs. qAC50 D614G | 0.8534 (p < 0.0001) | 0.4578 | 0.8853 (p < 0.0001) | 0.6765 |
| RBD-ACE2% IH vs. qAC50 delta | 0.8087 (p < 0.0001) | 0.4200 | 0.8475 (p < 0.0001) | 0.3844 |
| Parameter VS. Parameter | t3m | t10m | ||
|---|---|---|---|---|
| Spearman r (p-Value) | Pearson R² | Spearman r (p-Value) | Pearson R² | |
| IFN-γ vs. CD69+ TC cells | 0.7649 (p < 0.0001) | 0.3891 | 0.5030 (p = 0.0144) | 0.2360 |
| IFN-γ vs. CD69+ TH cells | 0.6961 (p = 0.0005) | 0.6568 | 0.7915 (p < 0.0001) | 0.5962 |
| IFN-γ vs. CD40L+ TH cells | 0.6171 (p = 0.0029) | 0.6344 | 0.4070 (p = 0.0539) | 0.0898 |
| Immune Parameter | t3m | ||
|---|---|---|---|
| BTI Group (Mean ± SD) * | Non-BTI Group (Mean ± SD) * | p–Values ** | |
| Humoral immune response | |||
| Anti-S IgG (index) | 21.83 ± 11.33 | 26.79 ± 8.508 | p = 0.2418 |
| Anti-S IgA (index) | 1.740 ± 0.959 | 2.948 ± 1.492 | p = 0.0077 |
| Anti-RBD IgG (BAU/mL) | 773.2 ± 245.4 | 981.6 ± 555.0 | p = 0.5883 |
| RBD-ACE2 inhibition (% IH) | 75.99 ± 32.58 | 88.19 ± 11.37 | p = 0.1888 |
| qAC50 D614G VoC | 208.1 ± 64.28 | 297.7 ± 186.8 | p = 0.3213 |
| qAC50 delta VoC | 62.86 ± 13.08 | 110.6 ± 54.69 | p = 0.0173 |
| Cellular immune response | |||
| IFN-γ (mIU/mL) | 1349 ± 936.7 | 1950 ± 2094 | p = 0.7394 |
| CD69+ TC cells (% of parent) | 17.64 ± 7.325 | 18.70 ± 8.107 | p = 0.8501 |
| CD69+ TH cells (% of parent) | 11.96 ± 6.834 | 16.87 ± 7.717 | p = 0.1570 |
| CD40L+ TH cells (% of parent) | 0.476 ± 0.451 | 0.551 ± 0.571 | p = 0.6058 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calcoen, B.; Callewaert, N.; Vandenbulcke, A.; Kerstens, W.; Imbrechts, M.; Vercruysse, T.; Dallmeier, K.; Van Weyenbergh, J.; Maes, P.; Bossuyt, X.; et al. High Incidence of SARS-CoV-2 Variant of Concern Breakthrough Infections Despite Residual Humoral and Cellular Immunity Induced by BNT162b2 Vaccination in Healthcare Workers: A Long-Term Follow-Up Study in Belgium. Viruses 2022, 14, 1257. https://doi.org/10.3390/v14061257
Calcoen B, Callewaert N, Vandenbulcke A, Kerstens W, Imbrechts M, Vercruysse T, Dallmeier K, Van Weyenbergh J, Maes P, Bossuyt X, et al. High Incidence of SARS-CoV-2 Variant of Concern Breakthrough Infections Despite Residual Humoral and Cellular Immunity Induced by BNT162b2 Vaccination in Healthcare Workers: A Long-Term Follow-Up Study in Belgium. Viruses. 2022; 14(6):1257. https://doi.org/10.3390/v14061257
Chicago/Turabian StyleCalcoen, Bas, Nico Callewaert, Aline Vandenbulcke, Winnie Kerstens, Maya Imbrechts, Thomas Vercruysse, Kai Dallmeier, Johan Van Weyenbergh, Piet Maes, Xavier Bossuyt, and et al. 2022. "High Incidence of SARS-CoV-2 Variant of Concern Breakthrough Infections Despite Residual Humoral and Cellular Immunity Induced by BNT162b2 Vaccination in Healthcare Workers: A Long-Term Follow-Up Study in Belgium" Viruses 14, no. 6: 1257. https://doi.org/10.3390/v14061257
APA StyleCalcoen, B., Callewaert, N., Vandenbulcke, A., Kerstens, W., Imbrechts, M., Vercruysse, T., Dallmeier, K., Van Weyenbergh, J., Maes, P., Bossuyt, X., Zapf, D., Dieckmann, K., Callebaut, K., Thibaut, H. J., Vanhoorelbeke, K., De Meyer, S. F., Maes, W., & Geukens, N. (2022). High Incidence of SARS-CoV-2 Variant of Concern Breakthrough Infections Despite Residual Humoral and Cellular Immunity Induced by BNT162b2 Vaccination in Healthcare Workers: A Long-Term Follow-Up Study in Belgium. Viruses, 14(6), 1257. https://doi.org/10.3390/v14061257