
Insight into the Epidemiology and Evolutionary History of Novel Goose Astrovirus-Associated Gout in Goslings in Southern China

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
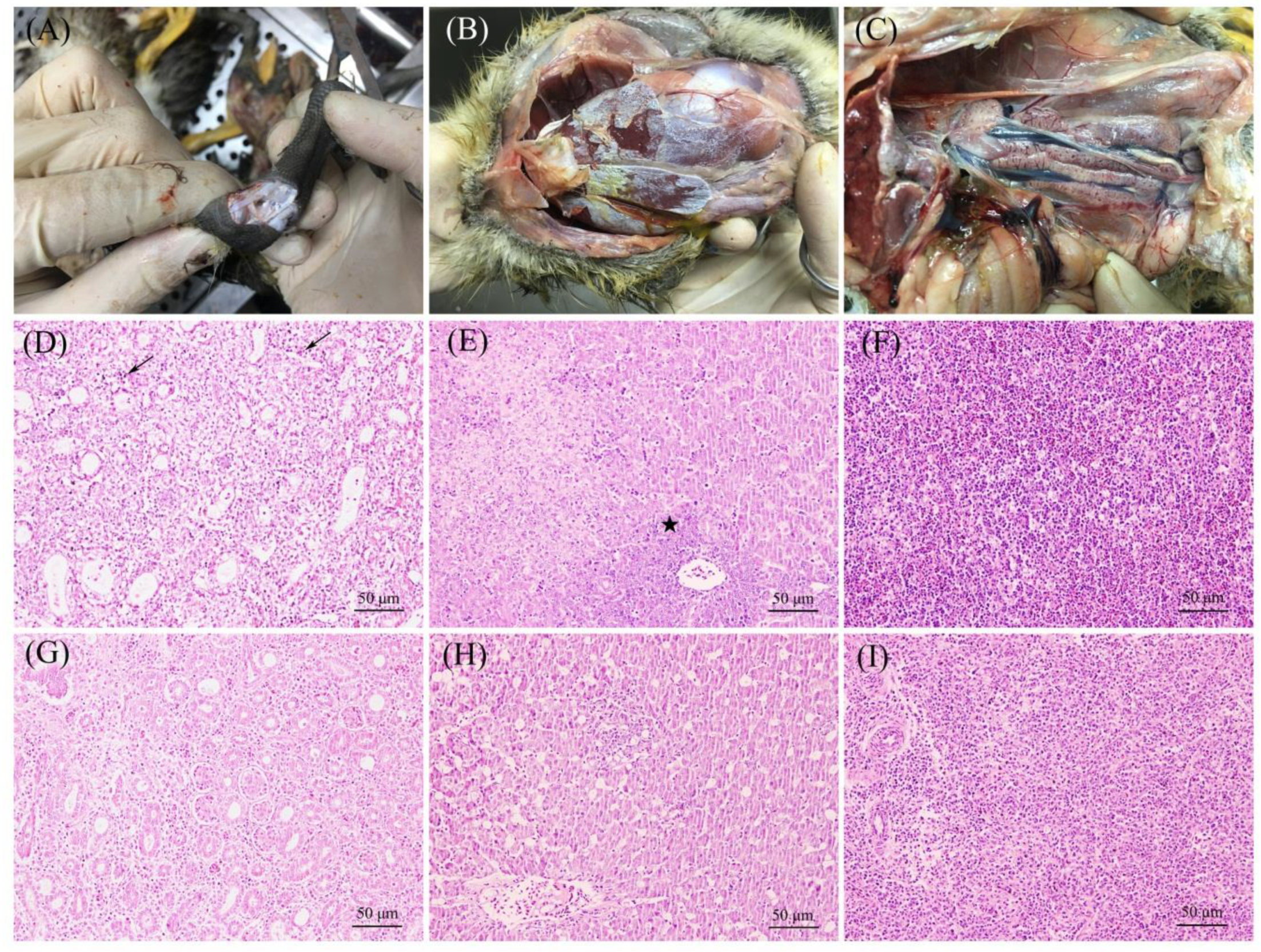
2.1. Clinical Signs Description and Sample Collection
2.2. Sample Processing, Virus Detection and Genome Sequencing
2.3. GoAstV Genome, Molecular Characteristics, Recombination and Phylogenetic Analysis
2.4. Evolutionary Dynamics and the Evolutionary History of GoAstV
3. Results
3.1. Clinical Signs and Virus Detection
3.2. Genome Sequence and Molecular Characteristics Analysis
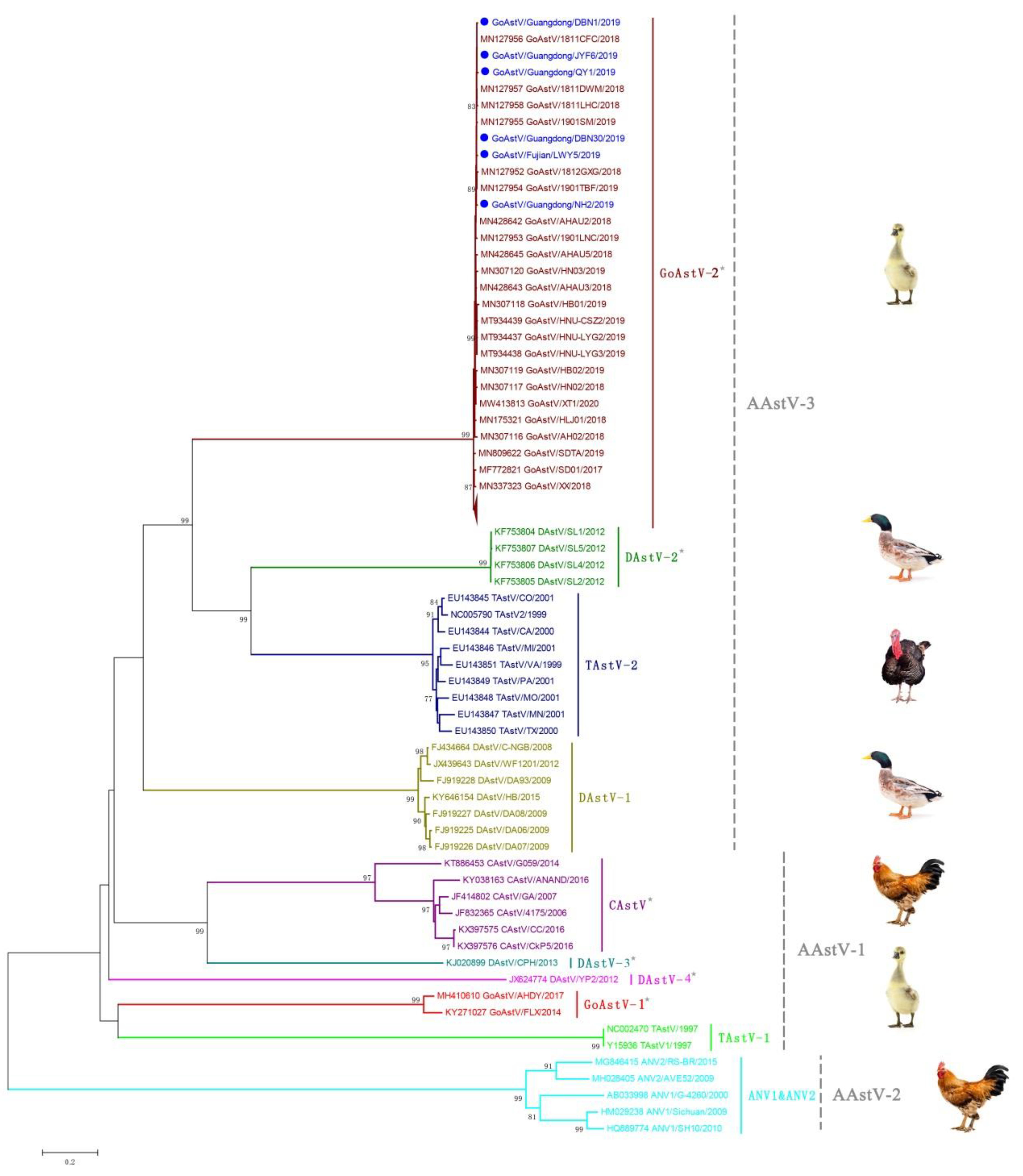
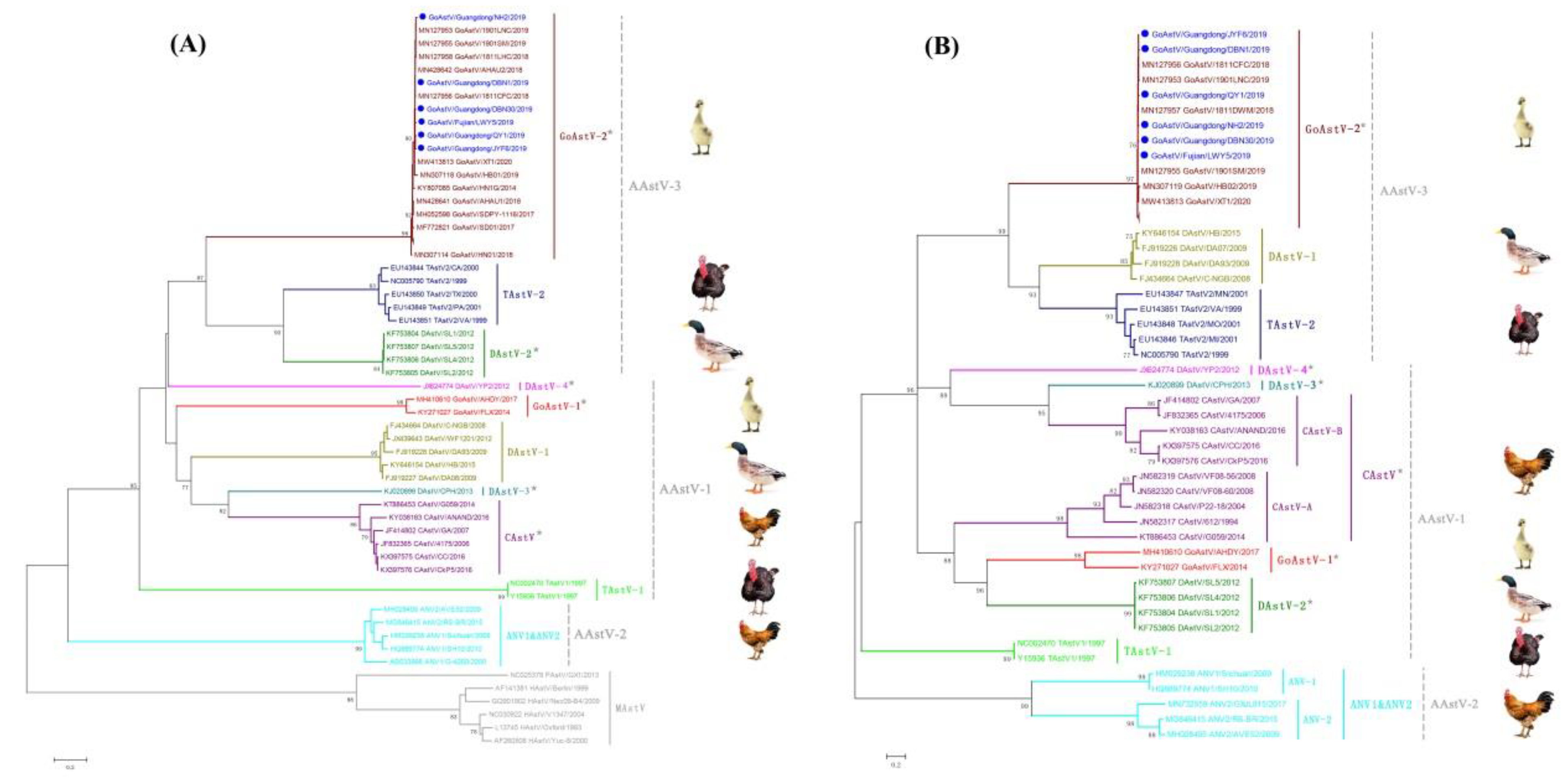
3.3. Phylogenetic Analysis and Classification of GoAstV
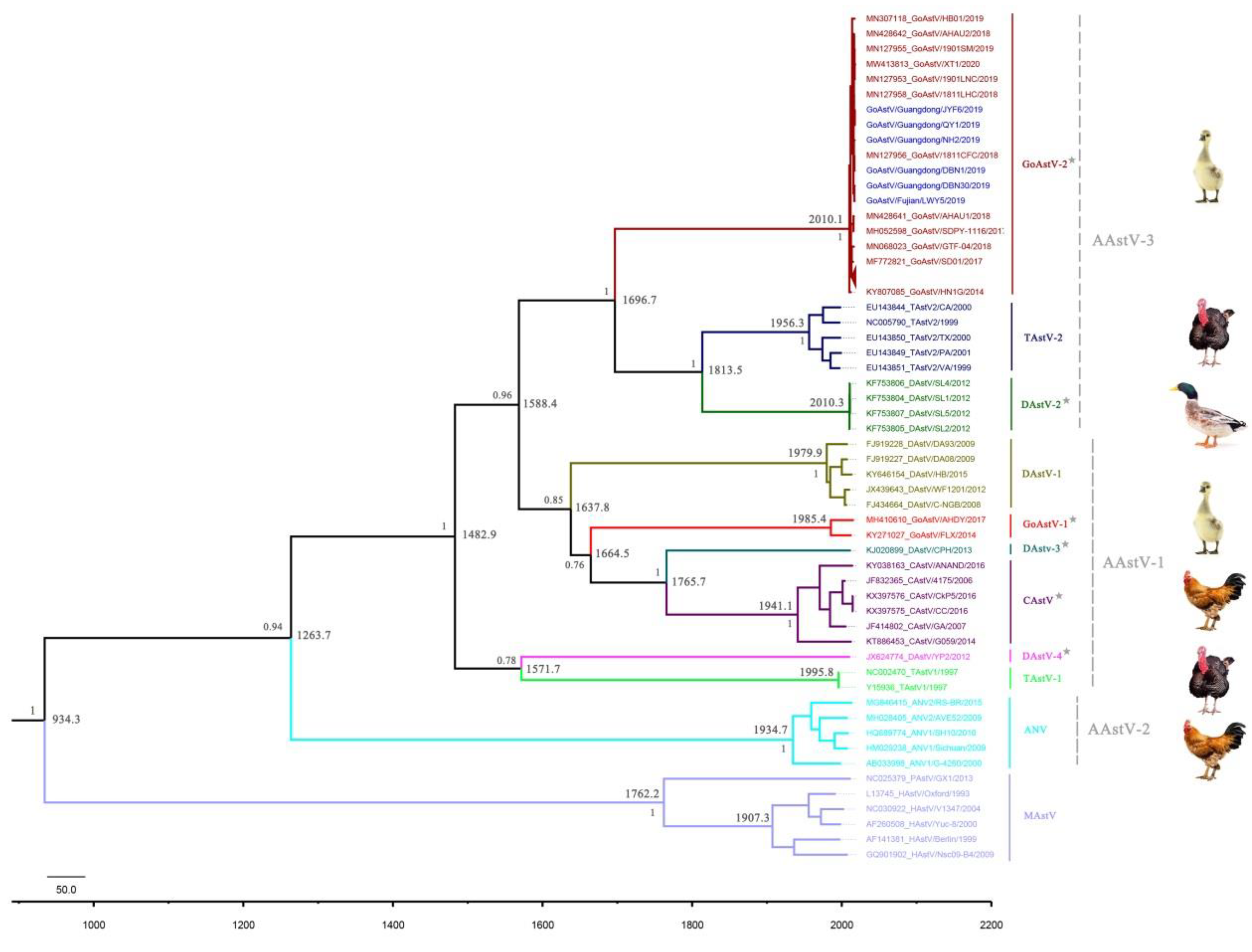
3.4. Divergence Time and Evolutionary History of GoAstV-2
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perot, P.; Lecuit, M.; Eloit, M. Astrovirus Diagnostics. Viruses 2017, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortez, V.; Meliopoulos, V.A.; Karlsson, E.A.; Hargest, V.; Johnson, C.; Schultz-Cherry, S. Astrovirus Biology and Pathogenesis. Annu. Rev. Virol. 2017, 4, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Donato, C.; Vijaykrishna, D. The Broad Host Range and Genetic Diversity of Mammalian and Avian Astroviruses. Viruses 2017, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, C.F.; DuBois, R.M. The Astrovirus Capsid: A Review. Viruses 2017, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Smyth, V.J. A Review of the Strain Diversity and Pathogenesis of Chicken Astrovirus. Viruses 2017, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Guix, S.; Krishna, N.K.; Méndez, E.; Monroe, S.S.; Pantin-Jackwood, M.; Schultz-Cherry, S. Family—Astroviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: San Diego, CA, USA, 2012; pp. 953–959. [Google Scholar]
- Chu, D.K.; Leung, C.Y.; Perera, H.K.; Ng, E.M.; Gilbert, M.; Joyner, P.H.; Grioni, A.; Ades, G.; Guan, Y.; Peiris, J.S.; et al. A novel group of avian astroviruses in wild aquatic birds. J. Virol. 2012, 86, 13772–13778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.; Wang, X.; Ning, K.; Liu, N.; Zhang, D. Genetic characterization of a new astrovirus in goslings suffering from gout. Arch. Virol. 2018, 163, 2865–2869. [Google Scholar] [CrossRef]
- Niu, X.; Tian, J.; Yang, J.; Jiang, X.; Wang, H.; Chen, H.; Yi, T.; Diao, Y. Novel Goose Astrovirus Associated Gout in Gosling, China. Vet. Microbiol. 2018, 220, 53–56. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, D.; Li, T.; Zhou, H.; Liu, X.; Wang, X.; Lu, H.; Gao, W.; Wang, Y.; Zou, X.; et al. An emerging novel goose astrovirus associated with gosling gout disease, China. Emerg. Microbes Infect. 2018, 7, 152–159. [Google Scholar] [CrossRef]
- Zhang, Q.; Cao, Y.; Wang, J.; Fu, G.; Sun, M.; Zhang, L.; Meng, L.; Cui, G.; Huang, Y.; Hu, X.; et al. Isolation and characterization of an astrovirus causing fatal visceral gout in domestic goslings. Emerg. Microbes Infect. 2018, 7, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Tian, J.; Tang, Y.; Diao, Y. Isolation and genomic characterization of gosling gout caused by a novel goose astrovirus. Transbound. Emerg. Dis. 2018, 65, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, B.; Yan, M.; Diao, Y.; Tang, Y. First report of a novel goose astrovirus outbreak in Cherry Valley ducklings in China. Transbound. Emerg. Dis. 2020, 67, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Yang, J.; Wang, Y.; Chen, H.; Diao, Y.; Tang, Y. Isolation and characterization of a duck-origin goose astrovirus in China. Emerg. Microbes Infect. 2020, 9, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Wang, H.; Wei, L.; Zhang, M.; Yang, J.; Chen, H.; Tang, Y.; Diao, Y. Epidemiological investigation of H9 avian influenza virus, Newcastle disease virus, Tembusu virus, goose parvovirus and goose circovirus infection of geese in China. Transbound. Emerg. Dis. 2018, 65, e304–e316. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Rambaut, A.; Shapiro, B.; Pybus, O.G. Bayesian coalescent inference of past population dynamics from molecular sequences. Mol. Biol. Evol. 2005, 22, 1185–1192. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Fang, B.; Liu, Y.; Cai, M.; Jun, J.; Ma, J.; Bu, D.; Wang, L.; Zhou, P.; Wang, H.; et al. Newly emerged porcine enteric alphacoronavirus in southern China: Identification, origin and evolutionary history analysis. Infect. Genet. Evol. 2018, 62, 179–187. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, F.; Liu, N.; Yang, L.; Zhang, D. Complete genome sequence of a novel avastrovirus in goose. Arch. Virol. 2017, 162, 2135–2139. [Google Scholar] [CrossRef]
- Bosch, A.; Pinto, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Yang, J.; He, D.; Diao, Y.; Tang, Y. Evidence of vertical transmission of novel astrovirus virus in goose. Vet. Microbiol. 2020, 244, 108657. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Mu, X.; Xu, X.; Ji, J.; Ma, K.; Leng, C.; Yao, L.; Kan, Y.; Bi, Y.; Xie, Q. Extensive genetic heterogeneity and molecular characteristics of emerging astroviruses causing fatal gout in goslings. Poult. Sci. 2022, 101, 101888. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Deng, T.; Song, Y.; Liu, J.; Jiang, Z.; Peng, Z.; Guo, Y.; Yang, L.; Qiao, H.; Xia, Y.; et al. Identification and genomic characterization of emerging goose astrovirus in central China, 2020. Transbound. Emerg. Dis. 2022, 69, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, H.; Wei, Q.; Xie, Q.; Zeng, Y.; Wu, C.; Yang, Q.; Tan, J.; Tan, M.; Kang, Z. Isolation and phylogenetic analysis of goose astrovirus type 1 from goslings with gout in Jiangxi province, China. Poult. Sci. 2022, 101, 101800. [Google Scholar] [CrossRef]
- Strain, E.; Kelley, L.A.; Schultz-Cherry, S.; Muse, S.V.; Koci, M.D. Genomic analysis of closely related astroviruses. J. Virol. 2008, 82, 5099–6103. [Google Scholar] [CrossRef] [Green Version]
- Babkin, I.V.; Tikunov, A.Y.; Zhirakovskaia, E.V.; Netesov, S.V.; Tikunova, N.V. High evolutionary rate of human astrovirus. Infect. Genet. Evol. 2012, 12, 435–442. [Google Scholar] [CrossRef]
- Li, J.Y.; Hu, W.Q.; Liu, T.N.; Zhang, H.H.; Opriessnig, T.; Xiao, C.T. Isolation and evolutionary analyses of gout-associated goose astrovirus causing disease in experimentally infected chickens. Poult. Sci. 2021, 100, 543–552. [Google Scholar] [CrossRef]
- Wu, W.; Xu, R.; Lv, Y.; Bao, E. Goose astrovirus infection affects uric acid production and excretion in goslings. Poult. Sci. 2020, 99, 1967–1974. [Google Scholar] [CrossRef]
- Wang, Z.; Li, L.; Liu, P.; Wang, C.; Lu, Q.; Liu, L.; Yang, Y.; Luo, Q.; Shao, H. Host innate immune responses of geese infected with goose origin nephrotic astrovirus. Microb. Pathog. 2021, 152, 104753–104763. [Google Scholar] [CrossRef]
- Wu, W.; Qiu, S.; Huang, H.; Xu, R.; Bao, E.; Lv, Y. Immune-related gene expression in the kidneys and spleens of goslings infected with goose nephritic astrovirus. Poult. Sci. 2021, 100, 100990. [Google Scholar] [CrossRef]
- Ren, D.; Li, T.; Zhang, X.; Yao, X.; Gao, W.; Xie, Q.; Zhang, J.; Shao, H.; Wan, Z.; Qin, A.; et al. OASL Triggered by Novel Goose Astrovirus via ORF2 Restricts Its Replication. J. Virol. 2020, 94, e01767-20. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Primer Sequences (5′-3′) | Product Length | Purpose |
|---|---|---|---|
| GoAstV-D: F | TATGCTCCTACTAAATGGGAC | 540 bp | virus detection |
| GoAstV-D: R | ACCAATGAGCCTAGATACTCG | ||
| GoAstV: F1 | CCCAAAACCCCTTTTCCTCACT | 1380 bp | genome amplification |
| GoAstV: R1 | TAATTGTCAAGGCTGTAGACCAC | ||
| GoAstV: F2 | GGTTGAGGTTTCTTATGCGGTA | 1286 bp | |
| GoAstV: R2 | CGCCCCTTTCTTCCATTGCT | ||
| GoAstV: F3 | AAGAAGGTGCGGAAGAGTGG | 1250 bp | |
| GoAstV: R3 | GGGGCCTATTTTGCACAGT | ||
| GoAstV: F4 | TGAACAATGTGGCTGGGGTGA | 1374 bp | |
| GoAstV: R4 | TATTTATGCTTCCGGTCGAGT | ||
| GoAstV: F5 | CTGGACCACCCGTTAATGACA | 1198 bp | |
| GoAstV: R5 | CTTGACCTGGATTCTGCCTGT | ||
| GoAstV: F6 | GGCTGCAAATACAGTTAAGCTTG | 1102 bp | |
| GoAstV: R6 | CACGCGATTTGTGTGGGTGAC |
| Astrovirus (GenBank Accession No.) | Genome Size | G + C Content (%) | Percent Identity (%) to GoAstV/Guangdong/QY1/2019 | |||
|---|---|---|---|---|---|---|
| Genome (nt) | ORF1a (aa) | RdRp (aa) | Cap (aa) | |||
| GoAstV/Guangdong/QY1/2019 | 7033 bp | 43.4 | NA a | NA | NA | NA |
| TAstV-2/CO/2001 (EU143845) | 7343 bp | 43.5 | 63.0 | 59.1 | 68.3 | 56.3 |
| DAstv-2/SL1/2012 (KF753804) | 7319 bp | 42.0 | 60.7 | 58.5 | 68.0 | 40.1 |
| DAstV-1/HB/2015 (KY646154) | 7754 bp | 41.8 | 60.0 | 48.5 | 63.9 | 56.2 |
| DAstV-3/CPH/2013 (KJ020899) | 7463 bp | 43.0 | 58.9 | 50.0 | 65.4 | 37.5 |
| GoAstV-1/FLX/2014 (KY271027) | 7299 bp | 41.6 | 58.0 | 47.4 | 61.0 | 42.5 |
| DAstV-4/YP2/2012 (JX624774) | 7287 bp | 43.2 | 56.9 | 42.3 | 61.3 | 34.3 |
| CAstV-A/G059/2014 (KT886453) | 7382 bp | 42.5 | 58.2 | 49.2 | 65.0 | 38.9 |
| CAstV-B/4175/2006 (JF832365) | 7008 bp | 42.9 | 58.0 | 49.1 | 57.9 | 32.4 |
| TAstV-1/1997 (NC002470) | 7003 bp | 43.5 | 54.6 | 40.4 | 56.8 | 41.3 |
| ANV/SH10/2019 (HQ889774) | 6928 bp | 44.8 | 52.2 | 27.0 | 53.0 | 28.0 |
| HAstV/Berlin/1999 (AF141381) | 6815 bp | 44.1 | 49.0 | 17.7 | 38.6 | 18.7 |
| AAstV Species | Average p-Distance between Different AAstV Species (Mean ± SE) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| GoAstV-1 * | GoAstV-2 * | TAstV-1 | TAstV-2 | DAstV-1 | DAstV-2 * | DAstV-3 * | DAstV-4 * | CAstV * | ANV | |
| GoAstV-2 * | 0.609 ± 0.021 | |||||||||
| TAstV-1 | 0.565 ± 0.20 | 0.594 ± 0.022 | ||||||||
| TAstV-2 | 0.595 ± 0.021 | 0.403 ± 0.021 | 0.607 ± 0.020 | |||||||
| DAstV-1 | 0.595 ± 0.022 | 0.399 ± 0.021 | 0.615 ± 0.020 | 0.261 ± 0.018 | ||||||
| DAstv-2 * | 0.424 ± 0.021 | 0.592 ± 0.021 | 0.590 ± 0.020 | 0.588 ± 0.021 | 0.597 ± 0.021 | |||||
| DAstV-3 * | 0.611 ± 0.021 | 0.624 ± 0.020 | 0.624 ± 0.022 | 0.597 ± 0.021 | 0.580 ± 0.021 | 0.595 ± 0.021 | ||||
| DAstV-4 * | 0.605 ± 0.021 | 0.655 ± 0.021 | 0.632 ± 0.019 | 0.624 ± 0.022 | 0.6260 ± 022 | 0.595 ± 0.021 | 0.517 ± 0.021 | |||
| CAstV | 0.494 ± 0.021 | 0.603 ± 0.021 | 0.592 ± 0.021 | 0.565 ± 0.020 | 0.582 ± 0.020 | 0.500 ± 0.021 | 0.592 ± 0.021 | 0.588 ± 0.021 | ||
| ANV | 0.695 ± 0.019 | 0.709 ± 0.020 | 0.588 ± 0.020 | 0.719 ± 0.020 | 0.721 ± 0.020 | 0.708 ± 0.020 | 0.723 ± 0.019 | 0.712 ± 0.020 | 0.702 ± 0.020 | |
| HAstV | 0.761 ± 0.018 | 0.823 ± 0.016 | 0.796 ± 0.017 | 0.786 ± 0.017 | 0.784 ± 0.018 | 0.788 ± 0.017 | 0.786 ± 0.017 | 0.773 ± 0.017 | 0.777 ± 0.017 | 0.755 ± 0.016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, X.; Hou, Z.; Liu, W.; Cao, N.; Liang, Y.; Li, B.; Jiang, D.; Li, W.; Xu, D.; Tian, Y.; et al. Insight into the Epidemiology and Evolutionary History of Novel Goose Astrovirus-Associated Gout in Goslings in Southern China. Viruses 2022, 14, 1306. https://doi.org/10.3390/v14061306
Fu X, Hou Z, Liu W, Cao N, Liang Y, Li B, Jiang D, Li W, Xu D, Tian Y, et al. Insight into the Epidemiology and Evolutionary History of Novel Goose Astrovirus-Associated Gout in Goslings in Southern China. Viruses. 2022; 14(6):1306. https://doi.org/10.3390/v14061306
Chicago/Turabian StyleFu, Xinliang, Zhanpeng Hou, Wenjun Liu, Nan Cao, Yu Liang, Bingxin Li, Danli Jiang, Wanyan Li, Danning Xu, Yunbo Tian, and et al. 2022. "Insight into the Epidemiology and Evolutionary History of Novel Goose Astrovirus-Associated Gout in Goslings in Southern China" Viruses 14, no. 6: 1306. https://doi.org/10.3390/v14061306
APA StyleFu, X., Hou, Z., Liu, W., Cao, N., Liang, Y., Li, B., Jiang, D., Li, W., Xu, D., Tian, Y., & Huang, Y. (2022). Insight into the Epidemiology and Evolutionary History of Novel Goose Astrovirus-Associated Gout in Goslings in Southern China. Viruses, 14(6), 1306. https://doi.org/10.3390/v14061306