
Influenza Virus Infections in Polarized Cells


Abstract
:1. Introduction
2. Virus–Receptor Attachment and Viral Entry
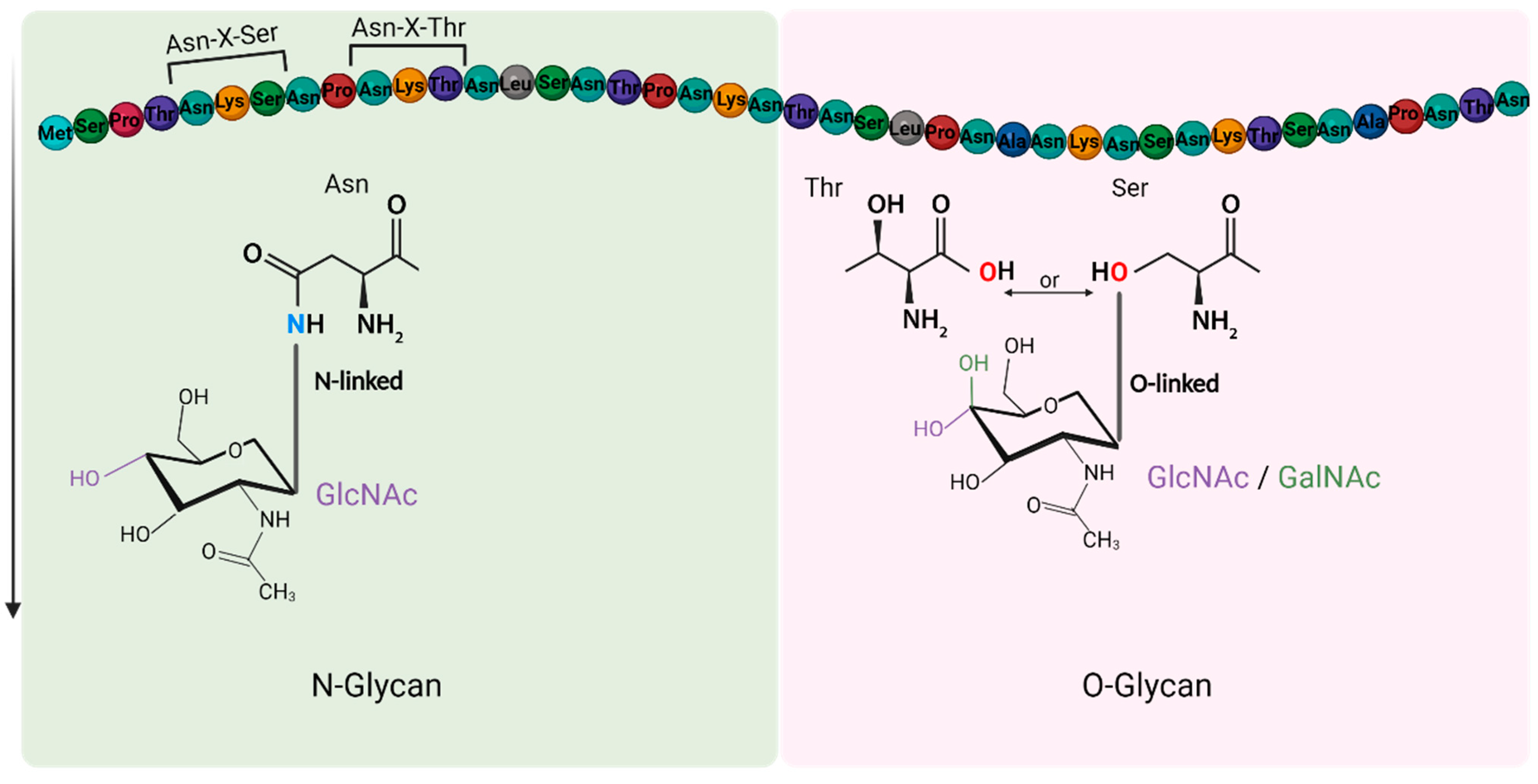
2.1. Influenza Receptor-Binding Site
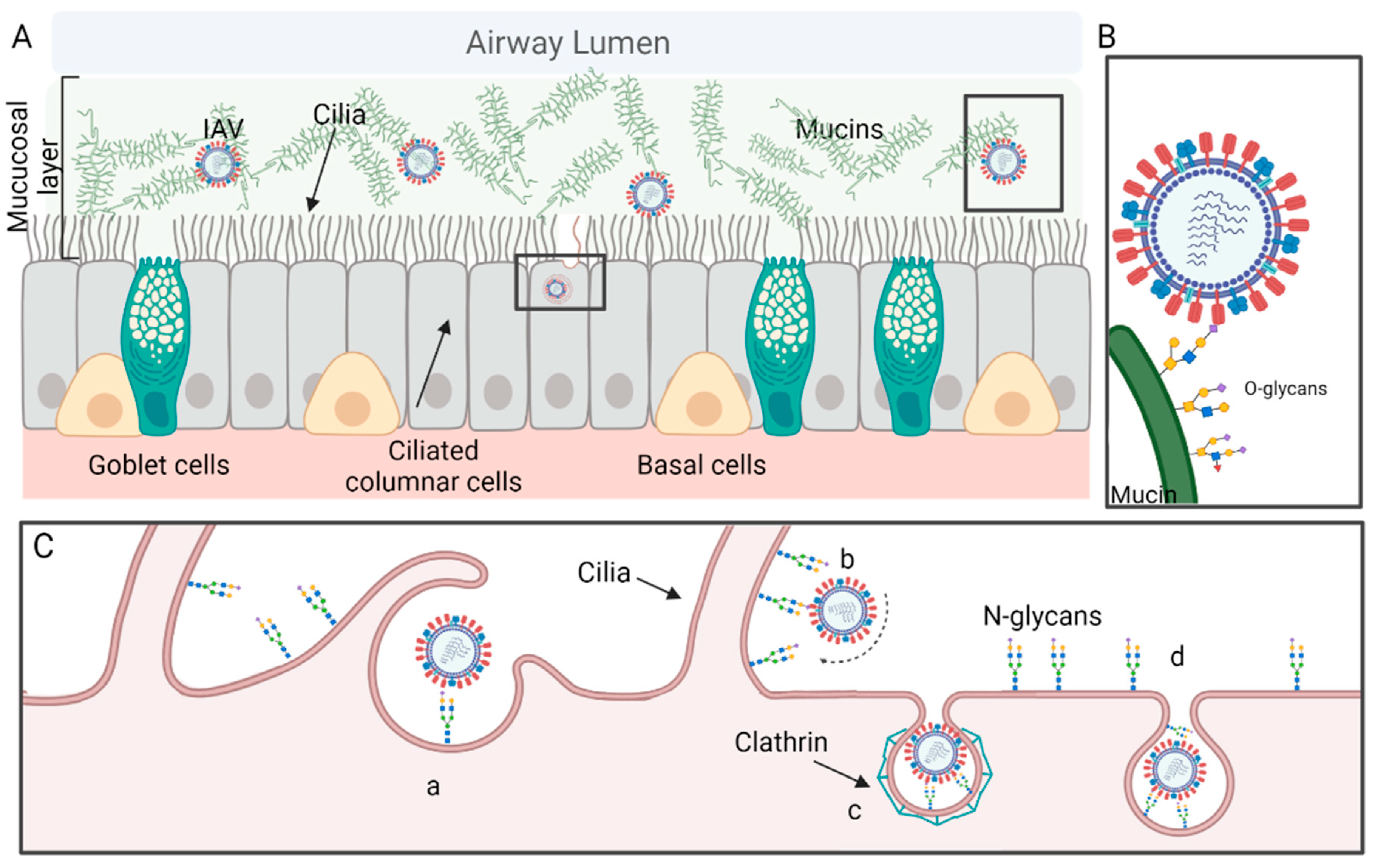
2.2. The Respiratory Tract
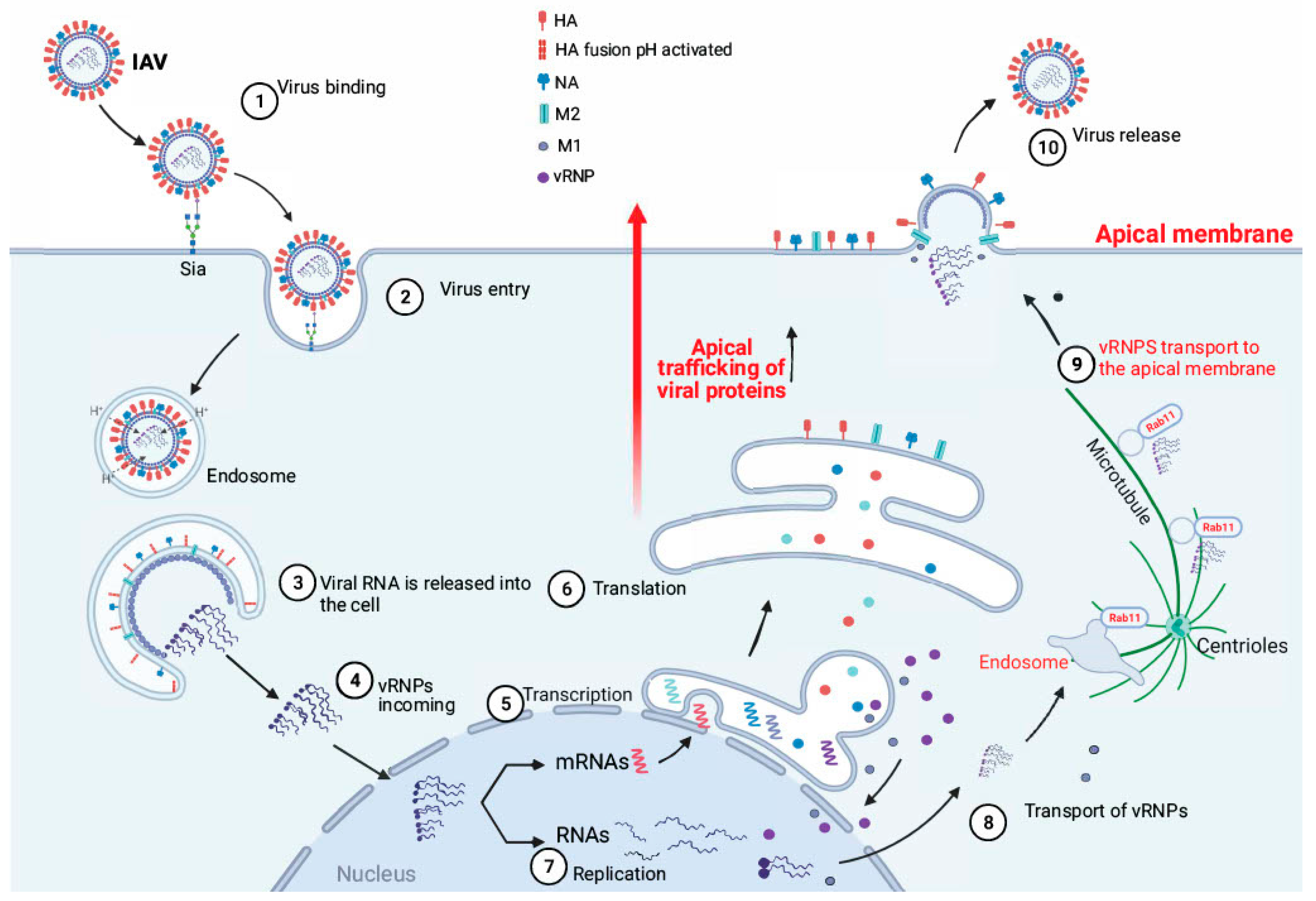
2.3. Attachment and Viral Entry
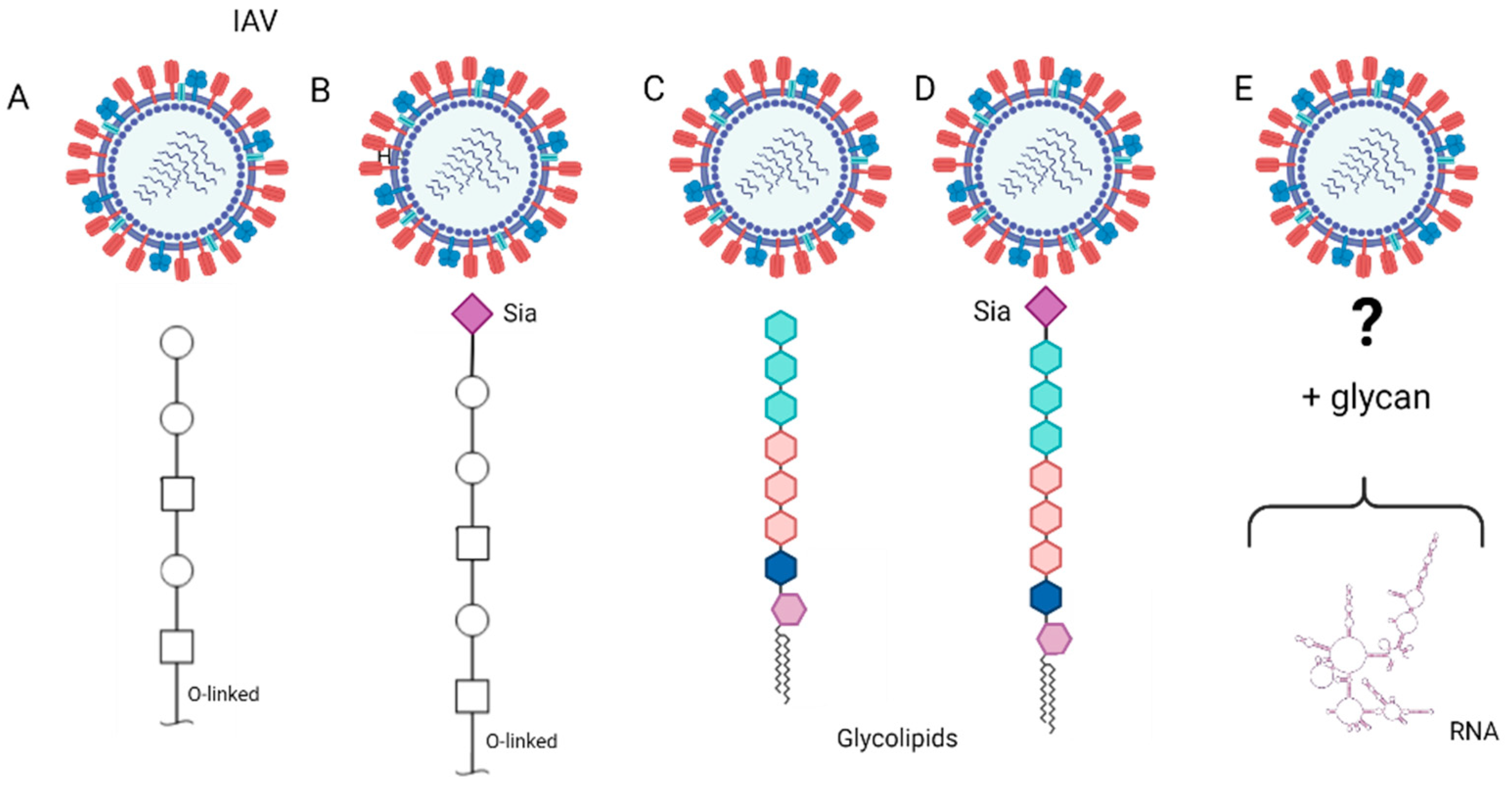
3. Receptor of IAVs
3.1. Sialylated N-Glycans Receptors
3.2. Other Glycan Receptors

4. Influenza Virus Infection Can Change the Complexity and Polarity of Cells
5. Importance of Studying Influenza Virus Infection in Polarized Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture * | Host | Polarization | Cell Type | α-2,6 Sia | α-2,3 Sia | References |
|---|---|---|---|---|---|---|
| HAE | Human | Y | Ciliated | ++ | ++ | [99,100] |
| Nonciliated | +++ | + | ||||
| hTEC | Human | Y | Ciliated | ++ | +++ | [24] |
| Nonciliated | +++ | - | ||||
| HNTEC 3D | Human | Y | - | +++ | ++ | [101] |
| Ex vivo | Human | Y | Ciliated | ++ | +++ | [24,100] |
| Nonciliated | +++ | - | ||||
| Basal | - | ++ | ||||
| PBEC | Swine | Y | Ciliated | + | + | [102,103] |
| Nonciliated | +++ | - | ||||
| Basal | + | +++ | ||||
| NE | Swine | Y | Ciliated | ++ | - | [103] |
| TE | Swine | Y | Unknown | ++ | - | |
| LE | Swine | Y | Unknown | ++ | +/− | |
| FTE | Ferret | Y | Ciliated | +++ | - | |
| Nonciliated | + | +++ | [104] | |||
| PD | Human | N | - | +++ | + | [99] |
| HNTEC 2D | Human | N | - | ++ | ++ | [101] |
| MDCK | Canine | N | - | + | ++ | [105] |
| MDCK-SIAT1 | Canine | N | - | +++ | + | [106,107] |
| MDCK-London | Canine | N | - | ++ | + | [107] |
| hMDCK | Canine | N | - | +++ | - | [108] |
| 16HB14o- | N | - | ++ | +++ | [109] |
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Jin, F.G.; Wang, P.; Wang, M.; Zhu, J.M. Isolation of influenza C virus from pigs and experimental infection of pigs with influenza C virus. J. Gen. Virol. 1983, 64 Pt 1, 177–182. [Google Scholar]
- Kimura, H.; Abiko, C.; Peng, G.; Muraki, Y.; Sugawara, K.; Hongo, S.; Kitame, F.; Mizuta, K.; Numazaki, Y.; Suzuki, H.; et al. Interspecies transmission of influenza C virus between humans and pigs. Virus Res. 1997, 48, 71–79. [Google Scholar] [CrossRef]
- Zhai, S.L.; Zhang, H.; Chen, S.N.; Zhou, X.; Lin, T.; Liu, R.X.; Lv, D.H.; Wen, X.H.; Wei, W.K.; Wang, D.; et al. Influenza D Virus in Animal Species in Guangdong Province, Southern China. Emerg. Infect. Dis. 2017, 23, 1392–1396. [Google Scholar] [CrossRef] [PubMed]
- Salem, E.; Cook, E.A.J.; Lbacha, H.A.; Oliva, J.; Awoume, F.; Aplogan, G.L.; Hymann, E.C.; Muloi, D.; Deem, S.L.; Alali, S.; et al. Serologic Evidence for Influenza C and D Virus among Ruminants and Camelids, Africa, 1991–2015. Emerg. Infect. Dis. 2017, 23, 1556–1559. [Google Scholar] [CrossRef] [Green Version]
- Quast, M.; Sreeniyasan, C.; Sexton, G.; Nedland, H.; Singrey, A.; Fawcett, L.; Miller, G.; Lauer, D.; Voss, S.; Pollock, S.; et al. Serological evidence for the presence of influenza D virus in small ruminants. Vet. Microbiol. 2015, 180, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Nedland, H.; Wollman, J.; Sreenivasan, C.; Quast, M.; Singrey, A.; Fawcett, L.; Christopher-Hennings, J.; Nelson, E.; Kaushik, R.S.; Wang, D.; et al. Serological evidence for the co-circulation of two lineages of influenza D viruses in equine populations of the Midwest United States. Zoonoses Public Health 2018, 65, E148–E154. [Google Scholar] [CrossRef]
- Hause, B.M.; Ducatez, M.; Collin, E.A.; Ran, Z.G.; Liu, R.X.; Sheng, Z.Z.; Armien, A.; Kaplan, B.; Chakravarty, S.; Hoppe, A.D.; et al. Isolation of a Novel Swine Influenza Virus from Oklahoma in 2011 Which Is Distantly Related to Human Influenza C Viruses. PLoS Pathog. 2013, 9, e1003176. [Google Scholar] [CrossRef] [Green Version]
- Hause, B.M.; Collin, E.A.; Liu, R.X.; Huang, B.; Sheng, Z.Z.; Lu, W.X.; Wang, D.; Nelson, E.A.; Li, F. Characterization of a Novel Influenza Virus in Cattle and Swine: Proposal for a New Genus in the Orthomyxoviridae Family. Mbio 2014, 5, e00031-14. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, L.; Eckard, L.; Epperson, W.B.; Long, L.P.; Smith, D.; Huston, C.; Genova, S.; Webby, R.; Wan, X.F. Influenza D virus infection in Mississippi beef cattle. Virology 2015, 486, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzoor, R.; Igarashi, M.; Takada, A. Influenza A Virus M2 Protein: Roles from Ingress to Egress. Int. J. Mol. Sci. 2017, 18, 2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [Green Version]
- Rota, P.A.; Wallis, T.R.; Harmon, M.W.; Rota, J.S.; Kendal, A.P.; Nerome, K. Cocirculation of two distinct evolutionary lineages of influenza type B virus since 1983. Virology 1990, 175, 59–68. [Google Scholar] [CrossRef]
- Kanegae, Y.; Sugita, S.; Endo, A.; Ishida, M.; Senya, S.; Osako, K.; Nerome, K.; Oya, A. Evolutionary pattern of the hemagglutinin gene of influenza B viruses isolated in Japan: Cocirculating lineages in the same epidemic season. J. Virol. 1990, 64, 2860–2865. [Google Scholar] [CrossRef] [Green Version]
- Lafond, K.E.; Porter, R.M.; Whaley, M.J.; Suizan, Z.; Ran, Z.; Aleem, M.A.; Thapa, B.; Sar, B.; Proschle, V.S.; Peng, Z.B.; et al. Global burden of influenza-associated lower respiratory tract infections and hospitalizations among adults: A systematic review and meta-analysis. PLoS Med. 2021, 18, e1003550. [Google Scholar] [CrossRef]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Simonsen, L.; Spreeuwenberg, P.; Lustig, R.; Taylor, R.J.; Fleming, D.M.; Kroneman, M.; Van Kerkhove, M.D.; Mounts, A.W.; Paget, W.J.; Teams, G.C. Global Mortality Estimates for the 2009 Influenza Pandemic from the GLaMOR Project: A Modeling Study. PLoS Med. 2013, 10, e1001558. [Google Scholar] [CrossRef] [Green Version]
- Principi, N.; Scala, A.; Daleno, C.; Esposito, S. Influenza C virus-associated community-acquired pneumonia in children. Influenza Other Resp. 2013, 7, 999–1003. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, Y.; Sugawara, K.; Abiko, C.; Ikeda, T.; Aoki, Y.; Mizuta, K.; Katsushima, N.; Katsushima, F.; Katsushima, Y.; Itagaki, T.; et al. Epidemiological information regarding the periodic epidemics of influenza C virus in Japan (1996–2013) and the seroprevalence of antibodies to different antigenic groups. J. Clin. Virol. 2014, 61, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, S.; Ohizumi, A.; Ohyama, S.; Homma, M. Follow-up study of type C influenza outbreak in a children’s home. Microbiol. Immunol. 1987, 31, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Ramey, A.M.; Poulson, R.L.; Gonzalez-Reiche, A.S.; Perez, D.R.; Stallknecht, D.E.; Brown, J.D. Genomic characterization of H14 subtype Influenza A viruses in new world waterfowl and experimental infectivity in mallards (Anas platyrhynchos). PLoS ONE 2014, 9, e95620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibricevic, A.; Pekosz, A.; Walter, M.J.; Newby, C.; Battaile, J.T.; Brown, E.G.; Holtzman, M.J.; Brody, S.L. Influenza virus receptor specificity and cell tropism in mouse and human airway epithelial cells. J. Virol. 2006, 80, 7469–7480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.; Yatim, S.; Peng, S.; Gunaratne, J.; Hunziker, W.; Ludwig, A. The Mammalian Crumbs Complex Defines a Distinct Polarity Domain Apical of Epithelial Tight Junctions. Curr. Biol. 2020, 30, 2791–2804.e6. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Boulan, E.; Macara, I.G. Organization and execution of the epithelial polarity programme. Nat. Rev. Mol. Cell Biol. 2014, 15, 225–242. [Google Scholar] [CrossRef]
- Skehel, J.J.; Wiley, D.C. Receptor binding and membrane fusion in virus entry: The influenza hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef]
- Arai, Y.; Elgendy, E.M.; Daidoji, T.; Ibrahim, M.S.; Ono, T.; Sriwilaijaroen, N.; Suzuki, Y.; Nakaya, T.; Matsumoto, K.; Watanabe, Y. H9N2 Influenza Virus Infections in Human Cells Require a Balance between Neuraminidase Sialidase Activity and Hemagglutinin Receptor Affinity. J. Virol. 2020, 94, e01210–e01220. [Google Scholar] [CrossRef]
- Guo, H.; Rabouw, H.; Slomp, A.; Dai, M.; van der Vegt, F.; van Lent, J.W.M.; McBride, R.; Paulson, J.C.; de Groot, R.J.; van Kuppeveld, F.J.M.; et al. Kinetic analysis of the influenza A virus HA/NA balance reveals contribution of NA to virus-receptor binding and NA-dependent rolling on receptor-containing surfaces. PLoS Pathog. 2018, 14, e1007233. [Google Scholar] [CrossRef] [Green Version]
- Reiter-Scherer, V.; Cuellar-Camacho, J.L.; Bhatia, S.; Haag, R.; Herrmann, A.; Lauster, D.; Rabe, J.P. Force Spectroscopy Shows Dynamic Binding of Influenza Hemagglutinin and Neuraminidase to Sialic Acid. Biophys. J. 2019, 116, 1577. [Google Scholar] [CrossRef]
- van Riel, D.; den Bakker, M.A.; Leijten, L.M.E.; Chutinimitkul, S.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Seasonal and Pandemic Human Influenza Viruses Attach Better to Human Upper Respiratory Tract Epithelium than Avian Influenza Viruses. Am. J. Pathol. 2010, 176, 1614–1618. [Google Scholar] [CrossRef] [PubMed]
- van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Human and avian influenza viruses target different cells in the lower respiratory tract of humans and other mammals. Am. J. Pathol. 2007, 171, 1215–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdugo, P.; Tam, P.Y.; Butler, J. Conformational structure of respiratory mucus studied by laser correlation spectroscopy. Biorheology 1983, 20, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Comstock, A.T.; Sajjan, U.S. Barrier function of airway tract epithelium. Tissue Barriers 2013, 1, e24997. [Google Scholar] [CrossRef]
- Moser, M.R.; Bender, T.R.; Margolis, H.S.; Noble, G.R.; Kendal, A.P.; Ritter, D.G. An outbreak of influenza aboard a commercial airliner. Am. J. Epidemiol. 1979, 110, 1–6. [Google Scholar] [CrossRef]
- Nguyen-Van-Tam, J.S.; Killingley, B.; Enstone, J.; Hewitt, M.; Pantelic, J.; Grantham, M.L.; Bueno de Mesquita, P.J.; Lambkin-Williams, R.; Gilbert, A.; Mann, A.; et al. Minimal transmission in an influenza A (H3N2) human challenge-transmission model within a controlled exposure environment. PLoS Pathog. 2020, 16, e1008704. [Google Scholar] [CrossRef]
- Killingley, B.; Nguyen-Van-Tam, J. Routes of influenza transmission. Influenza Other Resp. 2013, 7, 42–51. [Google Scholar] [CrossRef]
- Mitman, M. Aerial Infection. Br. Med. J. 1945, 1, 71–74. [Google Scholar] [CrossRef]
- Wells, W.F. On air-borne infection: Study II. droplets and droplet nuclei. Am. J. Epidemiol. 1934, 20, 611–618. [Google Scholar] [CrossRef]
- Bean, B.; Moore, B.M.; Sterner, B.; Peterson, L.R.; Gerding, D.N.; Balfour, H.H., Jr. Survival of influenza viruses on environmental surfaces. J. Infect. Dis. 1982, 146, 47–51. [Google Scholar] [CrossRef]
- Thomas, Y.; Vogel, G.; Wunderli, W.; Suter, P.; Witschi, M.; Koch, D.; Tapparel, C.; Kaiser, L. Survival of influenza virus on banknotes. Appl. Environ. Microbiol. 2008, 74, 3002–3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julenius, K.; Molgaard, A.; Gupta, R.; Brunak, S. Prediction, conservation analysis, and structural characterization of mammalian mucin-type O-glycosylation sites. Glycobiology 2005, 15, 153–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, S.G.; Burnet, F.M.; Fazekas de St Groth, S.; McCrea, J.F.; Stone, J.D. Mucins and mucoids in relation to influenza virus action; general discussion. Aust. J. Exp. Biol. Med. Sci. 1948, 26 Pt 5, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Vahey, M.D.; Fletcher, D.A. Influenza A virus surface proteins are organized to help penetrate host mucus. Elife 2019, 8, e43764. [Google Scholar] [CrossRef]
- Yoshimura, A.; Kuroda, K.; Kawasaki, K.; Yamashina, S.; Maeda, T.; Ohnishi, S. Infectious cell entry mechanism of influenza virus. J. Virol. 1982, 43, 284–293. [Google Scholar] [CrossRef] [Green Version]
- de Vries, E.; Tscherne, D.M.; Wienholts, M.J.; Cobos-Jimenez, V.; Scholte, F.; Garcia-Sastre, A.; Rottier, P.J.M.; de Haan, C.A.M. Dissection of the Influenza A Virus Endocytic Routes Reveals Macropinocytosis as an Alternative Entry Pathway. PLoS Pathog. 2011, 7, e1001329. [Google Scholar] [CrossRef]
- Akole, A.; Warner, J.M. Model of influenza virus acidification. PLoS ONE 2019, 14, e0214448. [Google Scholar] [CrossRef]
- Pinto, L.H.; Holsinger, L.J.; Lamb, R.A. Influenza-Virus M2 Protein Has Ion Channel Activity. Cell 1992, 69, 517–528. [Google Scholar] [CrossRef]
- Steinhauer, D.A.; Wharton, S.A.; Skehel, J.J.; Wiley, D.C.; Hay, A.J. Amantadine Selection of a Mutant Influenza-Virus Containing an Acid-Stable Hemagglutinin Glycoprotein—Evidence for Virus-Specific Regulation of the Ph of Glycoprotein Transport Vesicles. Proc. Natl. Acad. Sci. USA 1991, 88, 11525–11529. [Google Scholar] [CrossRef] [Green Version]
- Singanayagam, A.; Zambon, M.; Barclay, W.S. Influenza Virus with Increased pH of Hemagglutinin Activation Has Improved Replication in Cell Culture but at the Cost of Infectivity in Human Airway Epithelium. J. Virol. 2019, 93, e00058-19. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Yang, G.H.; DeBeauchamp, J.; Crumpton, J.C.; Kim, H.; Litt, L.; Wan, X.F.; Kercher, L.; Bowman, A.S.; Webster, R.G.; et al. HA stabilization promotes replication and transmission of swine H1N1 gamma influenza viruses in ferrets. eLife 2020, 9, e56236. [Google Scholar] [CrossRef] [PubMed]
- Bause, E.; Legler, G. The role of the hydroxy amino acid in the triplet sequence Asn-Xaa-Thr(Ser) for the N-glycosylation step during glycoprotein biosynthesis. Biochem. J. 1981, 195, 639–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connor, R.J.; Kawaoka, Y.; Webster, R.G.; Paulson, J.C. Receptor Specificity in Human, Avian, and Equine H2 and H3 Influenza-Virus Isolates. Virology 1994, 205, 17–23. [Google Scholar] [CrossRef]
- Brockhausen, I.; Kuhns, W. Role and metabolism of glycoconjugate sulfation. Trends Glycosci. Glyc. 1997, 9, 379–398. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Qi, J.; Khedri, Z.; Diaz, S.; Yu, H.; Chen, X.; Varki, A.; Shi, Y.; Gao, G.F. An Open Receptor-Binding Cavity of Hemagglutinin-Esterase-Fusion Glycoprotein from Newly-Identified Influenza D Virus: Basis for Its Broad Cell Tropism. PLoS Pathog. 2016, 12, e1005411. [Google Scholar]
- Herrler, G.; Szepanski, S.; Schultze, B. 9-O-acetylated sialic acid, a receptor determinant for influenza C virus and coronaviruses. Behring. Inst. Mitt. 1991, 89, 177–184. [Google Scholar]
- Wang, Q.H.; Tian, X.; Chen, X.R.; Ma, J.P. Structural basis for receptor specificity of influenza B virus hemagglutinin. Proc. Natl. Acad. Sci. USA 2007, 104, 16874–16879. [Google Scholar] [CrossRef] [Green Version]
- Varki, N.M.; Varki, A. Diversity in cell surface sialic acid presentations: Implications for biology and disease. Lab. Investig. 2007, 87, 851–857. [Google Scholar] [CrossRef] [Green Version]
- Matrosovich, M.N.; Gambaryan, A.S.; Tuzikov, A.B.; Byramova, N.E.; Mochalova, L.V.; Golbraikh, A.A.; Shenderovich, M.D.; Finne, J.; Bovin, N.V. Probing of the receptor-binding sites of the H1 and H3 influenza A and influenza B virus hemagglutinins by synthetic and natural sialosides. Virology 1993, 196, 111–121. [Google Scholar] [CrossRef]
- Rogers, G.N.; D’Souza, B.L. Receptor binding properties of human and animal H1 influenza virus isolates. Virology 1989, 173, 317–322. [Google Scholar] [CrossRef]
- Broszeit, F.; Tzarum, N.; Zhu, X.Y.; Nemanichvili, N.; Eggink, D.; Leenders, T.; Li, Z.S.; Liu, L.; Wolfert, M.A.; Papanikolaou, A.; et al. N-Glycolylneuraminic Acid as a Receptor for Influenza A Viruses. Cell Rep. 2019, 27, 3284–3294.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambaryan, A.S.; Matrosovich, T.Y.; Philipp, J.; Munster, V.J.; Fouchier, R.A.; Cattoli, G.; Capua, I.; Krauss, S.L.; Webster, R.G.; Banks, J.; et al. Receptor-binding profiles of H7 subtype influenza viruses in different host species. J. Virol. 2012, 86, 4370–4379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higa, H.H.; Rogers, G.N.; Paulson, J.C. Influenza virus hemagglutinins differentiate between receptor determinants bearing N-acetyl-, N-glycollyl-, and N,O-diacetylneuraminic acids. Virology 1985, 144, 279–282. [Google Scholar] [CrossRef]
- Masuda, H.; Suzuki, T.; Sugiyama, Y.; Horiike, G.; Murakami, K.; Miyamoto, D.; Hidari, K.I.P.J.; Ito, T.; Kida, H.; Kiso, M.; et al. Substitution of amino acid residue in influenza A virus hemagglutinin affects recognition of sialyl-oligosaccharides containing N-glycolylneuraminic acid. FEBS Lett. 1999, 464, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Wen, F.; Blackmon, S.; Olivier, A.K.; Li, L.; Guan, M.H.; Sun, H.L.; Wang, P.G.; Wan, X.F. Mutation W222L at the Receptor Binding Site of Hemagglutinin Could Facilitate Viral Adaption from Equine Influenza A(H3N8) Virus to Dogs. J. Virol. 2018, 92, e01115-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.X.; Tscherne, D.M.; McCullough, C.; Caffrey, M.; Garcia-Sastre, A.; Rong, L.J. Residue Y161 of Influenza Virus Hemagglutinin Is Involved in Viral Recognition of Sialylated Complexes from Different Hosts. J. Virol. 2012, 86, 4455–4462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, N.; Martin, B.E.; Yang, C.K.; Luo, F.; Wan, X.F. Association analyses of large-scale glycan microarray data reveal novel host-specific substructures in influenza A virus binding glycans. Sci. Rep. 2015, 5, 15778. [Google Scholar] [CrossRef] [Green Version]
- Byrd-Leotis, L.; Liu, R.P.; Bradley, K.C.; Lasanajak, Y.; Cummings, S.F.; Song, X.Z.; Heimburg-Molinaro, J.; Galloway, S.E.; Culhane, M.R.; Smith, D.F.; et al. Shotgun glycomics of pig lung identifies natural endogenous receptors for influenza viruses. Proc. Natl. Acad. Sci. USA 2014, 111, E2241–E2250. [Google Scholar] [CrossRef] [Green Version]
- Broszeit, F.; van Beek, R.J.; Unione, L.; Bestebroer, T.M.; Chapla, D.; Yang, J.Y.; Moremen, K.W.; Herfst, S.; Fouchier, R.A.M.; de Vries, R.P.; et al. Glycan remodeled erythrocytes facilitate antigenic characterization of recent A/H3N2 influenza viruses. Nat. Commun. 2021, 12, 5449. [Google Scholar] [CrossRef]
- Peng, W.J.; de Vries, R.P.; Grant, O.C.; Thompson, A.J.; McBride, R.; Tsogtbaatar, B.; Lee, P.S.; Razi, N.; Wilson, I.A.; Woods, R.J.; et al. Recent H3N2 Viruses Have Evolved Specificity for Extended, Branched Human-type Receptors, Conferring Potential for Increased Avidity. Cell Host Microbe 2017, 21, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Gulati, S.; Smith, D.F.; Cummings, R.D.; Couch, R.B.; Griesemer, S.B.; George, K.S.; Webster, R.G.; Air, G.M. Human H3N2 Influenza Viruses Isolated from 1968 To 2012 Show Varying Preference for Receptor Substructures with No Apparent Consequences for Disease or Spread. PLoS ONE 2013, 8, e66325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrd-Leotis, L.; Jia, N.; Dutta, S.; Trost, J.F.; Gao, C.; Cummings, S.F.; Braulke, T.; Muller-Loennies, S.; Heimburg-Molinaro, J.; Steinhauer, D.A.; et al. Influenza binds phosphorylated glycans from human lung. Sci Adv. 2019, 5, eaav2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrd-Leotis, L.; Gao, C.; Jia, N.; Mehta, A.Y.; Trost, J.; Cummings, S.F.; Heimburg-Molinaro, J.; Cummings, R.D.; Steinhauer, D.A. Antigenic Pressure on H3N2 Influenza Virus Drift Strains Imposes Constraints on Binding to Sialylated Receptors but Not Phosphorylated Glycans. J. Virol. 2019, 93, e01178-19. [Google Scholar] [CrossRef] [Green Version]
- Stray, S.J.; Cummings, R.D.; Air, G.M. Influenza virus infection of desialylated cells. Glycobiology 2000, 10, 649–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nycholat, C.M.; McBride, R.; Ekiert, D.C.; Xu, R.; Rangarajan, J.; Peng, W.; Razi, N.; Gilbert, M.; Wakarchuk, W.; Wilson, I.A.; et al. Recognition of sialylated poly-N-acetyllactosamine chains on N- and O-linked glycans by human and avian influenza A virus hemagglutinins. Angew. Chem. Int. Ed. Engl. 2012, 51, 4860–4863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, J.; Lau, K.; Lai, J.C.C.; Gagarinov, I.A.; Shi, Y.; McAtamney, S.; Chan, R.W.Y.; Nicholls, J.; von Itzstein, M.; Haselhorst, T. Unravelling the Role of O-glycans in Influenza A Virus Infection. Sci. Rep. 2018, 8, 16382. [Google Scholar] [CrossRef] [PubMed]
- Chu, V.C.; Whittaker, G.R. Influenza virus entry and infection require host cell N-linked glycoprotein. Proc. Natl. Acad. Sci. USA 2004, 101, 18153–18158. [Google Scholar] [CrossRef] [Green Version]
- Oshansky, C.M.; Pickens, J.A.; Bradley, K.C.; Jones, L.P.; Saavedra-Ebner, G.M.; Barber, J.P.; Crabtree, J.M.; Steinhauer, D.A.; Tompkins, S.M.; Tripp, R.A. Avian influenza viruses infect primary human bronchial epithelial cells unconstrained by sialic acid alpha2,3 residues. PLoS ONE 2011, 6, e21183. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, J.M.; Bourne, A.J.; Chen, H.; Guan, Y.; Peiris, J.S. Sialic acid receptor detection in the human respiratory tract: Evidence for widespread distribution of potential binding sites for human and avian influenza viruses. Respir. Res. 2007, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, Y.; Nishide, S.; Ose, T.; Suzuki, T.; Kato, I.; Fukuhara, H.; Fujioka, M.; Horiuchi, K.; Satoh, A.O.; Nepal, P.; et al. A Sialylated Voltage-Dependent Ca(2+) Channel Binds Hemagglutinin and Mediates Influenza A Virus Entry into Mammalian Cells. Cell Host Microbe 2018, 23, 809–818.e5. [Google Scholar] [CrossRef] [Green Version]
- Flynn, R.A.; Pedram, K.; Malaker, S.A.; Batista, P.J.; Smith, B.A.H.; Johnson, A.G.; George, B.M.; Majzoub, K.; Villalta, P.W.; Carette, J.E.; et al. Small RNAs are modified with N-glycans and displayed on the surface of living cells. Cell 2021, 184, 3109–3124.e22. [Google Scholar] [CrossRef] [PubMed]
- El-Hashash, A.H.; Warburton, D. Cell Polarity and Spindle Orientation in the Distal Epithelium of Embryonic Lung. Dev. Dynam. 2011, 240, 441–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berube, K.; Prytherch, Z.; Job, C.; Hughes, T. Human primary bronchial lung cell constructs: The new respiratory models. Toxicology 2010, 278, 311–318. [Google Scholar] [CrossRef]
- Chan, M.C.; Chan, R.W.; Yu, W.C.; Ho, C.C.; Chui, W.H.; Lo, C.K.; Yuen, K.M.; Guan, Y.I.; Nicholls, J.M.; Peiris, J.S. Influenza H5N1 virus infection of polarized human alveolar epithelial cells and lung microvascular endothelial cells. Respir. Res. 2009, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Banks, L. Upsetting the Balance: When Viruses Manipulate Cell Polarity Control. J. Mol. Biol. 2018, 430, 3481–3503. [Google Scholar] [CrossRef]
- Wang, Y.H.; Lu, X. Cell Polarity: A Key Defence Mechanism Against Infection and Cancer Cell Invasion? Cell Polarity 2015, 2, 167–186. [Google Scholar]
- Momose, F.; Kikuchi, Y.; Komase, K.; Morikawa, Y. Visualization of microtubule-mediated transport of influenza viral progeny ribonucleoprotein. Microbes Infect. 2007, 9, 1422–1433. [Google Scholar] [CrossRef]
- Naghavi, M.H.; Walsh, D. Microtubule Regulation and Function during Virus Infection. J. Virol. 2017, 91, e00538-17. [Google Scholar] [CrossRef] [Green Version]
- Amorim, M.J.; Bruce, E.A.; Read, E.K.; Foeglein, A.; Mahen, R.; Stuart, A.D.; Digard, P. A Rab11- and microtubule-dependent mechanism for cytoplasmic transport of influenza A virus viral RNA. J. Virol. 2011, 85, 4143–4156. [Google Scholar] [CrossRef] [Green Version]
- Wohlgemuth, N.; Lane, A.P.; Pekosz, A. Influenza A Virus M2 Protein Apical Targeting Is Required for Efficient Virus Replication. J. Virol. 2018, 92, e01425-18. [Google Scholar] [CrossRef] [Green Version]
- Peteranderl, C.; Kuznetsova, I.; Schulze, J.; Hardt, M.; Lecuona, E.; Sznajder, J.I.; Vadasz, I.; Morty, R.E.; Pleschka, S.; Wolff, T.; et al. Influenza A Virus Infection Induces Apical Redistribution of Na+, K+-ATPase in Lung Epithelial Cells In Vitro and In Vivo. Am. J. Resp. Cell Mol. 2019, 61, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, H.; Fang, M.; Chen, X.; Zeng, X. A human cell polarity protein Lgl2 regulates influenza A virus nucleoprotein exportation from nucleus in MDCK cells. J. Biosci. 2020, 45, 67. [Google Scholar] [CrossRef] [PubMed]
- Golebiewski, L.; Liu, H.B.; Javier, R.T.; Rice, A.P. The Avian Influenza Virus NS1 ESEV PDZ Binding Motif Associates with Dlg1 and Scribble To Disrupt Cellular Tight Junctions. J. Virol. 2011, 85, 10639–10648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.B.; Golebiewski, L.; Dow, E.C.; Krug, R.M.; Javier, R.T.; Rice, A.P. The ESEV PDZ-Binding Motif of the Avian Influenza A Virus NS1 Protein Protects Infected Cells from Apoptosis by Directly Targeting Scribble. J. Virol. 2010, 84, 11164–11174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Liu, H.B.; Rice, A.P. Regulation of Interferon-beta by MAGI-1 and Its Interaction with Influenza A Virus NS1 Protein with ESEV PBM. PLoS ONE 2012, 7, e41251. [Google Scholar]
- McCartney, M.M.; Linderholm, A.L.; Yamaguchi, M.S.; Falcon, A.K.; Harper, R.W.; Thompson, G.R.; Ebeler, S.E.; Kenyon, N.J.; Davis, C.E.; Schivo, M. Predicting influenza and rhinovirus infections in airway cells utilizing volatile emissions. J. Infect. Dis. 2021, 224, 1742–1750. [Google Scholar] [CrossRef]
- Chen, A.J.; Dong, J.; Yuan, X.H.; Bo, H.; Li, S.Z.; Wang, C.; Duan, Z.J.; Zheng, L.S. Anti-H7N9 avian influenza A virus activity of interferon in pseudostratified human airway epithelium cell cultures. Virol. J. 2019, 16, 44. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Liu, G.; Jia, Y.; Yang, J.; Shi, J.; Dong, J.; Wei, J.; Liu, X. Characterization of air-liquid interface culture of A549 alveolar epithelial cells. Braz J. Med. Biol. Res. 2017, 51, e6950. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Bukreyev, A.; Thompson, C.I.; Watson, B.; Peeples, M.E.; Collins, P.L.; Pickles, R.J. Infection of ciliated cells by human parainfluenza virus type 3 in an in vitro model of human airway epithelium. J. Virol. 2005, 79, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.I.; Barclay, W.S.; Zambon, M.C.; Pickles, R.J. Infection of human airway epithelium by human and avian strains of influenza a virus. J. Virol. 2006, 80, 8060–8068. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.Y.; Liu, J.; Yang, Y.; Wu, M.; Ye, L.N.; Chen, S.; Zhang, T.; Zeng, Z.H.; Zhang, K.; Cai, K.H.; et al. Coupled CRC 2D and ALI 3D Cultures Express Receptors of Emerging Viruses and Are More Suitable for the Study of Viral Infections Compared to Conventional Cell Lines. Stem Cells Int. 2020, 2020, 2421689. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.H.; Yang, W.; Beineke, A.; Dijkman, R.; Matrosovich, M.; Baumgartner, W.; Thiel, V.; Valentin-Weigand, P.; Meng, F.D.; Herrler, G. The differentiated airway epithelium infected by influenza viruses maintains the barrier function despite a dramatic loss of ciliated cells. Sci. Rep. 2016, 6, 39668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Poucke, S.G.; Nicholls, J.M.; Nauwynck, H.J.; Van Reeth, K. Replication of avian, human and swine influenza viruses in porcine respiratory explants and association with sialic acid distribution. Virol. J. 2010, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Goldsmith, C.S.; Maines, T.R.; Belser, J.A.; Gustin, K.M.; Pekosz, A.; Zaki, S.R.; Katz, J.M.; Tumpey, T.M. Tropism and infectivity of influenza virus, including highly pathogenic avian H5N1 virus, in ferret tracheal differentiated primary epithelial cell cultures. J. Virol. 2013, 87, 2597–2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, S.W.; Lorbach, J.N.; Nolting, J.M.; Stull, J.W.; Jackwood, D.J.; Davis, I.C.; Bowman, A.S. Madin-Darby canine kidney cell sialic acid receptor modulation induced by culture medium conditions: Implications for the isolation of influenza A virus. Influenza Other Respir. Viruses 2019, 13, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Matrosovich, M.; Matrosovich, T.; Carr, J.; Roberts, N.A.; Klenk, H.D. Overexpression of the alpha-2,6-sialyltransferase in MDCK cells increases influenza virus sensitivity to neuraminidase inhibitors. J. Virol. 2003, 77, 8418–8425. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.C.; Kappes, M.A.; Chen, M.C.; Lin, C.C.; Wang, T.T. Distinct susceptibility and applicability of MDCK derivatives for influenza virus research. PLoS ONE 2017, 12, e0172299. [Google Scholar] [CrossRef] [Green Version]
- Takada, K.; Kawakami, C.; Fan, S.; Chiba, S.; Zhong, G.; Gu, C.; Shimizu, K.; Takasaki, S.; Sakai-Tagawa, Y.; Lopes, T.J.S.; et al. A humanized MDCK cell line for the efficient isolation and propagation of human influenza viruses. Nat. Microbiol. 2019, 4, 1268–1273. [Google Scholar] [CrossRef]
- Huipao, N.; Borwornpinyo, S.; Wiboon-Ut, S.; Campbell, C.R.; Lee, I.H.; Hiranyachattada, S.; Sukasem, C.; Thitithanyanont, A.; Pholpramool, C.; Cook, D.I.; et al. P2Y6 receptors are involved in mediating the effect of inactivated avian influenza virus H5N1 on IL-6 & CXCL8 mRNA expression in respiratory epithelium. PLoS ONE 2017, 12, e0176974. [Google Scholar]
- Pamies, D.; Bal-Price, A.; Chesne, C.; Coecke, S.; Dinnyes, A.; Eskes, C.; Grillari, R.; Gstraunthaler, G.; Hartung, T.; Jennings, P.; et al. Advanced Good Cell Culture Practice for Human Primary, Stem Cell-Derived and Organoid Models as well as Microphysiological Systems. Altex-Altern. Anim. Exp. 2018, 35, 353–378. [Google Scholar] [CrossRef]
- Schamberger, A.C.; Staab-Weijnitz, C.A.; Mise-Racek, N.; Eickelberg, O. Cigarette smoke alters primary human bronchial epithelial cell differentiation at the air-liquid interface. Sci. Rep. 2015, 5, 8163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Dufour, J.M. Cell lines: Valuable tools or useless artifacts. Spermatogenesis 2012, 2, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.P.; Kumar, C.; Bohl, S.; Klingmueller, U.; Mann, M. Comparative Proteomic Phenotyping of Cell Lines and Primary Cells to Assess Preservation of Cell Type-specific Functions. Mol. Cell Proteom. 2009, 8, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.W.; Plancarte, M.; Ball, E.E.; Weiss, C.M.; Gonzales-Viera, O.; Holcomb, K.; Ma, Z.M.; Allen, A.M.; Reader, J.R.; Duignan, P.J.; et al. Respiratory Tract Explant Infection Dynamics of Influenza A Virus in California Sea Lions, Northern Elephant Seals, and Rhesus Macaques. J. Virol. 2021, 95, e0040321. [Google Scholar] [CrossRef]
- Chan, L.L.Y.; Bui, C.T.H.; Mok, C.K.P.; Ng, M.M.T.; Nicholls, J.M.; Peiris, J.S.M.; Chan, M.C.W.; Chan, R.W.Y. Evaluation of the human adaptation of influenza A/H7N9 virus in PB2 protein using human and swine respiratory tract explant cultures. Sci. Rep. 2016, 6, 3540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzetto, E.; Bortolami, A.; Fusaro, A.; Mazzacan, E.; Maniero, S.; Vascellari, M.; Beato, M.S.; Schiavon, E.; Chiapponi, C.; Terregino, C.; et al. Replication of Influenza D Viruses of Bovine and Swine Origin in Ovine Respiratory Explants and Their Attachment to the Respiratory Tract of Bovine, Sheep, Goat, Horse, and Swine. Front. Microbiol. 2020, 11, 1136. [Google Scholar] [CrossRef]
- Jones, R. The glycoproteins of secretory cells in airway epithelium. Ciba Found. Symp. 1978, 1, 175–193. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Praena, B.; Wan, X.-F. Influenza Virus Infections in Polarized Cells. Viruses 2022, 14, 1307. https://doi.org/10.3390/v14061307
Praena B, Wan X-F. Influenza Virus Infections in Polarized Cells. Viruses. 2022; 14(6):1307. https://doi.org/10.3390/v14061307
Chicago/Turabian StylePraena, Beatriz, and Xiu-Feng Wan. 2022. "Influenza Virus Infections in Polarized Cells" Viruses 14, no. 6: 1307. https://doi.org/10.3390/v14061307
APA StylePraena, B., & Wan, X. -F. (2022). Influenza Virus Infections in Polarized Cells. Viruses, 14(6), 1307. https://doi.org/10.3390/v14061307