
An Equine Model for Vaccination against a Hepacivirus: Insights into Host Responses to E2 Recombinant Protein Vaccination and Subsequent Equine Hepacivirus Inoculation


 , , , , , , , ,
, , , , , , , ,  , add
Show full author list
, add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. EqHV E2 Recombinant Protein Production
2.3. Vaccination and EqHV Inoculation
2.4. Sample Collection
2.5. Laboratory Analysis
2.5.1. Detection of EqHV RNA in Serum
2.5.2. Detection and Isotyping of Anti-EqHV E2-specific IgG (E2 IgG) in Serum
2.5.3. Detection of Anti-EqHV NS3-Specific IgG in Serum
2.5.4. Monitoring of Haematology, Serum Biochemistry, and Clinical Parameters
2.5.5. 1H Nuclear Magnetic Resonance (NMR) Spectroscopy of Serum Samples
2.5.6. RNA Extraction and RNA Sequencing (RNA-seq) of Liver Tissue
2.5.7. Histopathology, Immunohistochemistry (IHC), and Fluorescent in Situ Hybridization (FISH) of Liver Tissue
2.5.8. Detection of EqHV RNA in Liver Tissue
3. Results
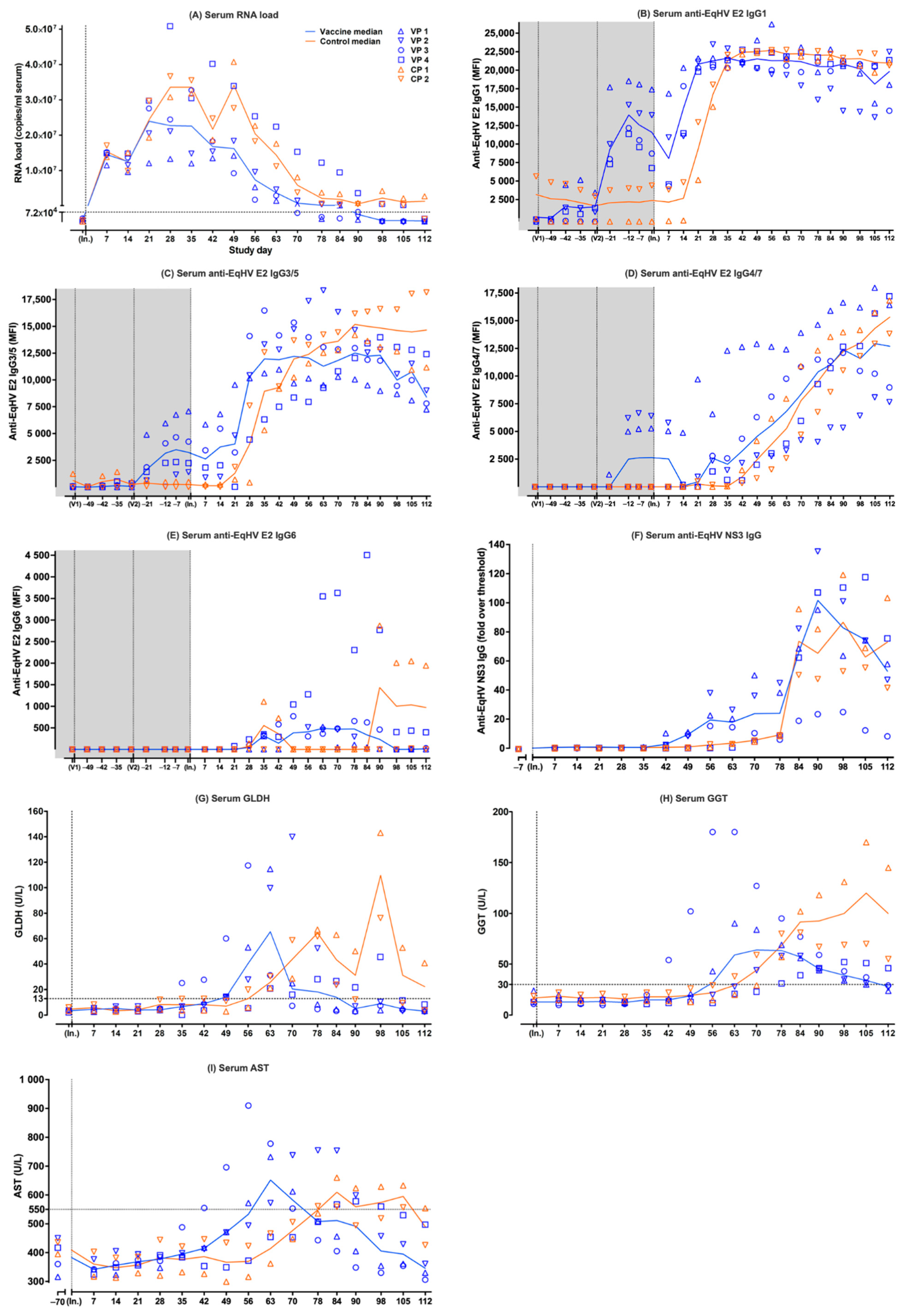
3.1. EqHV RNA in Serum of Vaccine Ponies and Control Ponies
3.2. Anti-EqHV E2-Specific IgG Isotypes in Serum of Vaccine Ponies and Control Ponies
3.3. Anti-EqHV NS3-Specific IgG in Serum of Vaccine Ponies and Control Ponies
3.4. Liver-Associated Serum Biochemistry Parameters
3.5. Clinical Scoring
3.6. Serum Metabolomics
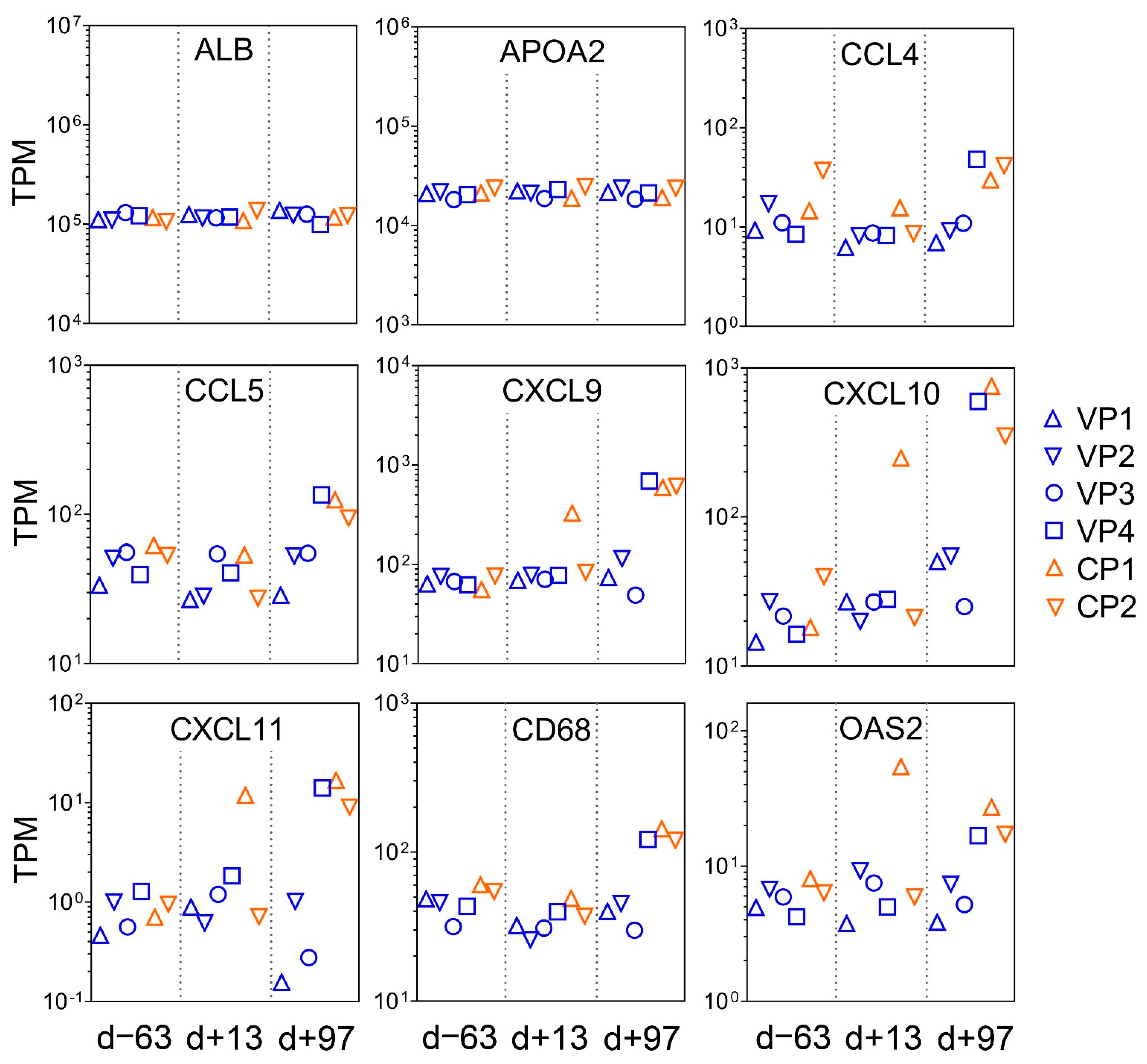
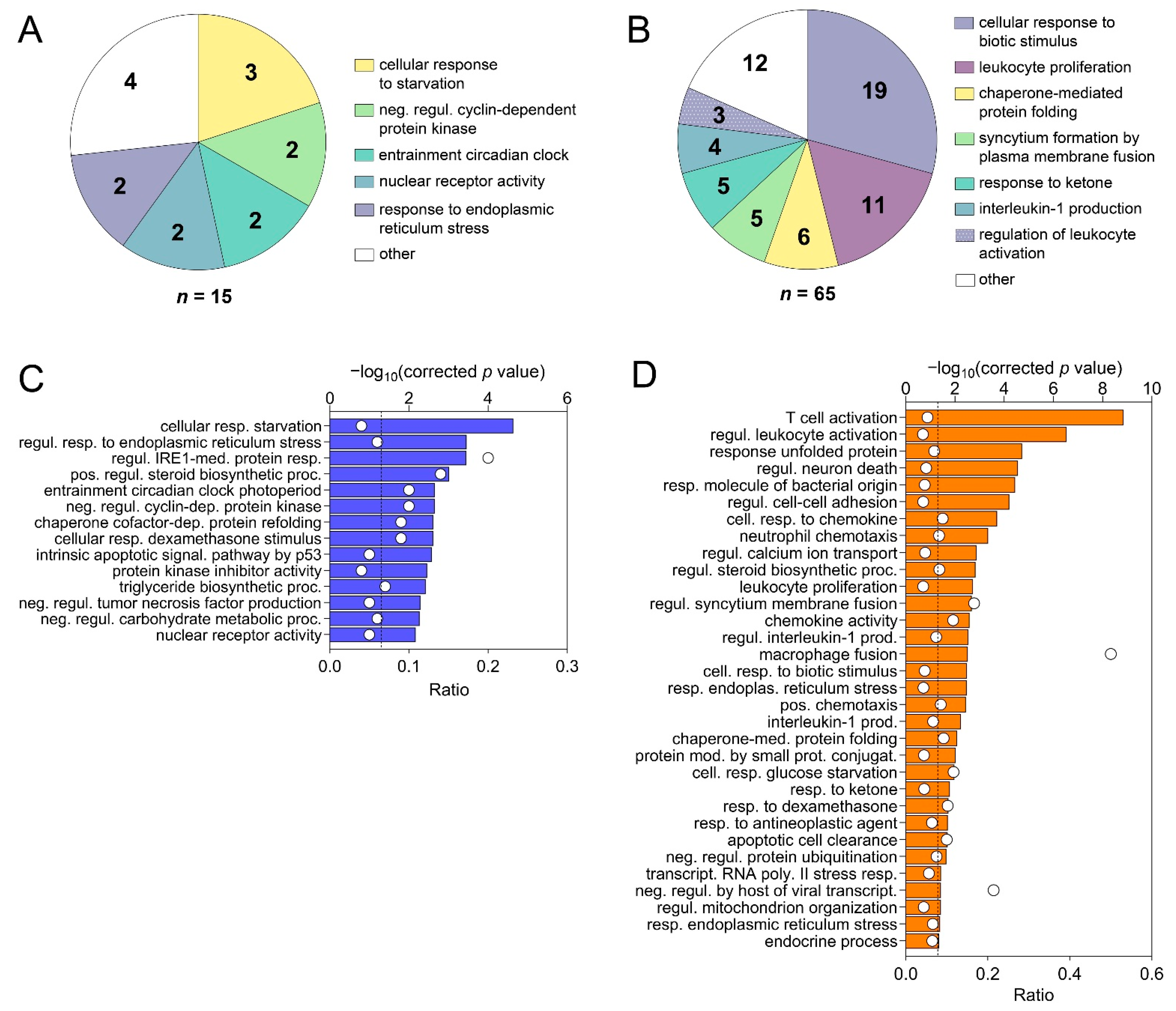
3.7. RNA-seq of Liver Tissue
3.8. Histopathology, IHC, FISH, and qPCR of Liver Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, D.B.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, A.S.; Pletnev, A.; Rico-Hesse, R.; Stapleton, J.T.; et al. Proposed update to the taxonomy of the genera Hepacivirus and Pegivirus within the Flaviviridae family. J. Gen. Virol. 2016, 97, 2894–2907. [Google Scholar] [CrossRef] [PubMed]
- Hartlage, A.S.; Cullen, J.M.; Kapoor, A. The Strange, Expanding World of Animal Hepaciviruses. Annu. Rev. Virol. 2016, 3, 53–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, A.; Simmonds, P.; Gerold, G.; Qaisar, N.; Jain, K.; Henriquez, J.A.; Firth, C.; Hirschberg, D.L.; Rice, C.M.; Shields, S.; et al. Characterization of a canine homolog of hepatitis C virus. Proc. Natl. Acad. Sci. USA 2011, 108, 11608–11613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsay, J.D.; Evanoff, R.; Wilkinson, T.E., Jr.; Divers, T.J.; Knowles, D.P.; Mealey, R.H. Experimental transmission of equine hepacivirus in horses as a model for hepatitis C virus. Hepatology 2015, 61, 1533–1546. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kasai, H.; Yamashita, A.; Okuyama-Dobashi, K.; Yasumoto, J.; Maekawa, S.; Enomoto, N.; Okamoto, T.; Matsuura, Y.; Morimatsu, M.; et al. Hallmarks of hepatitis C virus in equine hepacivirus. J. Virol. 2014, 88, 13352–13366. [Google Scholar] [CrossRef] [Green Version]
- Pfaender, S.; Walter, S.; Grabski, E.; Todt, D.; Bruening, J.; Romero-Brey, I.; Gather, T.; Brown, R.J.; Hahn, K.; Puff, C.; et al. Immune protection against reinfection with nonprimate hepacivirus. Proc. Natl. Acad. Sci. USA 2017, 114, E2430–E2439. [Google Scholar] [CrossRef] [Green Version]
- Pfaender, S.; Cavalleri, J.M.; Walter, S.; Doerrbecker, J.; Campana, B.; Brown, R.J.; Burbelo, P.D.; Postel, A.; Hahn, K.; Anggakusuma; et al. Clinical course of infection and viral tissue tropism of hepatitis C virus-like nonprimate hepaciviruses in horses. Hepatology 2015, 61, 447–459. [Google Scholar] [CrossRef]
- Scheel, T.K.; Kapoor, A.; Nishiuchi, E.; Brock, K.V.; Yu, Y.; Andrus, L.; Gu, M.; Renshaw, R.W.; Dubovi, E.J.; McDonough, S.P.; et al. Characterization of nonprimate hepacivirus and construction of a functional molecular clone. Proc. Natl. Acad. Sci. USA 2015, 112, 2192–2197. [Google Scholar] [CrossRef] [Green Version]
- Bailey, J.R.; Barnes, E.; Cox, A.L. Approaches, Progress, and Challenges to Hepatitis C Vaccine Development. Gastroenterology 2019, 156, 418–430. [Google Scholar] [CrossRef] [Green Version]
- Horohov, D.W. The equine immune responses to infectious and allergic disease: A model for humans? Mol. Immunol. 2015, 66, 89–96. [Google Scholar] [CrossRef]
- Karagianni, A.E.; Lisowski, Z.M.; Hume, D.A.; Scott Pirie, R. The equine mononuclear phagocyte system: The relevance of the horse as a model for understanding human innate immunity. Equine Vet. J. 2020, 53, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Wagner, B.; Miller, D.C.; Lear, T.L.; Antczak, D.F. The complete map of the Ig heavy chain constant gene region reveals evidence for seven IgG isotypes and for IgD in the horse. J. Immunol. 2004, 173, 3230–3242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, M.J.; Wagner, B.; Woof, J.M. The different effector function capabilities of the seven equine IgG subclasses have implications for vaccine strategies. Mol. Immunol. 2008, 45, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Keggan, A.; Freer, H.; Rollins, A.; Wagner, B. Production of seven monoclonal equine immunoglobulins isotyped by multiplex analysis. Vet. Immunol. Immunopathol. 2013, 153, 187–193. [Google Scholar] [CrossRef]
- Wagner, B. Immunoglobulins and immunoglobulin genes of the horse. Dev. Comp. Immunol. 2006, 30, 155–164. [Google Scholar] [CrossRef]
- Walker, M.R.; Eltahla, A.A.; Mina, M.M.; Li, H.; Lloyd, A.R.; Bull, R.A. Envelope-Specific IgG3 and IgG1 Responses Are Associated with Clearance of Acute Hepatitis C Virus Infection. Viruses 2020, 12, 75. [Google Scholar] [CrossRef] [Green Version]
- Stamataki, Z.; Coates, S.; Evans, M.J.; Wininger, M.; Crawford, K.; Dong, C.; Fong, Y.L.; Chien, D.; Abrignani, S.; Balfe, P.; et al. Hepatitis C virus envelope glycoprotein immunization of rodents elicits cross-reactive neutralizing antibodies. Vaccine 2007, 25, 7773–7784. [Google Scholar] [CrossRef]
- Lee, S.; Nguyen, M.T. Recent advances of vaccine adjuvants for infectious diseases. Immune Netw. 2015, 15, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Iwaki, T.; Figuera Losada, M.; Ploplis, V.; Castellino, F. Rapid selection of Drosophila S2 cells with the puromycin resistance gene. BioTechniques 2003, 35, 482–484, 486. [Google Scholar] [CrossRef] [Green Version]
- Burbelo, P.D.; Dubovi, E.J.; Simmonds, P.; Medina, J.L.; Henriquez, J.A.; Mishra, N.; Wagner, J.; Tokarz, R.; Cullen, J.M.; Iadarola, M.J.; et al. Serology-Enabled Discovery of Genetically Diverse Hepaciviruses in a New Host. J. Virol. 2012, 86, 6171–6178. [Google Scholar] [CrossRef] [Green Version]
- Wagner, B.; Freer, H.; Rollins, A.; Erb, H.N. A fluorescent bead-based multiplex assay for the simultaneous detection of antibodies to B. burgdorferi outer surface proteins in canine serum. Vet. Immunol. Immunopathol. 2011, 140, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, A.S.; Lunn, D.P.; Holmes, M.A. Monoclonal antibodies to subclass-specific antigenic determinants on equine immunoglobulin gamma chains and their characterization. Vet. Immunol. Immunopathol. 1998, 62, 153–165. [Google Scholar] [CrossRef]
- Burbelo, P.D.; Ching, K.H.; Klimavicz, C.M.; Iadarola, M.J. Antibody profiling by Luciferase Immunoprecipitation Systems (LIPS). J. Vis. Exp. 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling The False Discovery Rate - A Practical And Powerful Approach To Multiple Testing. J. R. Statist. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Wagner, G.P.; Kin, K.; Lynch, V.J. Measurement of mRNA abundance using RNA-seq data: RPKM measure is inconsistent among samples. Theory Biosci. 2012, 131, 281–285. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Shaw, A.E.; Hughes, J.; Gu, Q.; Behdenna, A.; Singer, J.B.; Dennis, T.; Orton, R.J.; Varela, M.; Gifford, R.J.; Wilson, S.J.; et al. Fundamental properties of the mammalian innate immune system revealed by multispecies comparison of type I interferon responses. PLoS Biol. 2017, 15, e2004086. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postel, A.; Hansmann, F.; Baechlein, C.; Fischer, N.; Alawi, M.; Grundhoff, A.; Derking, S.; Tenhundfeld, J.; Pfankuche, V.M.; Herder, V.; et al. Presence of atypical porcine pestivirus (APPV) genomes in newborn piglets correlates with congenital tremor. Sci. Rep. 2016, 6, 27735. [Google Scholar] [CrossRef] [Green Version]
- Altan, E.; Li, Y.; Sabino-Santos, G., Jr.; Sawaswong, V.; Barnum, S.; Pusterla, N.; Deng, X.; Delwart, E. Viruses in Horses with Neurologic and Respiratory Diseases. Viruses 2019, 11, 942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badenhorst, M.; de Heus, P.; Auer, A.; Rumenapf, T.; Tegtmeyer, B.; Kolodziejek, J.; Nowotny, N.; Steinmann, E.; Cavalleri, J.V. No Evidence of Mosquito Involvement in the Transmission of Equine Hepacivirus (Flaviviridae) in an Epidemiological Survey of Austrian Horses. Viruses 2019, 11, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pronost, S.; Fortier, C.; Marcillaud-Pitel, C.; Tapprest, J.; Foursin, M.; Saunier, B.; Pitel, P.H.; Paillot, R.; Hue, E.S. Further Evidence for in Utero Transmission of Equine Hepacivirus to Foals. Viruses 2019, 11, 1124. [Google Scholar] [CrossRef] [Green Version]
- Goehring, L.S.; Wagner, B.; Bigbie, R.; Hussey, S.B.; Rao, S.; Morley, P.S.; Lunn, D.P. Control of EHV-1 viremia and nasal shedding by commercial vaccines. Vaccine 2010, 28, 5203–5211. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Wolfisberg, R.; Fahnoe, U.; Patel, R.S.; Trivedi, S.; Kumar, A.; Sharma, H.; Nielsen, L.; McDonough, S.P.; Bukh, J.; et al. Pathogenesis, MicroRNA-122 Gene-Regulation, and Protective Immune Responses After Acute Equine Hepacivirus Infection. Hepatology 2021, 74, 1148–1163. [Google Scholar] [CrossRef]
- Gather, T.; Walter, S.; Pfaender, S.; Todt, D.; Feige, K.; Steinmann, E.; Cavalleri, J.M. Acute and chronic infections with nonprimate hepacivirus in young horses. Vet. Res. 2016, 47, 97. [Google Scholar] [CrossRef] [Green Version]
- Nie, A.; Sun, B.; Fu, Z.; Yu, D. Roles of aminoacyl-tRNA synthetases in immune regulation and immune diseases. Cell Death Dis. 2019, 10, 901. [Google Scholar] [CrossRef] [Green Version]
- Metz, P.; Reuter, A.; Bender, S.; Bartenschlager, R. Interferon-stimulated genes and their role in controlling hepatitis C virus. J. Hepatol. 2013, 59, 1331–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomer, A.; Brown, R.J.P.; Pfaender, S.; Deterding, K.; Reuter, G.; Orton, R.; Seitz, S.; Bock, C.T.; Cavalleri, J.M.V.; Pietschmann, T.; et al. Intra-host analysis of hepaciviral glycoprotein evolution reveals signatures associated with viral persistence and clearance. Virus Evol. 2022, 8, veac007. [Google Scholar] [CrossRef] [PubMed]
- Divers, T.J.; Tennant, B.C.; Kumar, A.; McDonough, S.; Cullen, J.; Bhuva, N.; Jain, K.; Chauhan, L.S.; Scheel, T.K.H.; Lipkin, W.I.; et al. New Parvovirus Associated with Serum Hepatitis in Horses after Inoculation of Common Biological Product. Emerg. Infect. Dis. 2018, 24, 303–310. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Parameters Score Overview | ||||||
|---|---|---|---|---|---|---|
| Vaccine Pony 1 | Vaccine Pony 2 | Vaccine Pony 3 | Vaccine Pony 4 | Control Pony 1 | Control Pony 2 | |
| D+7 | 0 | 0 | 0 | 1 | 1 | 0 |
| D+14 | 1 | 0 | 0 | 1 | 2 | 4 |
| D+21 | 0 | 0 | 0 | 1 | 0 | 0 |
| D+28 | 0 | 4 | 0 | 0 | 1 | 0 |
| D+35 | 0 | 0 | 1 | 0 | 1 | 0 |
| D+42 | 0 | 0 | 3 | 1 | 1 | 0 |
| D+49 | 1 | 2 | 5 | 1 | 3 | 0 |
| D+56 | 4 | 1 | 5 | 0 | 0 | 1 |
| D+63 | 5 | 5 | 4 | 1 | 1 | 3 |
| D+70 | 3 | 4 | 3 | 1 | 2 | 4 |
| D+78 | 1 | 4 | 2 | 2 | 3 | 5 |
| D+84 | 1 | 3 | 1 | 3 | 5 | 5 |
| D+90 | 1 | 2 | 2 | 4 | 6 | 3 |
| D+98 | 2 | 1 | 1 | 4 | 5 | 3 |
| D+105 | 1 | 1 | 1 | 2 | 5 | 3 |
| D+112 | 0 | 0 | 1 | 1 | 5 | 2 |
| Total blood parameters score | 20 | 27 | 29 | 23 | 41 | 33 |
| + Total clinical parameters score | 0 | 0 | 2 | 0 | 0 | 0 |
| Total overall clinical score | 20 | 27 | 31 | 23 | 41 | 33 |
| Metabolite | Mean Difference | t | p Value | Adj. p Value |
|---|---|---|---|---|
| Pyruvate | 0.027 | 5.334 | 0.000 | 0.001 |
| Acetoacetate | −0.007 | −5.258 | 0.000 | 0.001 |
| Formate | −0.013 | −5.177 | 0.000 | 0.001 |
| Methanol | −0.058 | −5.153 | 0.000 | 0.001 |
| Albumin | −0.168 | −3.647 | 0.001 | 0.017 |
| Tot.CCE | −12.367 | −3.532 | 0.002 | 0.018 |
| Total protein | −0.064 | −3.458 | 0.002 | 0.018 |
| Carbon chains | −2.594 | −3.443 | 0.002 | 0.018 |
| Sphingomyelins | −0.175 | −3.283 | 0.003 | 0.023 |
| Sub8.Tot.Chol. | −0.024 | −3.087 | 0.005 | 0.031 |
| Fatty acids | −0.514 | −3.072 | 0.006 | 0.031 |
| Sub7.Tot.Chol. | −0.023 | −3.039 | 0.006 | 0.031 |
| Triglycerids | −0.203 | −2.946 | 0.008 | 0.032 |
| Multi unsat.CCE | −0.652 | −2.899 | 0.008 | 0.032 |
| Histidine | −0.013 | −2.871 | 0.009 | 0.032 |
| Alpha unsat.CCE | −1.477 | −2.842 | 0.010 | 0.032 |
| Glutamine | −0.091 | −2.829 | 0.010 | 0.032 |
| Unsat.CCE | −2.496 | −2.820 | 0.010 | 0.032 |
| Sub9.Tot.Chol. | −0.018 | −2.817 | 0.010 | 0.032 |
| Tot.cholesterol | −0.127 | −2.737 | 0.012 | 0.035 |
| Creatinine | −0.037 | −2.719 | 0.013 | 0.035 |
| Mannose | 0.014 | 2.711 | 0.013 | 0.035 |
| Lactate | 0.425 | 2.579 | 0.017 | 0.045 |
| Sub6.Tot.Chol. | −0.015 | −2.537 | 0.019 | 0.047 |
| Mean Liver Viral Load (Viral Copies/50 ng Total RNA; LLOQ = 1 × 103 Copies/50 ng Total RNA) | Serum Viral Load (Viral Copies/mL Serum; LLOQ = 7.2 × 104 Copies/mL Serum) | |
|---|---|---|
| Day +13 | Day +14 | |
| Vaccine Pony 1 | 8.72 × 104 | 9.51 × 106 |
| Vaccine Pony 2 | 3.55 × 104 | 1.14 × 107 |
| Vaccine Pony 3 | 7.52 × 104 | 1.36 × 107 |
| Vaccine Pony 4 | 1.30 × 105 | 1.47 × 107 |
| Control Pony 1 | 8.77 × 104 | 1.50 × 107 |
| Control Pony 2 | 7.66 × 104 | 9.80 × 106 |
| Day +97 | Day +98 | |
| Vaccine Pony 1 | 1.27 × 103 | <LLOQ |
| Vaccine Pony 2 | <LLOQ | <LLOQ |
| Vaccine Pony 3 | <LLOQ c | <LLOQ |
| Vaccine Pony 4 | 1.00 × 104 | 4.66 × 105 |
| Control Pony 1 | 9.28 × 103 | 4.13 × 106 |
| Control Pony 2 | <LLOQ | 3.01 × 105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badenhorst, M.; Saalmüller, A.; Daly, J.M.; Ertl, R.; Stadler, M.; Puff, C.; de le Roi, M.; Baumgärtner, W.; Engelmann, M.; Brandner, S.; et al. An Equine Model for Vaccination against a Hepacivirus: Insights into Host Responses to E2 Recombinant Protein Vaccination and Subsequent Equine Hepacivirus Inoculation. Viruses 2022, 14, 1401. https://doi.org/10.3390/v14071401
Badenhorst M, Saalmüller A, Daly JM, Ertl R, Stadler M, Puff C, de le Roi M, Baumgärtner W, Engelmann M, Brandner S, et al. An Equine Model for Vaccination against a Hepacivirus: Insights into Host Responses to E2 Recombinant Protein Vaccination and Subsequent Equine Hepacivirus Inoculation. Viruses. 2022; 14(7):1401. https://doi.org/10.3390/v14071401
Chicago/Turabian StyleBadenhorst, Marcha, Armin Saalmüller, Janet M. Daly, Reinhard Ertl, Maria Stadler, Christina Puff, Madeleine de le Roi, Wolfgang Baumgärtner, Michael Engelmann, Sabine Brandner, and et al. 2022. "An Equine Model for Vaccination against a Hepacivirus: Insights into Host Responses to E2 Recombinant Protein Vaccination and Subsequent Equine Hepacivirus Inoculation" Viruses 14, no. 7: 1401. https://doi.org/10.3390/v14071401
APA StyleBadenhorst, M., Saalmüller, A., Daly, J. M., Ertl, R., Stadler, M., Puff, C., de le Roi, M., Baumgärtner, W., Engelmann, M., Brandner, S., Junge, H. K., Pratscher, B., Volz, A., Saunier, B., Krey, T., Wittmann, J., Heelemann, S., Delarocque, J., Wagner, B., ... Cavalleri, J. -M. V. (2022). An Equine Model for Vaccination against a Hepacivirus: Insights into Host Responses to E2 Recombinant Protein Vaccination and Subsequent Equine Hepacivirus Inoculation. Viruses, 14(7), 1401. https://doi.org/10.3390/v14071401