

Microcalorimetry: A Novel Application to Measure In Vitro Phage Susceptibility of Staphylococcus aureus in Human Serum
 , ,
, ,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. S. aureus Isolates and Phage Cocktails

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Clonal Complex | MSSA/MRSA | Source | Accession Number | References |
|---|---|---|---|---|---|
| Mup15 | CC15 | MSSA | Nose, healthy carrier (The Netherlands, 1999–2001) | ERS12471228 | [34,35] |
| Mup3199 | CC25 | MSSA | Nose, healthy carrier (The Netherlands, 1999–2001) | ERS12471229 | [36] |
| Mup2723 | CC30 | MSSA | Nose, healthy carrier (The Netherlands, 1999–2001) | ERS12471230 | [36] |
| Mup2396 | CC45 | MSSA | Nose, healthy carrier (The Netherlands, 1999–2001) | ERS12471231 | [36] |
| SA2704 | CC72 | MSSA | Nose, healthy carrier (The Netherlands, 1999–2001) | ERS12471232 | [36] |
| MW2 (USA400) | CC1 | MRSA | Blood (USA, 1998) | NC_003923.1 | [37] |
| Mu50 | CC5 | MRSA | VISA *, surgical wound (Japan, 1997) | BA000017.4 | [38] |
| SAC042W (USA300) | CC8 | MRSA | Skin abscess (USA, 2006–2008) | ERS12471233 | [39,40] |
| M116 | CC8 | MRSA | Osteomyelitis (Indonesia, 2011) | ERS12471234 | [41] |
| Rww146 | CC398 | MRSA | Live-stock associated clinical isolate | ERS12471235 | [42,43] |
| RN4220 | CC8 | MSSA | Derivative of strain 8324-4 | NZ_AFGU00000000.1 | [28] |
| R5 | CC30 | MSSA | RIVM, Phage typing strain | ERS12471236 | [29] |
2.2. Single Phage Isolation and Production
2.3. Next Generation Sequencing and Data Analysis
2.3.1. S. aureus Isolates
2.3.2. Phage Cocktails and Single Phages
2.4. Spot Test
2.5. Optical Density (OD) Assay
2.6. Efficiency of Plating (EOP)
2.7. Microcalorimetry (MC)
2.7.1. Serum Used for Microcalorimetry
2.7.2. Phage Cocktails
2.7.3. Single Phages
3. Results
3.1. Genetic Characteristics of Selected Clinical S. aureus Strains and Phage Cocktails
3.2. Determination of Phage Susceptibility Using Conventional In Vitro Assays
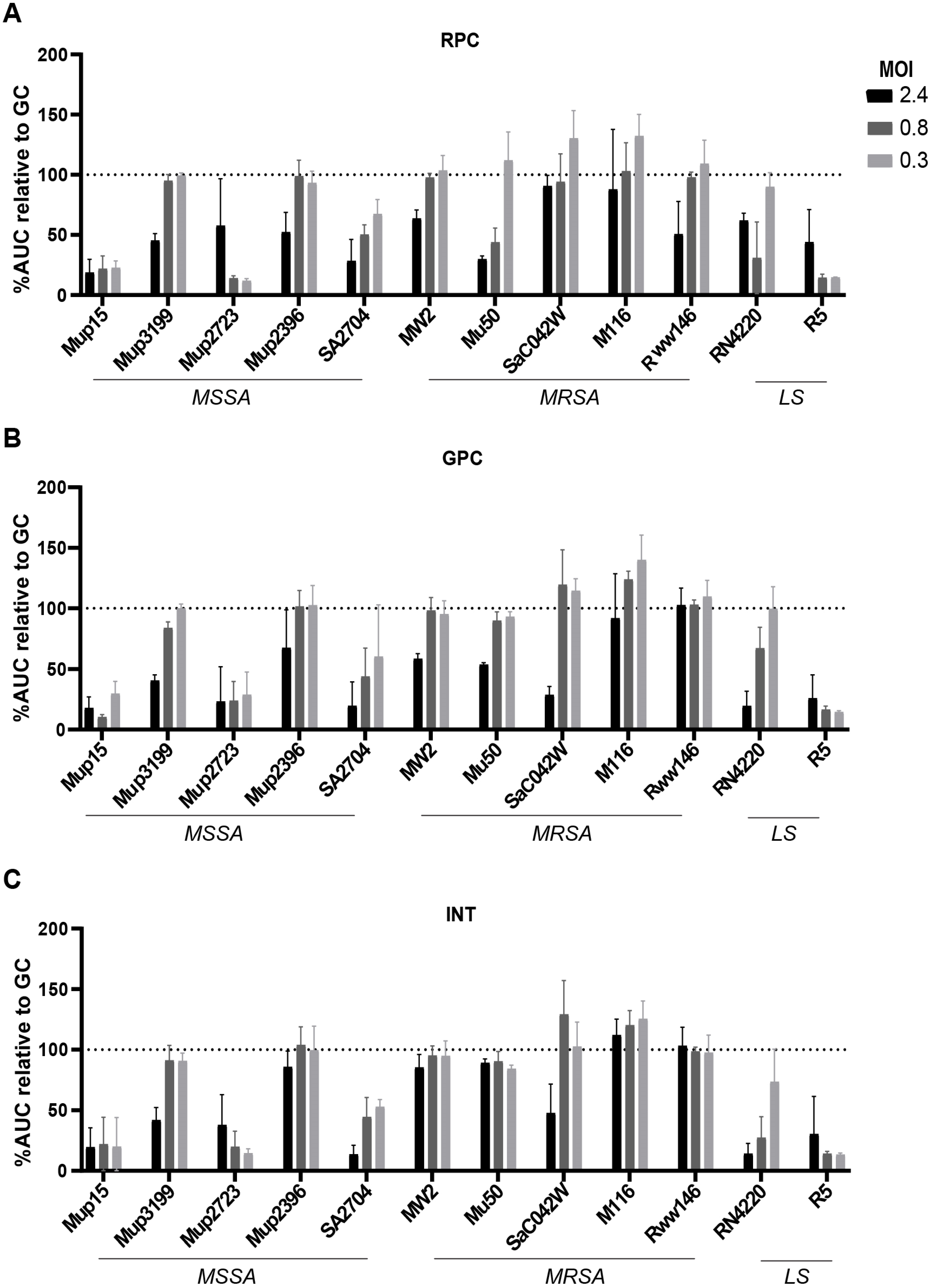
3.3. Susceptibility to Commercial Phage Cocktails in Human Serum Measured Using Microcalorimetry
3.4. Susceptibility to Single Phages Using the Conventional In Vitro Assays
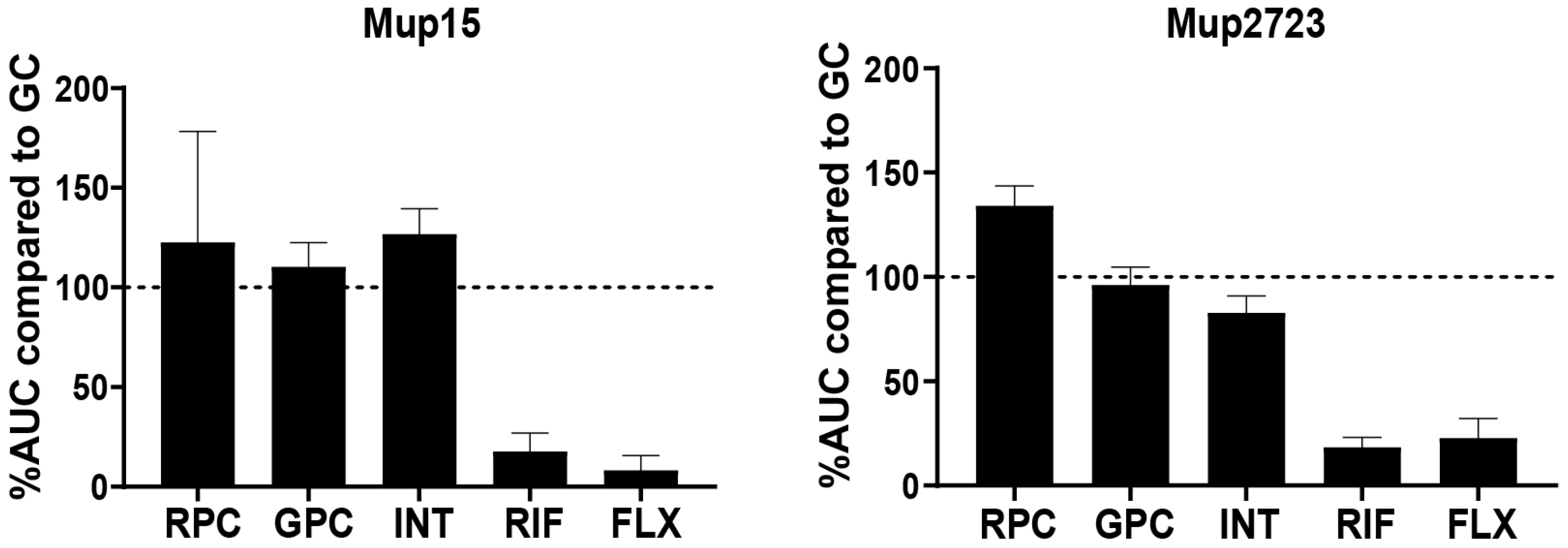
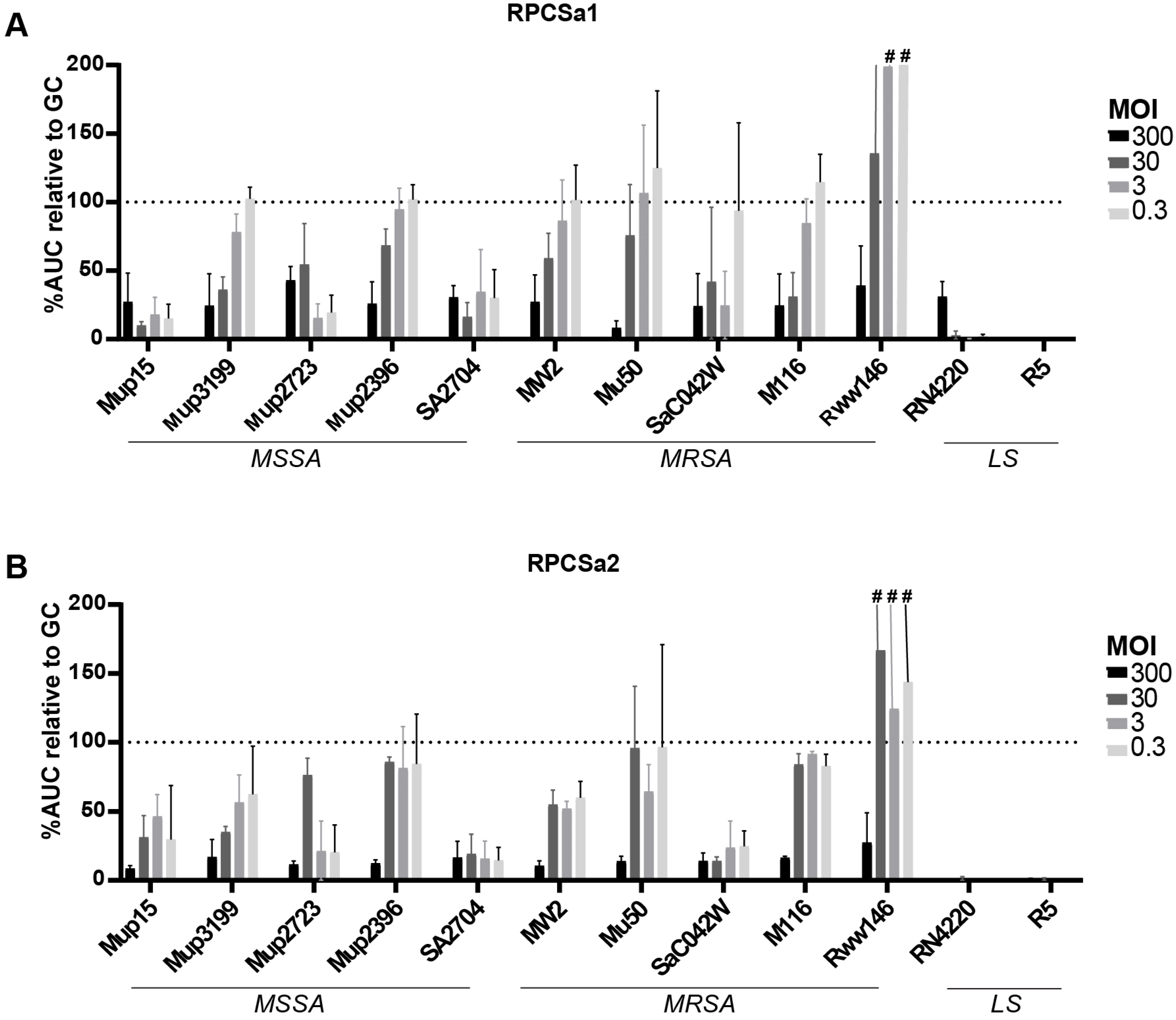
3.5. Susceptibility to Single Phages in Human Serum Measured Using Microcalorimetry
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, A.S.; De Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Prim. 2018, 4, 18033. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, J.A.; Holden, M.T.G. Staphylococcus aureus: Superbug, super genome? Trends Microbiol. 2004, 12, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Hassoun, A.; Linden, P.K.; Friedman, B. Incidence, prevalence, and management of MRSA bacteremia across patient populations—A review of recent developments in MRSA management and treatment. Crit. Care 2017, 21, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European economic area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delattre, R.; Seurat, J.; Haddad, F.; Nguyen, T.-T.; Gaborieau, B.; Kane, R.; Dufour, N.; Ricard, J.-D.; Guedj, J.; Debarbieux, L. Title combination of in vivo phage therapy data with in silico model highlights key parameters for treatment efficacy. Cell Rep. 2022, 39, 110825. [Google Scholar] [CrossRef]
- El Haddad, L.; Harb, C.P.; Gebara, M.A.; Stibich, M.A.; Chemaly, R.F. A systematic and critical review of bacteriophage therapy against multidrug-resistant ESKAPE organisms in humans. Clin. Infect. Dis. 2019, 69, 167–178. [Google Scholar] [CrossRef]
- Jamal, M.; Bukhari, S.M.A.U.S.; Andleeb, S.; Ali, M.; Raza, S.; Nawaz, M.A.; Hussain, T.; Rahman, S.u.; Shah, S.S.A. Bacteriophages: An overview of the control strategies against multiple bacterial infections in different fields. J. Basic Microbiol. 2019, 59, 123–133. [Google Scholar] [CrossRef]
- Fauconnier, A. Phage therapy regulation: From night to dawn. Viruses 2019, 11, 352. [Google Scholar] [CrossRef] [Green Version]
- Suh, G.A.; Lodise, T.P.; Tamma, P.D.; Knisely, J.M.; Alexander, J.; Aslam, S.; Barton, K.D.; Bizzell, E.; Totten, K.M.C.; Campbell, J.L.; et al. Considerations for the use of phage therapy in clinical practice. Antimicrob. Agents Chemother. 2022, 66, e02071-21. [Google Scholar] [CrossRef]
- Glonti, T.; Pirnay, J.P. In vitro techniques and measurements of phage characteristics that are important for phage therapy success. Viruses 2022, 14, 1490. [Google Scholar] [CrossRef]
- Cheung, A.L.; Bayer, A.S.; Zhang, G.; Gresham, H.; Xiong, Y.Q. Regulation of virulence determinants in vitro and in vivo in Staphylococcus aureus. FEMS Immunol. Med. Microbiol. 2004, 40, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Gerlach, D.; Du, X.; Larsen, J.; Stegger, M.; Kuhner, P.; Peschel, A.; Xia, G.; Winstel, V. An accessory wall teichoic acid glycosyltransferase protects Staphylococcus aureus from the lytic activity of podoviridae. Sci. Rep. 2015, 5, 17219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidenmaier, C.; Peschel, A. Teichoic acids and related cell-wall glycopolymers in gram-positive physiology and host interactions. Nat. Rev. Microbiol. 2008, 6, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Xia, G.; Luhachack, L.G.; Campbell, J.; Meredith, T.C.; Chen, C.; Winstel, V.; Gekeler, C.; Irazoqui, J.E.; Peschel, A.; et al. Methicillin resistance in Staphylococcus aureus requires glycosylated wall teichoic acids. Proc. Natl. Acad. Sci. USA 2012, 109, 18909–18914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistretta, N.; Brossaud, M.; Telles, F.; Sanchez, V.; Talaga, P.; Rokbi, B. Glycosylation of Staphylococcus aureus cell wall teichoic acid is influenced by environmental conditions. Sci. Rep. 2019, 9, 3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plumet, L.; Ahmad-Mansour, N.; Dunyach-Remy, C.; Kissa, K.; Sotto, A.; Lavigne, J.P.; Costechareyre, D.; Molle, V. Bacteriophage therapy for Staphylococcus aureus Infections: A review of animal models, treatments, and clinical trials. Front. Cell. Infect. Microbiol. 2022, 12, 808. [Google Scholar] [CrossRef]
- Kim, H.K.; Missiakas, D.; Schneewind, O. Mouse models for infectious diseases caused by Staphylococcus aureus. J. Immunol. Methods 2014, 410, 88–99. [Google Scholar] [CrossRef]
- Mrochen, D.M.; Fernandes de Oliveira, L.M.; Raafat, D.; Holtfreter, S. Staphylococcus aureus host tropism and its implications for murine infection models. Int. J. Mol. Sci. 2020, 21, 7061. [Google Scholar] [CrossRef]
- Doke, S.K.; Dhawale, S.C. Alternatives to animal testing: A review. Saudi Pharm. J. 2015, 23, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.D.; DeLeo, F.R. Staphylococcus aureus protein A promotes immune suppression. MBio 2013, 4, e00764-13. [Google Scholar] [CrossRef]
- Falugi, F.; Kim, H.K.; Missiakas, D.M.; Schneewind, O. Role of protein A in the evasion of host adaptive immune responses by Staphylococcus aureus. MBio 2013, 4, e00575-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultan, A.R.; Tavakol, M.; Lemmens-Den Toom, N.A.; Croughs, P.D.; Verkaik, N.J.; Verbon, A.; van Wamel, W.J.B. Real time monitoring of Staphylococcus aureus biofilm sensitivity towards antibiotics with isothermal microcalorimetry. PLoS ONE 2022, 17, e0260272. [Google Scholar] [CrossRef] [PubMed]
- Maskow, T.; Kiesel, B.; Schubert, T.; Yong, Z.; Harms, H.; Yao, J. Calorimetric real time monitoring of lambda prophage induction. J. Virol. Methods 2010, 168, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Tkhilaishvili, T.; Di Luca, M.; Abbandonato, G.; Maiolo, E.M.; Klatt, A.B.; Reuter, M.; Möncke-Buchner, E.; Trampuz, A. Real-time assessment of bacteriophage T3-derived antimicrobial activity against planktonic and biofilm-embedded Escherichia coli by isothermal microcalorimetry. Res. Microbiol. 2018, 169, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Tkhilaishvili, T.; Wang, L.; Tavanti, A.; Trampuz, A.; Di Luca, M. Antibacterial efficacy of two commercially available bacteriophage formulations, staphylococcal bacteriophage and PYO bacteriophage, against methicillin-resistant Staphylococcus aureus: Prevention and eradication of biofilm formation and control of a systemic infection of galleria Mellonella Larvae. Front. Microbiol. 2020, 11, 110. [Google Scholar] [CrossRef] [Green Version]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Van Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; et al. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- King, J.M.; Kulhankova, K.; Stach, C.S.; Vu, B.G.; Salgado-pabón, W. Phenotypes and virulence among Staphylococcus aureus USA100, USA200, USA300, USA400, and USA600 clonal lineages. Msphere 2016, 1, e00071-16. [Google Scholar] [CrossRef] [Green Version]
- Nair, D.; Memmi, G.; Hernandez, D.; Bard, J.; Beaume, M.; Gill, S.; Francois, P.; Cheung, A.L. Whole-genome sequencing of Staphylococcus aureus strain RN4220, a key laboratory strain used in virulence research, identifies mutations that affect not only virulence factors but also the fitness of the strain. J. Bacteriol. 2011, 193, 2332–2335. [Google Scholar] [CrossRef] [Green Version]
- Van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; Van Kessel, K.P.M.; Van Strijp, J.A.G. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on β-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Villarroel, J.; Larsen, M.V.; Kilstrup, M.; Nielsen, M. Metagenomic analysis of therapeutic PYO phage cocktails from 1997 to 2014. Viruses 2017, 9, 328. [Google Scholar] [CrossRef]
- Leitner, L.; Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Chkonia, I.; Rigvava, S.; Chkhotua, A.; Changashvili, G.; McCallin, S.; Schneider, M.P.; et al. Intravesical bacteriophages for treating urinary tract infections in patients undergoing transurethral resection of the prostate: A randomised, placebo-controlled, double-blind clinical trial. Lancet Infect. Dis. 2021, 21, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Leitner, L.; Sybesma, W.; Chanishvili, N.; Goderdzishvili, M.; Chkhotua, A.; Ujmajuridze, A.; Schneider, M.P.; Sartori, A.; Mehnert, U.; Bachmann, L.M.; et al. Bacteriophages for treating urinary tract infections in patients undergoing transurethral resection of the prostate: A randomized, placebo-controlled, double-blind clinical trial. BMC Urol. 2017, 17, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erol, H.B.; Kaskatepe, B.; Ozturk, S.; Safi Oz, Z. The comparison of lytic activity of isolated phage and commercial intesti bacteriophage on ESBL producer E. coli and determination of Ec_P6 phage efficacy with in vivo galleria Mellonella Larvae model. Microb. Pathog. 2022, 167, 105563. [Google Scholar] [CrossRef]
- Van Trijp, M.J.C.A.; Melles, D.C.; Snijders, S.V.; Wertheim, H.F.L.; Verbrugh, H.A.; van Belkum, A.; van Wamel, W.J. Genotypes, superantigen gene profiles, and presence of exfoliative toxin genes in clinical methicillin-susceptible Staphylococcus aureus isolates. Diagn. Microbiol. Infect. Dis. 2010, 66, 222–224. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, H.F.L.; Vos, M.C.; Ott, A.; Van Belkum, A.; Voss, A.; Kluytmans, J.A.J.W.; Van Keulen, P.H.J.; Vandenbroucke-Grauls, C.M.J.E.; Meester, M.H.M.; Verbrugh, H.A. Risk and outcome of nosocomial Staphylococcus aureus bacteraemia in nasal carriers versus non-carriers. Lancet 2004, 364, 703–705. [Google Scholar] [CrossRef]
- Wertheim, H.F.L.; van Leeuwen, W.B.; Snijders, S.; Vos, M.C.; Voss, A.; Vandenbroucke-Grauls, C.M.J.E.; Kluytmans, J.A.J.W.; Verbrugh, H.A.; van Belkum, A. Associations between Staphylococcus aureus genotype, infection, and in-hospital mortality: A nested case-control study. J. Infect. Dis. 2005, 192, 1196–1200. [Google Scholar] [CrossRef] [Green Version]
- Baba, T.; Takeuchi, F.; Kuroda, M.; Yuzawa, H.; Aoki, K.I.; Oguchi, A.; Nagai, Y.; Iwama, N.; Asano, K.; Naimi, T.; et al. Genome and virulence determinants of high virulence community-acquired MRSA. Lancet 2002, 359, 1819–1827. [Google Scholar] [CrossRef]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Kobayashi, N.; Cui, L.; Oguchi, A.; Aoki, K.I.; et al. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Den Reijer, P.M.; Haisma, E.M.; Lemmens-Den Toom, N.A.; Willemse, J.; Koning, R.A.; Demmers, J.A.A.; Dekkers, D.H.W.; Rijkers, E.; El Ghalbzouri, A.; Nibbering, P.H.; et al. Detection of alpha-toxin and other virulence factors in biofilms of Staphylococcus aureus on polystyrene and a human epidermalmodel. PLoS ONE 2016, 11, e0145722. [Google Scholar] [CrossRef]
- Faden, H.; Lesse, A.J.; Trask, J.; Hill, J.A.; Hess, D.J.; Dryja, D.; Lee, Y.H. Importance of colonization site in the current epidemic of staphylococcal skin abscesses. Pediatrics 2010, 125, e618–e624. [Google Scholar] [CrossRef]
- Sultan, A.R.; Swierstra, J.W.; Lemmens-den Toom, N.A.; Snijders, S.V.; Maňásková, S.H.; Verbon, A.; van Wamel, W.J.B. Production of staphylococcal complement inhibitor (SCIN) and other immune modulators during the early stages of Staphylococcus aureus biofilm formation in a mammalian cell culture medium. Infect. Immun. 2018, 86, e00352-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slingerland, B.C.G.C.; Tavakol, M.; McCarthy, A.J.; Lindsay, J.A.; Snijders, S.V.; Wagenaar, J.A.; van Belkum, A.; Vos, M.C.; Verbrugh, H.A.; van Wamel, W.J.B. Survival of Staphylococcus aureus ST398 in the human nose after artificial inoculation. PLoS ONE 2012, 7, e48896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, A.J.; van Wamel, W.; Vandendriessche, S.; Larsen, J.; Denis, O.; Garcia-Graells, C.; Uhlemann, A.C.; Lowy, F.D.; Skov, R.; Lindsay, J.A. Staphylococcus aureus CC398 clade associated with human-to-human transmission. Appl. Environ. Microbiol. 2012, 78, 8845–8848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luong, T.; Salabarria, A.C.; Edwards, R.A.; Roach, D.R. Standardized bacteriophage purification for personalized phage therapy. Nat. Protoc. 2020, 15, 2867–2890. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Gardner, S.N.; Slezak, T.; Hall, B.G. KSNP3.0: SNP Detection and phylogenetic analysis of genomes without genome alignment or reference genome. Bioinformatics 2015, 31, 2877–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (ITOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Moller, A.G.; Winston, K.; Ji, S.; Wang, J.; Hargita Davis, M.N.; Solís-Lemus, C.R.; Read, T.D. Genes influencing phage host range in Staphylococcus aureus on a species-wide scale. mSphere 2021, 6, e01263-20. [Google Scholar] [CrossRef]
- Cheng, R.; Huang, F.; Wu, H.; Lu, X.; Yan, Y.; Yu, B.; Wang, X.; Zhu, B. A nucleotide-sensing endonuclease from the Gabija bacterial defense system. Nucleic Acids Res. 2021, 49, 5216–5229. [Google Scholar] [CrossRef]
- Ka, D.; Oh, H.; Park, E.; Kim, J.H.; Bae, E. Structural and functional evidence of bacterial antiphage protection by thoeris defense system via NAD+ degradation. Nat. Commun. 2020, 11, 2816. [Google Scholar] [CrossRef]
- Payne, L.J.; Todeschini, T.C.; Wu, Y.; Perry, B.J.; Ronson, C.W.; Fineran, P.C.; Nobrega, F.L.; Jackson, S.A. Identification and classification of antiviral defence systems in bacteria and archaea with PADLOC reveals new system types. Nucleic Acids Res. 2021, 49, 10868–10878. [Google Scholar] [CrossRef] [PubMed]
- Atha, D.H.; Ingham, K.C. Mechanism of precipitation of proteins by polyethylene glycols. Analysis in terms of excluded volume. J. Biol. Chem. 1981, 256, 12108–12117. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Fu, L.; Niu, B.; Wu, S.; Wooley, J. Ultrafast clustering algorithms for metagenomic sequence analysis. Brief. Bioinform. 2012, 13, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-One FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive κ-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IQ Tree. Available online: http://www.iqtree.org/ (accessed on 1 June 2022).
- André Rambaut FigTree—v1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 4 July 2022).
- Hall, T. BioEdit: A user friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Clokie, M.R.J.; Kropinski, A.M. Bacteriophages; Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions; Human Press: Totowa, NJ, USA, 2009. [Google Scholar]
- Sigg, A.P.; Mariotti, M.; Grütter, A.E.; Lafranca, T.; Leitner, L.; Bonkat, G.; Braissant, O. A method to determine the efficacy of a commercial phage preparation against uropathogens in urine and artificial urine determined by isothermal microcalorimetry. Microorganisms 2022, 10, 845. [Google Scholar] [CrossRef]
- Dabul, A.N.G.; Camargo, I.L.B.C. Clonal complexes of Staphylococcus aureus: All mixed and together. FEMS Microbiol. Lett. 2014, 351, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Egido, J.E.; Costa, A.R.; Aparicio-Maldonado, C.; Haas, P.-J.; Brouns, S.J.J. Mechanisms and clinical importance of bacteriophage resistance. FEMS Microbiol. Rev. 2021, 46, fuab048. [Google Scholar] [CrossRef]
- Mirzaei, M.K.; Nilsson, A.S. Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar] [CrossRef]
- Shinde, P.; Stamatos, N.; Doub, J.B. Human plasma significantly reduces bacteriophage infectivity against Staphylococcus aureus clinical isolates. Cureus 2022, 14, e23777. [Google Scholar] [CrossRef] [PubMed]
- Carroll-Portillo, A.; Coffman, C.N.; Varga, M.G.; Alcock, J.; Singh, S.B.; Lin, H.C. Standard bacteriophage purification procedures cause loss in numbers and activity. Viruses 2021, 13, 328. [Google Scholar] [CrossRef] [PubMed]
- Erol, H.B.; Kaskatepe, B.; Bakkaloglu, Z.; Suzuk Yildiz, S. The evaluation of five commercial bacteriophage cocktails against methicillin-resistant Staphylococcus aureus isolated from Nasal Swab samples. Arch. Microbiol. 2021, 203, 5735–5743. [Google Scholar] [CrossRef]
- Neguţ, A.C.; Chifiriuc, M.C.; Săndulescu, O.; Streinu-Cercel, A.; Oprea, M.; Drăgulescu, E.C.; Gheorghe, I.; Berciu, I.; Coralia, B.; Popa, M.; et al. Bacteriophage-driven inhibition of biofilm formation in Staphylococcus strains from patients attending a Romanian reference center for infectious diseases. FEMS Microbiol. Lett. 2016, 363, fnw193. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, S.; Coffey, A.; Edwards, R.; Meaney, W.; Fitzgerald, G.F.; Ross, R.P. Genome of staphylococcal phage K: A new lineage of Myoviridae infecting gram-positive bacteria with a low G+C content. J. Bacteriol. 2004, 186, 2862–2871. [Google Scholar] [CrossRef] [Green Version]
- Azam, A.H.; Tanji, Y. Peculiarities of Staphylococcus aureus phages and their possible application in phage therapy. Appl. Microbiol. Biotechnol. 2019, 103, 4279–4289. [Google Scholar] [CrossRef] [PubMed]
- Moller, A.G.; Lindsay, J.A.; Read, T.D.; Johnson, K.N. Determinants of phage host range in Staphylococcus species. Appl. Environ. Microbiol. 2019, 85, 209–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dąbrowska, K.; Abedon, S.T. Pharmacologically aware phage therapy: Pharmacodynamic and pharmacokinetic obstacles to phage antibacterial action in animal and human bodies. Microbiol. Mol. Biol. Rev. 2019, 83, e00012-19. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.; Hall, A.; Zahn, H.; Eisenberg, M.; Erickson, S. Bacteriophage-based detection of Staphylococcus aureus in human serum. Viruses 2022, 14, 1748. [Google Scholar] [CrossRef] [PubMed]
- Biswas, B. Propagation of S. aureus phage K in presence of human blood. Biomed. J. Sci. Tech. Res. 2019, 18, 11–13. [Google Scholar] [CrossRef]
- Łusiak-Szelachowska, M.; Zaczek, M.; Weber-Dabrowska, B.; Międzybrodzki, R.; Kłak, M.; Fortuna, W.; Letkiewicz, S.; Rogóz, P.; Szufnarowski, K.; Jończyk-Matysiak, E.; et al. Phage neutralization by sera of patients receiving phage therapy. Viral Immunol. 2014, 27, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodyra-Stefaniak, K.; Kaźmierczak, Z.; Majewska, J.; Sillankorva, S.; Miernikiewicz, P.; Międzybrodzki, R.; Górski, A.; Azeredo, J.; Lavigne, R.; Lecion, D.; et al. Natural and induced antibodies against phages in humans: Induction kinetics and immunogenicity for structural proteins of PB1-related phages. Phage 2020, 1, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Fabijan, A.P.; Lin, R.C.Y.; Ho, J.; Maddocks, S.; Iredell, J.R. Safety and tolerability of bacteriophage therapy in severe Staphylococcus aureus infection. bioRxiv 2019. bioRxiv: 619999. [Google Scholar] [CrossRef]







| MSSA Strains | MRSA Strains | Lab Strains | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genes * | Mup15 | Mup3199 | Mup2723 | Mup2396 | SA2704 | MW2 | Mu50 | SaC042W | M116 | Rww146 | RN4220 | R5 | |
| isdB | + # | + | + | + | + | + | Moller et al. (2021) [48] | ||||||
| mrpF | + | + | + | + | + | + | |||||||
| relA | + | + | + | + | + | + | |||||||
| tarP | + | ||||||||||||
| phoR | + | + | + | + | + | + | |||||||
| tarS | + | + | + | + | + | + | |||||||
| tarM | + | + | + | + | + | + | |||||||
| fmtC | + | + | + | + | + | + | |||||||
| tagH | + | + | + | + | + | + | |||||||
| trpA | + | + | + | + | + | + | |||||||
| tarJ | + | + | + | + | + | + | |||||||
| sodM2 | + | + | + | + | + | + | |||||||
| hsdR | + | + | + | + | + | + | + | + | |||||
| hsdM | + | + | + | + | + | + | + | + | |||||
| sau3AIR | + | ||||||||||||
| gajA | + | PADLOC [51] | |||||||||||
| gajB | + | ||||||||||||
| drt4 | |||||||||||||
| thsB | + | ||||||||||||
| thsA | + | ||||||||||||
| lmuA | + | ||||||||||||
| lmuB | + | ||||||||||||
| avs2 | + | + | |||||||||||
| RPC | GPC | INT | RPCSa2 | RPCSa1 | ||
|---|---|---|---|---|---|---|
| MSSA strains | Mup15 | ++ # | ++ | ++ | ++++ | ++++ |
| Mup3199 | +++ | +++ | ||||
| Mup2723 | ++ | ++ | ++ | ++++ | ++++ | |
| Mup2396 | ||||||
| SA2704 | + | + | + | +++ | +++ | |
| MRSA strains | MW2 | + | + | +++ | +++ | |
| Mu50 | ||||||
| SaC042W | ++ | ++ | +++ | +++ | ||
| M116 | ++ | + | +++ | |||
| Rww146 | ||||||
| Lab strains | RN4220 | ++ | ++ | +++ | ++++ | |
| R5 | + | + | ++ | +++ | +++ | |
| RPC | GPC | INT | RPCSa2 | RPCSa1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ST | OD | EOP | ST | OD | EOP | ST | OD | EOP | ST | OD | EOP | ST | OD | EOP | |
| Mup3199 | -# | + | + | - | + | + | - | + | + | ||||||
| Mup2396 | - | + | - | - | + | + | - | + | + | ||||||
| MW2 | - | + | + | + | - | + | |||||||||
| Mu50 | - | + | - | - | + | - | - | + | + | - | + | + | |||
| SaC042W | + | - | + | - | + | + | |||||||||
| M116 | + | - | + | + | - | + | - | + | + | ||||||
| Rww146 | - | + | - | - | + | - | - | + | - | ||||||
| RN4220 | - | + | + | ||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molendijk, M.M.; Phan, M.V.T.; Bode, L.G.M.; Strepis, N.; Prasad, D.K.; Worp, N.; Nieuwenhuijse, D.F.; Schapendonk, C.M.E.; Boekema, B.K.H.L.; Verbon, A.; et al. Microcalorimetry: A Novel Application to Measure In Vitro Phage Susceptibility of Staphylococcus aureus in Human Serum. Viruses 2023, 15, 14. https://doi.org/10.3390/v15010014
Molendijk MM, Phan MVT, Bode LGM, Strepis N, Prasad DK, Worp N, Nieuwenhuijse DF, Schapendonk CME, Boekema BKHL, Verbon A, et al. Microcalorimetry: A Novel Application to Measure In Vitro Phage Susceptibility of Staphylococcus aureus in Human Serum. Viruses. 2023; 15(1):14. https://doi.org/10.3390/v15010014
Chicago/Turabian StyleMolendijk, Michèle M., My V. T. Phan, Lonneke G. M. Bode, Nikolas Strepis, Divyae K. Prasad, Nathalie Worp, David F. Nieuwenhuijse, Claudia M. E. Schapendonk, Bouke K. H. L. Boekema, Annelies Verbon, and et al. 2023. "Microcalorimetry: A Novel Application to Measure In Vitro Phage Susceptibility of Staphylococcus aureus in Human Serum" Viruses 15, no. 1: 14. https://doi.org/10.3390/v15010014
APA StyleMolendijk, M. M., Phan, M. V. T., Bode, L. G. M., Strepis, N., Prasad, D. K., Worp, N., Nieuwenhuijse, D. F., Schapendonk, C. M. E., Boekema, B. K. H. L., Verbon, A., Koopmans, M. P. G., Graaf, M. d., & van Wamel, W. J. B. (2023). Microcalorimetry: A Novel Application to Measure In Vitro Phage Susceptibility of Staphylococcus aureus in Human Serum. Viruses, 15(1), 14. https://doi.org/10.3390/v15010014