

Phage K gp102 Drives Temperature-Sensitive Antibacterial Activity on USA300 MRSA
 , and
, and
Abstract
:1. Introduction
2. Methods
2.1. Bacteria and Bacteriophages
2.2. Phage Production and Quantification
2.3. Isolation of Phage K Mutants
2.4. Frequency of gp102 Mutations
2.5. Phage Sequencing and Analysis
2.6. Bacterial Sequencing and Analysis
2.7. Bacterial Growth Suppression in Broth
2.8. Bacterial Killing in Broth
2.9. Molecular Modeling
2.10. Adsorption Assays
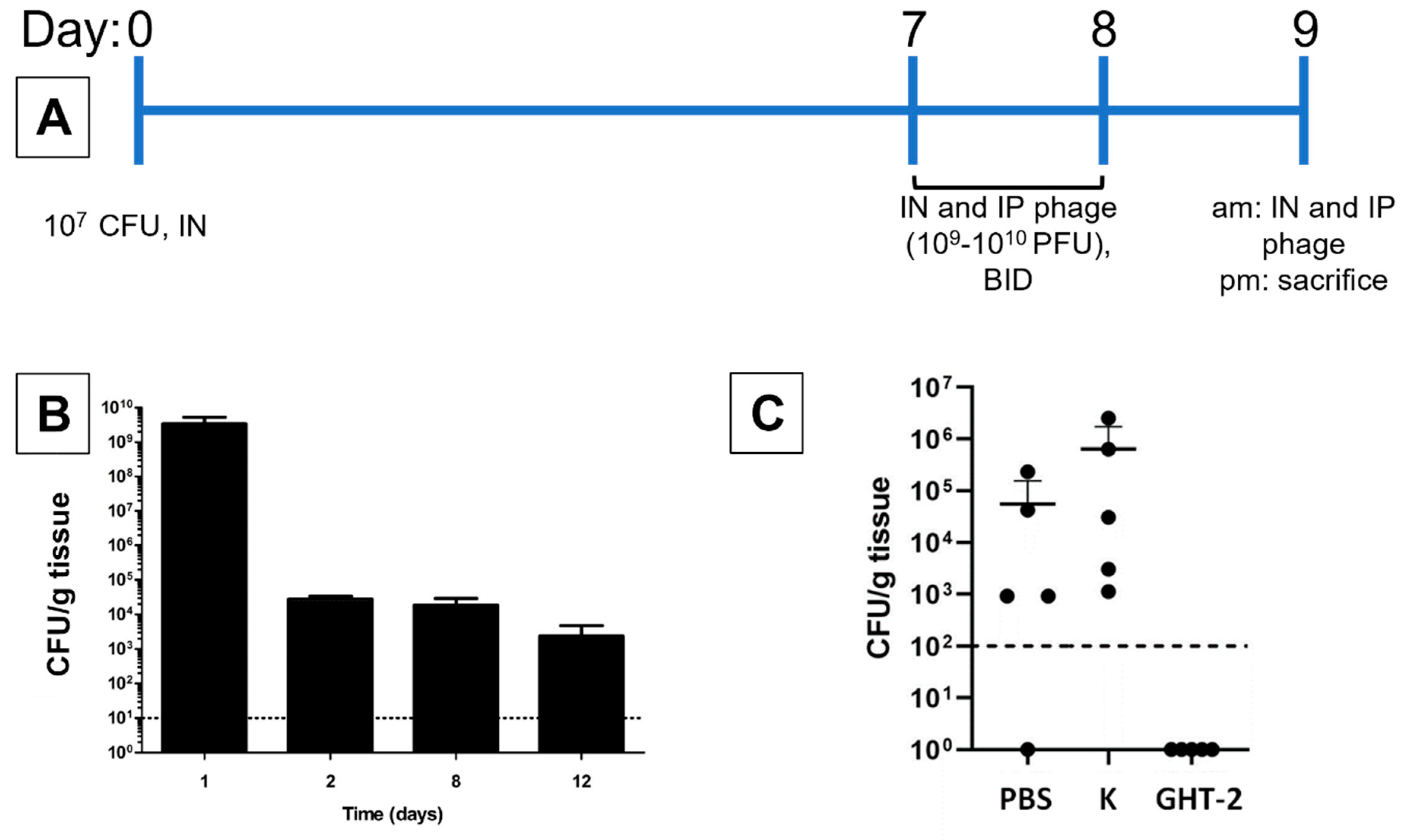
2.11. Animal Experiments
2.12. gp102 Expression
3. Results
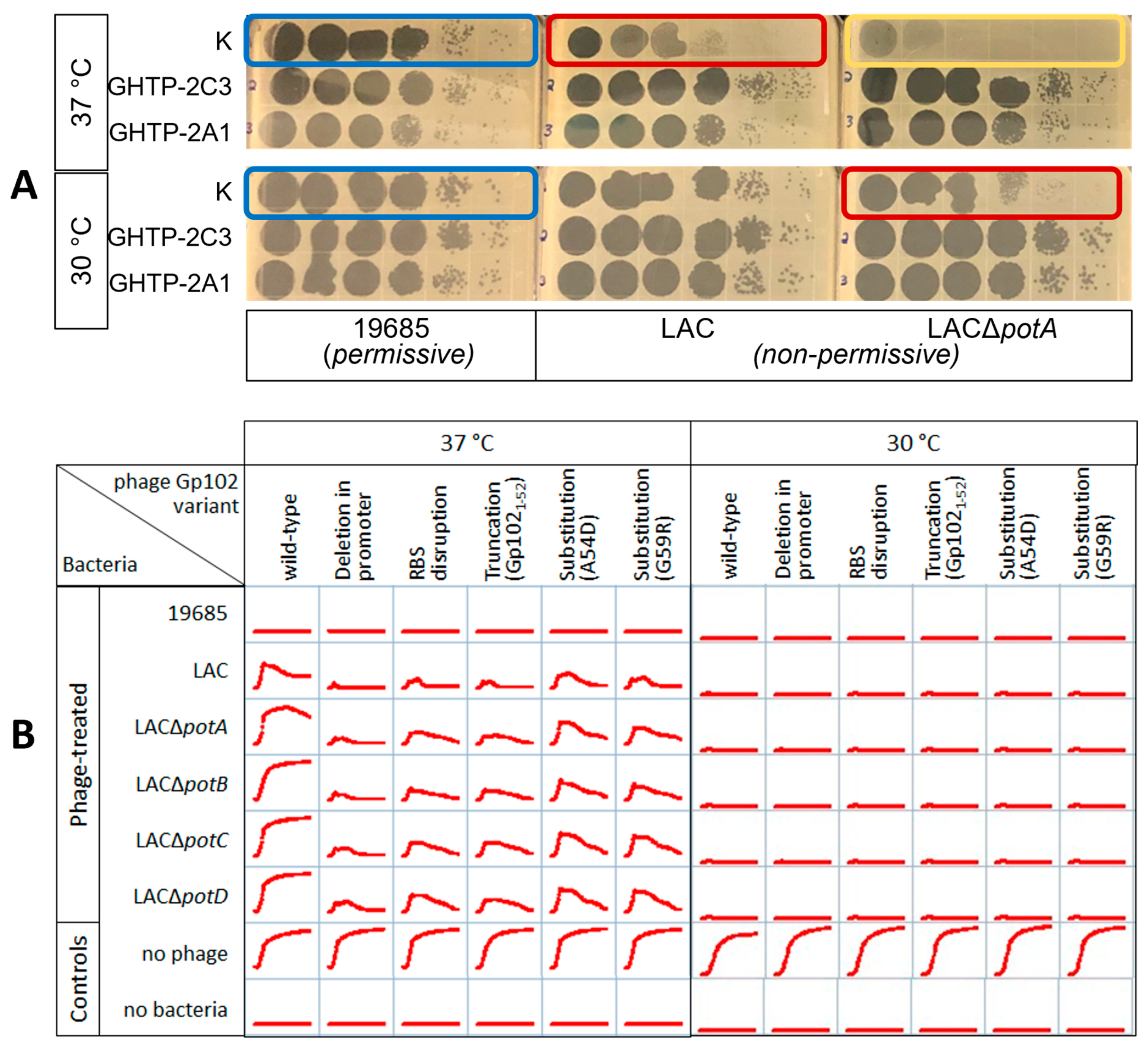
3.1. Isolation and Identification of Phage K gp102 Mutants
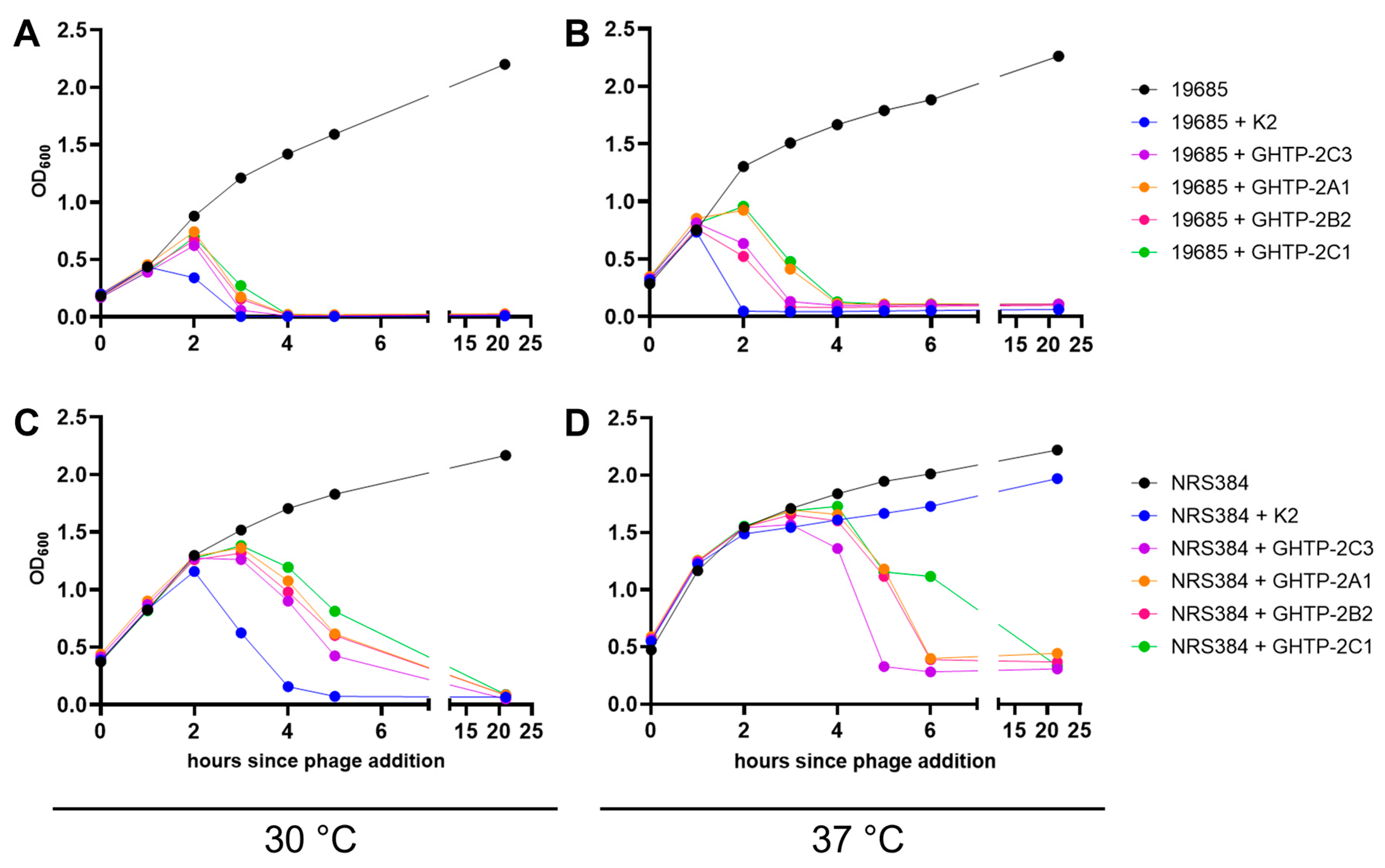
3.2. In Vitro Antibacterial Phenotypes of Phage K gp102 Mutants
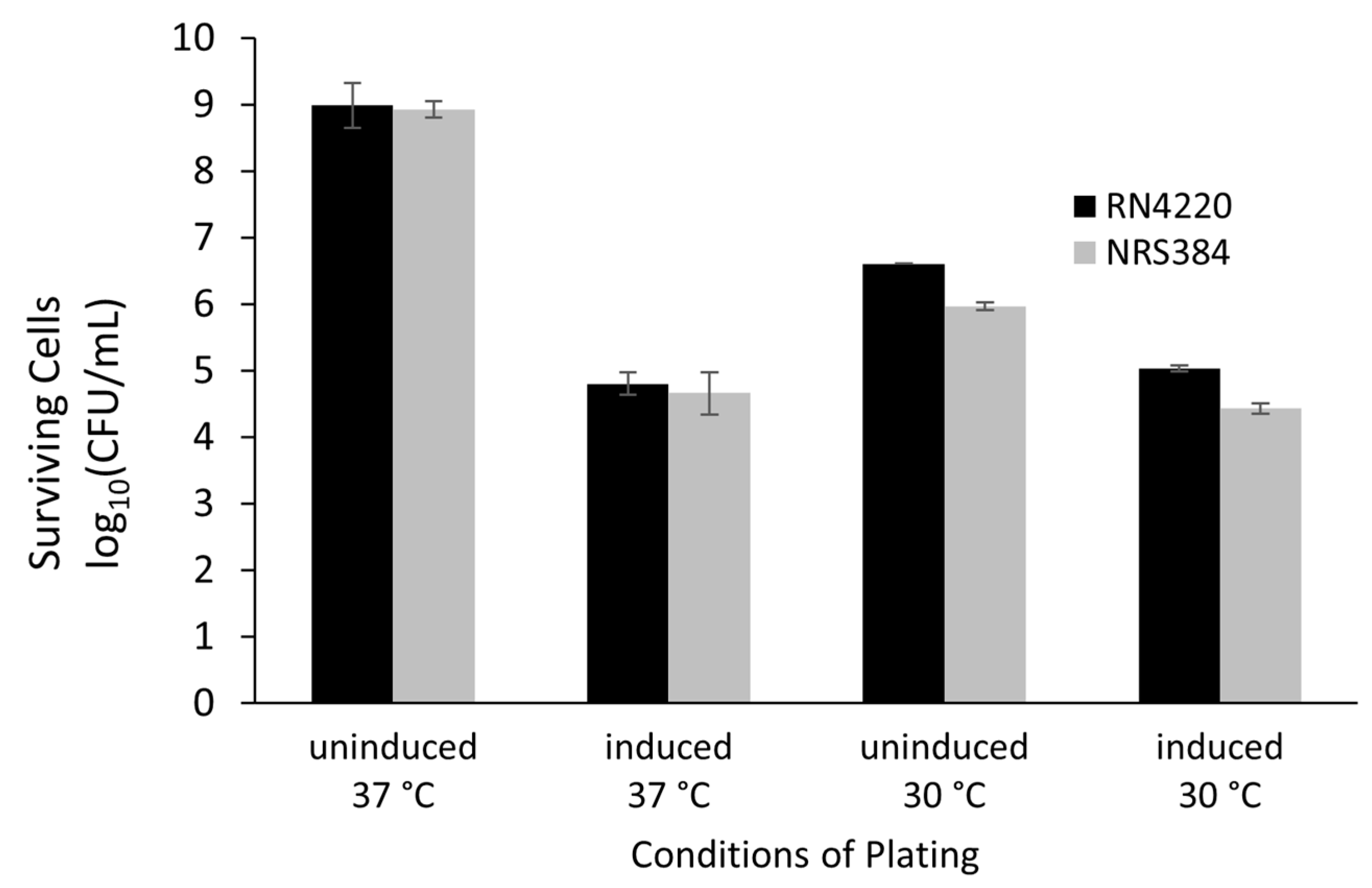
3.3. In Vivo Antibacterial Activity of K and Its gp102 Mutants
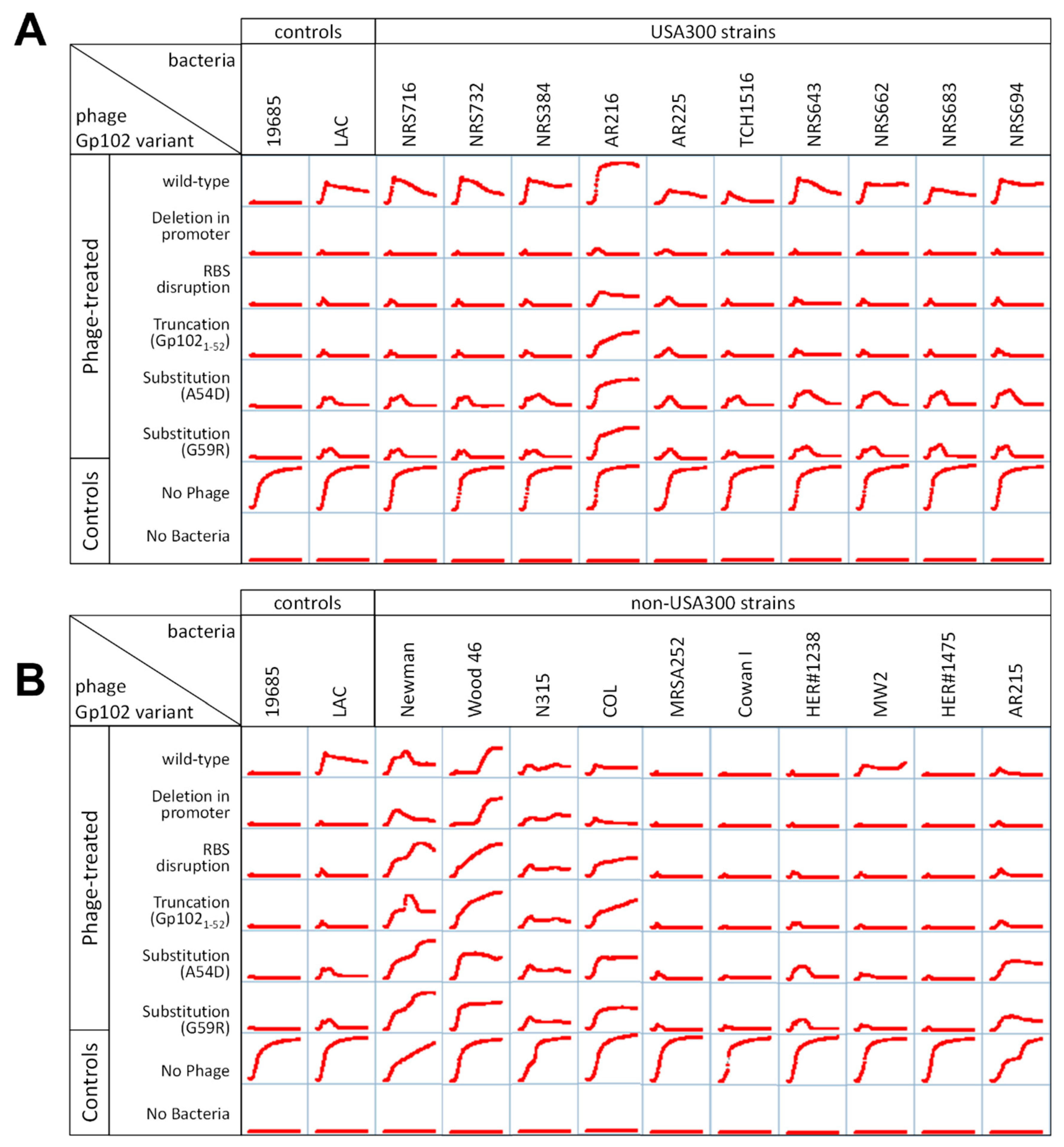
3.4. Correlation of gp102-Mediated Temperature-Sensitivity with S. aureus Lineage
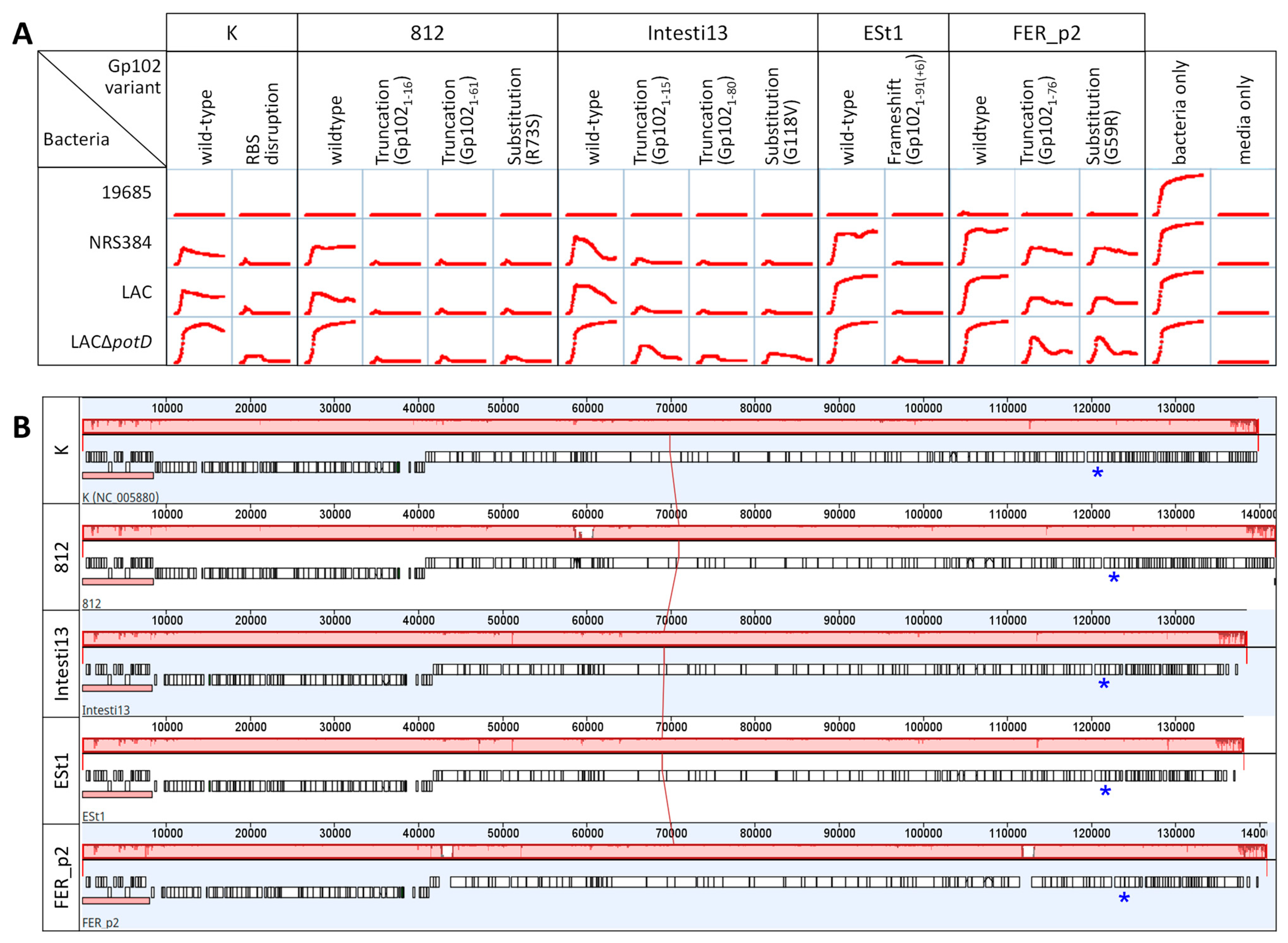
3.5. Effect of gp102 Mutations in Other Kayvirus Phages
3.6. Effects of gp102 Mutations on Phage Infection Cycle
3.7. Gene Expression
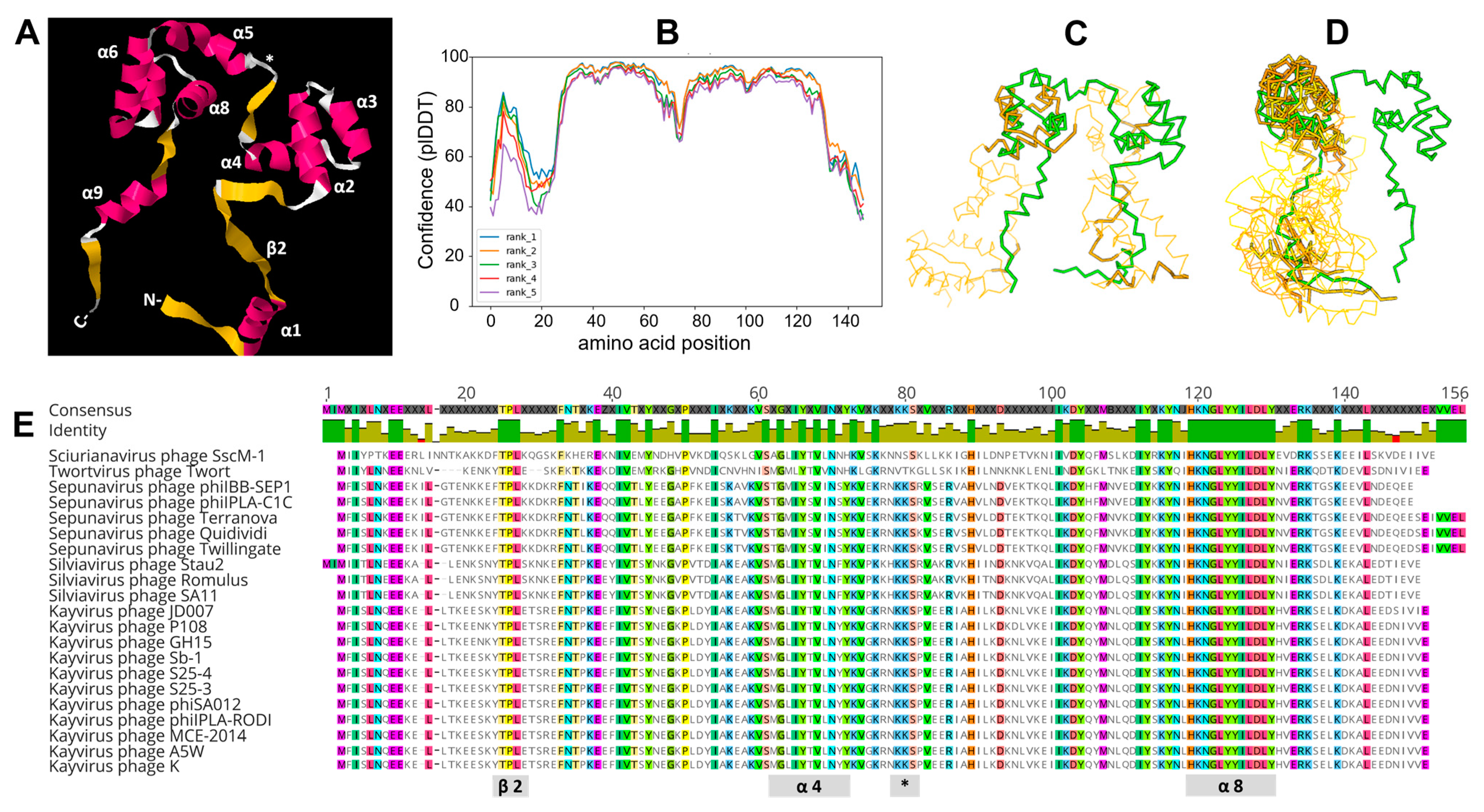
3.8. Molecular Modeling
3.9. Taxonomic Distribution of gp102 Homologs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- LaPlante, K.L.; Rybak, M.J. Impact of high-inoculum Staphylococcus aureus on the activities of nafcillin, vancomycin, linezolid, and daptomycin, alone and in combination with gentamicin, in an in vitro pharmacodynamic model. Antimicrob. Agents Chemother. 2004, 48, 4665–4672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haney, E.F.; Hancock, R.E.W. Addressing Antibiotic Failure—Beyond Genetically Encoded Antimicrobial Resistance. Front. Drug Discov. 2022, 2, 892975. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV). ICTV Master Species List 2021.v1 (MSL#37). 2021. Available online: https://ictv.global/msl (accessed on 15 December 2022).
- Hatfull, G.F. Chapter 7—The Secret Lives of Mycobacteriophages. In Advances in Virus Research; Łobocka, M., Szybalski, W.T., Eds.; Academic Press: Waltham, MA, USA, 2012; Volume 82, pp. 179–288. [Google Scholar]
- Kwan, T.; Liu, J.; DuBow, M.; Gros, P.; Pelletier, J. The complete genomes and proteomes of 27 Staphylococcus aureus bacteriophages. Proc. Natl. Acad. Sci. USA 2005, 102, 5174–5179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berjon-Otero, M.; Lechuga, A.; Mehla, J.; Uetz, P.; Salas, M.; Redrejo-Rodriguez, M. Bam35 Tectivirus Intraviral Interaction Map Unveils New Function and Localization of Phage ORFan Proteins. J. Virol. 2017, 91, e00870-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehla, J.; Dedrick, R.M.; Caufield, J.H.; Wagemans, J.; Sakhawalkar, N.; Johnson, A.; Hatfull, G.F.; Uetz, P. Virus-host protein-protein interactions of mycobacteriophage Giles. Sci. Rep. 2017, 7, 16514. [Google Scholar] [CrossRef] [Green Version]
- Wagemans, J.; Blasdel, B.G.; Van den Bossche, A.; Uytterhoeven, B.; De Smet, J.; Paeshuyse, J.; Cenens, W.; Aertsen, A.; Uetz, P.; Delattre, A.S.; et al. Functional elucidation of antibacterial phage ORFans targeting Pseudomonas aeruginosa. Cell. Microbiol. 2014, 16, 1822–1835. [Google Scholar] [CrossRef]
- Liu, J.; Dehbi, M.; Moeck, G.; Arhin, F.; Bauda, P.; Bergeron, D.; Callejo, M.; Ferretti, V.; Ha, N.; Kwan, T.; et al. Antimicrobial drug discovery through bacteriophage genomics. Nat. Biotechnol. 2004, 22, 185–191. [Google Scholar] [CrossRef]
- Dedrick, R.M.; Marinelli, L.J.; Newton, G.L.; Pogliano, K.; Pogliano, J.; Hatfull, G.F. Functional requirements for bacteriophage growth: Gene essentiality and expression in mycobacteriophage Giles. Mol. Microbiol. 2013, 88, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Fischer, D. Identification and investigation of ORFans in the viral world. BMC Genom. 2008, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, M.; Chastanet, A.; Debarbouille, M. New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, gram-positive bacteria. Appl. Environ. Microbiol. 2004, 70, 6887–6891. [Google Scholar] [CrossRef]
- Prabhakara, R.; Foreman, O.; De Pascalis, R.; Lee, G.M.; Plaut, R.D.; Kim, S.Y.; Stibitz, S.; Elkins, K.L.; Merkel, T.J. Epicutaneous model of community-acquired Staphylococcus aureus skin infections. Infect. Immun. 2013, 81, 1306–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, P.J. Genetic Manipulation of Staphylococcus aureus. In Staphylococcus: Molecular Genetics; Lindsay, J.A., Ed.; Caister Academic Press: Norfolk, UK, 2008; pp. 89–129. [Google Scholar]
- David, M.Z.; Taylor, A.; Lynfield, R.; Boxrud, D.J.; Short, G.; Zychowski, D.; Boyle-Vavra, S.; Daum, R.S. Comparing pulsed-field gel electrophoresis with multilocus sequence typing, spa typing, staphylococcal cassette chromosome mec (SCCmec) typing, and PCR for panton-valentine leukocidin, arcA, and opp3 in methicillin-resistant Staphylococcus aureus isolates at a U.S. Medical Center. J. Clin. Microbiol. 2013, 51, 814–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strauß, L.; Stegger, M.; Akpaka, P.E.; Alabi, A.; Breurec, S.; Coombs, G.; Egyir, B.; Larsen, A.R.; Laurent, F.; Monecke, S.; et al. Origin, evolution, and global transmission of community-acquired Staphylococcus aureus ST8. Proc. Natl. Acad. Sci. USA 2017, 114, E10596–E10604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murai, T.; Okazaki, K.; Kinoshita, K.; Uehara, Y.; Zuo, H.; Lu, Y.; Ono, Y.; Sasaki, T.; Hiramatsu, K.; Horikoshi, Y. Comparison of USA300 with non-USA300 methicillin-resistant Staphylococcus aureus in a neonatal intensive care unit. Int. J. Infect. Dis. 2019, 79, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Gabler, F.; Nam, S.Z.; Till, S.; Mirdita, M.; Steinegger, M.; Soding, J.; Lupas, A.N.; Alva, V. Protein Sequence Analysis Using the MPI Bioinformatics Toolkit. Curr. Protoc. Bioinform. 2020, 72, e108. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Holm, L. DALI and the persistence of protein shape. Protein Sci. 2020, 29, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Bae, T.; Glass, E.M.; Schneewind, O.; Missiakas, D. Generating a collection of insertion mutations in the Staphylococcus aureus genome using bursa aurealis. Methods Mol. Biol. 2008, 416, 103–116. [Google Scholar] [CrossRef]
- Saez Moreno, D.; Visram, Z.; Mutti, M.; Restrepo-Cordoba, M.; Hartmann, S.; Kremers, A.I.; Tisakova, L.; Schertler, S.; Wittmann, J.; Kalali, B.; et al. epsilon(2)-Phages Are Naturally Bred and Have a Vastly Improved Host Range in Staphylococcus aureus over Wild Type Phages. Pharmaceuticals 2021, 14, 325. [Google Scholar] [CrossRef]
- Gallet, R.; Kannoly, S.; Wang, I.N. Effects of bacteriophage traits on plaque formation. BMC Microbiol. 2011, 11, 181. [Google Scholar] [CrossRef]
- Singh, S.K.; Sabatinos, S.; Forsburg, S.; Bastia, D. Regulation of replication termination by Reb1 protein-mediated action at a distance. Cell 2010, 142, 868–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, R.; Choudhury, M.; Zaman, S.; Singh, S.; Santosh, V.; Bastia, D.; Escalante, C.R. Functional architecture of the Reb1-Ter complex of Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 2016, 113, E2267–E2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barylski, J.; Enault, F.; Dutilh, B.E.; Schuller, M.B.; Edwards, R.A.; Gillis, A.; Klumpp, J.; Knezevic, P.; Krupovic, M.; Kuhn, J.H.; et al. Analysis of Spounaviruses as a Case Study for the Overdue Reclassification of Tailed Phages. Syst. Biol. 2020, 69, 110–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehman, S.M.; Mearns, G.; Rankin, D.; Cole, R.A.; Smrekar, F.; Branston, S.D.; Morales, S. Design and Preclinical Development of a Phage Product for the Treatment of Antibiotic-Resistant Staphylococcus aureus Infections. Viruses 2019, 11, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botka, T.; Pantůček, R.; Mašlaňová, I.; Benešík, M.; Petráš, P.; Růžičková, V.; Havlíčková, P.; Varga, M.; Žemličková, H.; Koláčková, I.; et al. Lytic and genomic properties of spontaneous host-range Kayvirus mutants prove their suitability for upgrading phage therapeutics against staphylococci. Sci. Rep. 2019, 9, 5475. [Google Scholar] [CrossRef] [Green Version]
- Estrella, L.A.; Quinones, J.; Henry, M.; Hannah, R.M.; Pope, R.K.; Hamilton, T.; Teneza-Mora, N.; Hall, E.; Biswajit, B. Characterization of novel Staphylococcus aureus lytic phage and defining their combinatorial virulence using the OmniLog(R) system. Bacteriophage 2016, 6, e1219440. [Google Scholar] [CrossRef] [Green Version]
- Ajuebor, J.; Buttimer, C.; Arroyo-Moreno, S.; Chanishvili, N.; Gabriel, E.M.; O’Mahony, J.; McAuliffe, O.; Neve, H.; Franz, C.; Coffey, A. Comparison of Staphylococcus Phage K with Close Phage Relatives Commonly Employed in Phage Therapeutics. Antibiotics 2018, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Jensen, K.C.; Hair, B.B.; Wienclaw, T.M.; Murdock, M.H.; Hatch, J.B.; Trent, A.T.; White, T.D.; Haskell, K.J.; Berges, B.K. Isolation and Host Range of Bacteriophage with Lytic Activity against Methicillin-Resistant Staphylococcus aureus and Potential Use as a Fomite Decontaminant. PLoS ONE 2015, 10, e0131714. [Google Scholar] [CrossRef]
- Ma, X.X.; Ito, T.; Tiensasitorn, C.; Jamklang, M.; Chongtrakool, P.; Boyle-Vavra, S.; Daum, R.S.; Hiramatsu, K. Novel type of staphylococcal cassette chromosome mec identified in community-acquired methicillin-resistant Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2002, 46, 1147–1152. [Google Scholar] [CrossRef] [Green Version]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A.; Mongodin, E.F.; et al. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Bachrach, U.; Fischer, R.; Klein, I. Occurrence of polyamines in coliphages T5, phiX174 and in phage-infected bacteria. J. Gen. Virol. 1975, 26, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, B.; Chakravorty, M. Effect of spermidine on bacteriophage P22 infection. J. Virol. 1978, 28, 736–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbe, J.; Bunk, B.; Rohde, M.; Schobert, M. Sequencing and characterization of Pseudomonas aeruginosa phage JG004. BMC Microbiol. 2011, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Joshi, G.S.; Spontak, J.S.; Klapper, D.G.; Richardson, A.R. Arginine catabolic mobile element encoded speG abrogates the unique hypersensitivity of Staphylococcus aureus to exogenous polyamines. Mol. Microbiol. 2011, 82, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, B.N.; Dubin, D.T. The role of polyamines in the neutralization of bacteriophage deoxyribonucleic acid. J. Biol. Chem. 1960, 235, 769–775. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage K Isolate | Genetic Change 1 | Predicted Effect on Gp102 | |
|---|---|---|---|
| Location in Genome (Location Relative to ORF) | CDS Change | ||
| K2 2 | 120766…121209 (1…444) | None (WT) | None (WT) |
| GHTP-2C3 | 120721 120732…(−45…−33) | -TTATGATATAGT (deletion in promoter) | Impaired transcription |
| GHTP-2A1 | 120753 (−13) | G → A (RBS disruption) | Impaired translation |
| GHTP-2B2 | 120922 (157) | G → T (ochre mutation) | Truncation after amino acid 52 |
| GHTP-2C1 | 120926 (161) | C → A | A54D |
| GHTP-2D1 | 120940 (175) | G → A | G59R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehman, S.M.; Kongari, R.; Glass, A.M.; Koert, M.; Ray, M.D.; Plaut, R.D.; Stibitz, S. Phage K gp102 Drives Temperature-Sensitive Antibacterial Activity on USA300 MRSA. Viruses 2023, 15, 17. https://doi.org/10.3390/v15010017
Lehman SM, Kongari R, Glass AM, Koert M, Ray MD, Plaut RD, Stibitz S. Phage K gp102 Drives Temperature-Sensitive Antibacterial Activity on USA300 MRSA. Viruses. 2023; 15(1):17. https://doi.org/10.3390/v15010017
Chicago/Turabian StyleLehman, Susan M., Rohit Kongari, Adam M. Glass, Matthew Koert, Melissa D. Ray, Roger D. Plaut, and Scott Stibitz. 2023. "Phage K gp102 Drives Temperature-Sensitive Antibacterial Activity on USA300 MRSA" Viruses 15, no. 1: 17. https://doi.org/10.3390/v15010017
APA StyleLehman, S. M., Kongari, R., Glass, A. M., Koert, M., Ray, M. D., Plaut, R. D., & Stibitz, S. (2023). Phage K gp102 Drives Temperature-Sensitive Antibacterial Activity on USA300 MRSA. Viruses, 15(1), 17. https://doi.org/10.3390/v15010017