
Characterization of a Fungal Virus Representing a Novel Genus in the Family Alphaflexiviridae


 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. S. sclerotiorum Isolates and Culture Conditions
2.2. Total-RNA Extraction and RT–PCR
2.3. Terminal-Sequence Cloning of SsAFV1
2.4. Structural and Phylogenetic Analyses
2.5. Viral-Particle Purification and Observation of SsAFV1
2.6. Biological Characteristics of AHS31 Strain
2.7. Statistical Analyses
3. Results
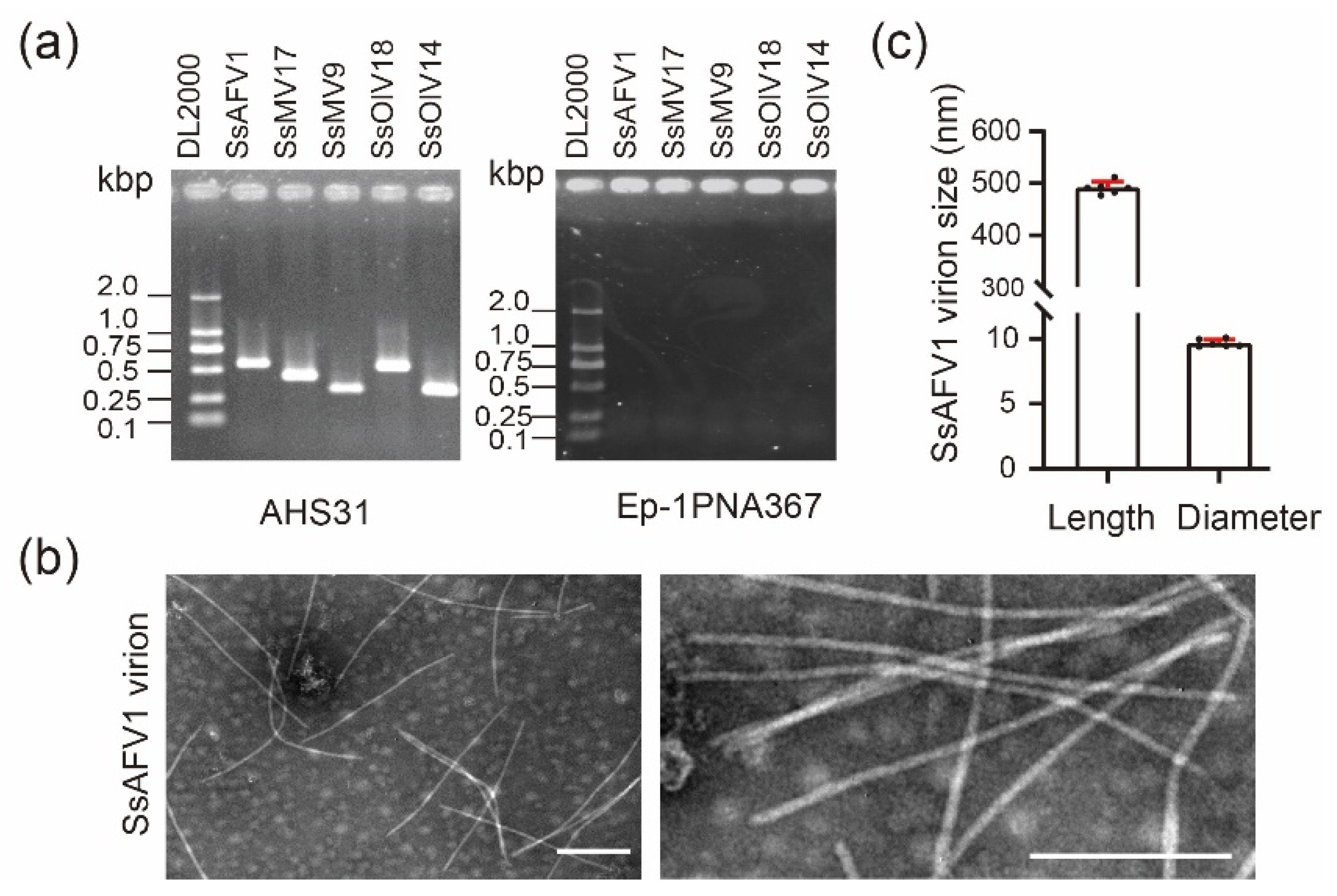
3.1. Biological Characteristics of AHS31 Strain
3.2. Virus Verification and Virion Observation
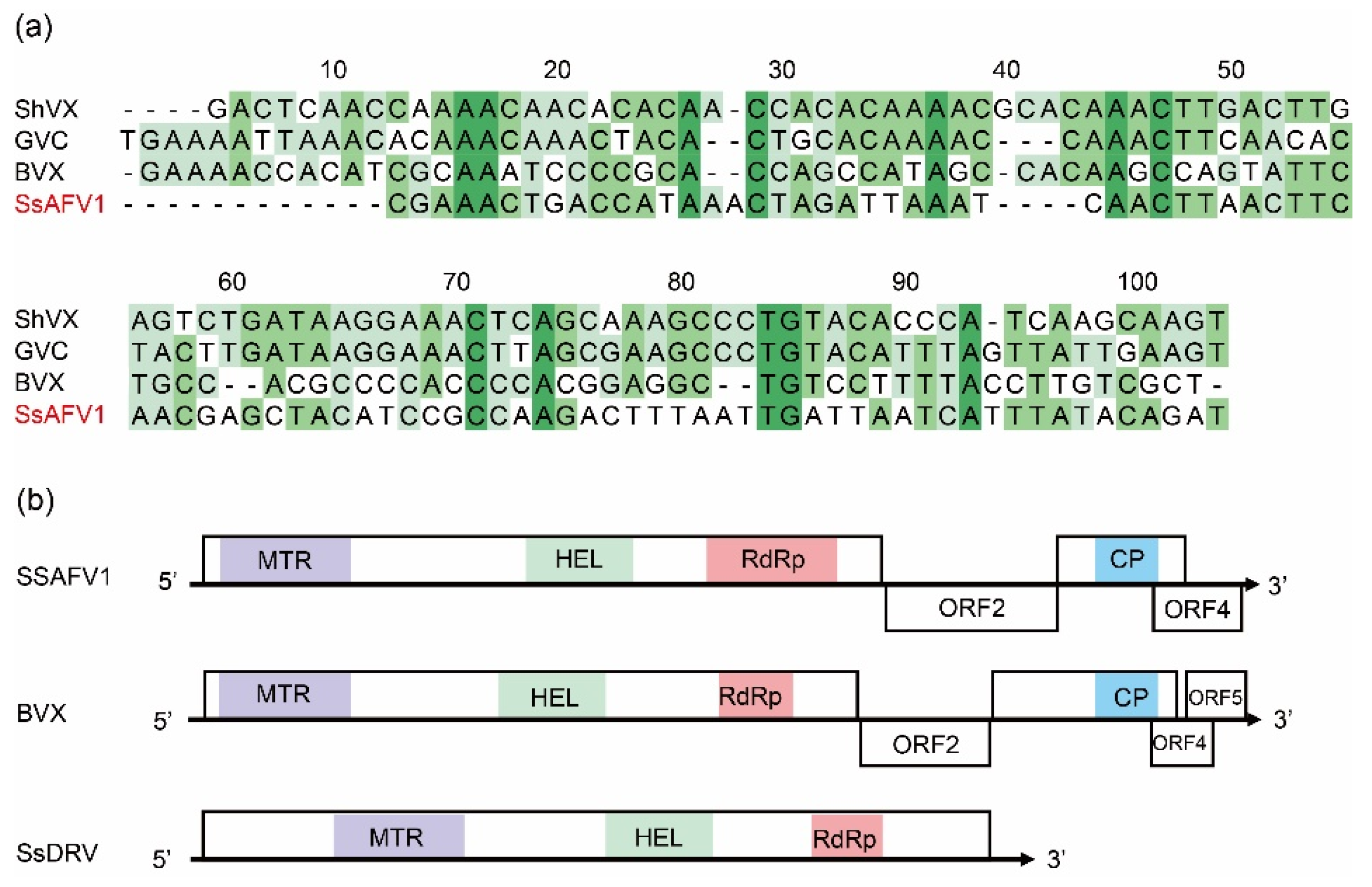
3.3. Genomic Sequence and Organization of SsAFV1
3.4. Multiple-Alignment and Phylogenetic Analysis of Predicted Proteins of SsAFV1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Derbyshire, M.; Denton-Giles, M.; Hegedus, D.; Seifbarghy, S.; Rollins, J.; van Kan, J.; Seidl, M.F.; Faino, L.; Mbengue, M.; Navaud, O.; et al. The complete genome sequence of the phytopathogenic fungus Sclerotinia sclerotiorum reveals insights into the genome architecture of broad host range pathogens. Genome Biol. Evol. 2017, 9, 593–618. [Google Scholar] [CrossRef]
- Chitrampalam, P.; Figuli, P.J.; Matheron, M.E.; Subbarao, K.V.; Pryor, B.M. Biocontrol of lettuce drop caused by Sclerotinia sclerotiorum and S. minor in desert agroecosystems. Plant Dis. 2008, 92, 1625–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peltier, A.J.; Bradley, C.A.; Chilvers, M.I.; Malvick, D.K.; Mueller, D.S.; Wise, K.A.; Esker, P.D. Biology, yield loss and control of sclerotinia stem rot of soybean. J. Integ. Pest Mngmt. 2012, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Derbyshire, M.C.; Denton-Giles, M. The control of sclerotinia stem rot on oilseed rape (Brassica napus): Current practices and future opportunities. Plant Pathol. 2016, 65, 859–877. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ullah, C.; Reichelt, M.; Beran, F.; Yang, Z.L.; Gershenzon, J.; Hammerbacher, A.; Vassao, D.G. The phytopathogenic fungus Sclerotinia sclerotiorum detoxifies plant glucosinolate hydrolysis products via an isothiocyanate hydrolase. Nat. Commun. 2020, 11, 3090. [Google Scholar] [CrossRef]
- Yang, G.; Tang, L.; Gong, Y.; Xie, J.; Fu, Y.; Jiang, D.; Li, G.; Collinge, D.B.; Chen, W.; Cheng, J. A cerato-platanin protein SsCP1 targets plant PR1 and contributes to virulence of Sclerotinia sclerotiorum. New Phytol. 2018, 217, 739–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Feng, X.; Chen, Y.; Chen, C.; Zhou, M. Occurrence and characterization of dimethachlon insensitivity in Sclerotinia sclerotiorum in Jiangsu province of China. Plant Dis. 2009, 93, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hou, Y.; Chen, C.; Zhou, M. Detection of resistance in Sclerotinia sclerotiorum to carbendazim and dimethachlon in Jiangsu Province of China. Australas. Plant Pathol. 2014, 43, 307–312. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant. Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Fu, Y.; Jiang, D.; Mu, F.; Cheng, J.; Lin, Y.; Li, B.; Marzano, S.L.; Xie, J. Interannual dynamics, diversity and evolution of the virome in Sclerotinia sclerotiorum from a single crop field. Virus Evol. 2021, 7, veab032. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.; Li, B.; Cheng, S.; Jia, J.; Jiang, D.; Fu, Y.; Cheng, J.; Lin, Y.; Chen, T.; Xie, J. Nine viruses from eight lineages exhibiting new evolutionary modes that co-infect a hypovirulent phytopathogenic fungus. PLoS Pathog. 2021, 17, e1009823. [Google Scholar] [CrossRef]
- Qu, Z.; Zhao, H.; Zhang, H.; Wang, Q.; Yao, Y.; Cheng, J.; Lin, Y.; Xie, J.; Fu, Y.; Jiang, D. Bio-priming with a hypovirulent phytopathogenic fungus enhances the connection and strength of microbial interaction network in rapeseed. NPJ Biofilms Microbiomes 2020, 6, 45. [Google Scholar] [CrossRef]
- Zhang, H.; Xie, J.; Fu, Y.; Cheng, J.; Qu, Z.; Zhao, Z.; Cheng, S.; Chen, T.; Li, B.; Wang, Q.; et al. A 2-kb mycovirus converts a pathogenic fungus into a beneficial endophyte for Brassica protection and yield enhancement. Mol. Plant 2020, 13, 1420–1433. [Google Scholar] [CrossRef]
- Xiao, X.; Cheng, J.; Tang, J.; Fu, Y.; Jiang, D.; Baker, T.S.; Ghabrial, S.A.; Xie, J. A novel partitivirus that confers hypovirulence on plant pathogenic fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Ghabrial, S.A.; Xie, J. Virus-mediated suppression of host non-self recognition facilitates horizontal transmission of heterologous viruses. PLoS Pathog. 2017, 13. [Google Scholar] [CrossRef] [Green Version]
- Morozov, S.Y.; Agranovsky, A.A. Alphaflexiviruses (Alphaflexiviridae). Encyclopedia of Virology, 4th ed.; Academic Press of Elsevier: Amsterdam, The Netherlands, 2021; pp. 140–148. [Google Scholar]
- Xie, J.; Wei, D.; Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.; Peng, Y. Characterization of debilitation-associated mycovirus infecting the plant-pathogenic fungus Sclerotinia sclerotiorum. J. Gen. Virol. 2006, 87, 241–249. [Google Scholar] [CrossRef]
- Potgieter, A.C.; Page, N.A.; Liebenberg, J.; Wright, I.M.; Landt, O.; van Dijk, A.A. Improved strategies for sequence-independent amplification and sequencing of viral double-stranded RNA genomes. J. Gen. Virol. 2009, 90, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Brown, N.P.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghabrial, S.A. Origin, adaptation and evolutionary pathways of fungal viruses. Virus Genes 1998, 16, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Janda, M.; Ahlquist, P. RNA-dependent replication, transcription, and persistence of brome mosaic virus RNA replicons in S. cerevisiae. Cell 1993, 72, 961–970. [Google Scholar] [CrossRef]
- Mascia, T.; Nigro, F.; Abdallah, A.; Ferrara, M.; De Stradis, A.; Faedda, R.; Palukaitis, P.; Gallitelli, D. Gene silencing and gene expression in phytopathogenic fungi using a plant virus vector. Proc. Natl. Acad. Sci. USA 2014, 111, 4291–4296. [Google Scholar] [CrossRef] [Green Version]
- Bian, R.; Andika, I.B.; Pang, T.; Lian, Z.; Wei, S.; Niu, E.; Wu, Y.; Kondo, H.; Liu, X.; Sun, L. Facilitative and synergistic interactions between fungal and plant viruses. Proc. Natl. Acad. Sci. USA 2020, 117, 3779–3788. [Google Scholar] [CrossRef]
- Mascia, T.; Labarile, R.; Doohan, F.; Gallitelli, D. Tobacco mosaic virus infection triggers an RNAi-based response in Phytophthora infestans. Sci. Rep. 2019, 9, 2657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andika, I.B.; Wei, S.; Cao, C.; Salaipeth, L.; Kondo, H.; Sun, L. Phytopathogenic fungus hosts a plant virus: A naturally occurring cross-kingdom viral infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Domingo, E.; Holland, J.J. RNA virus mutations and fitness for survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef]
- García-Arenal, F.; Fraile, A.; Malpica, J.M. Variability and genetic structure of plant virus populations. Annu. Rev. Phytopathol. 2001, 39, 157–186. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.L.; Roossinck, M.J. Genetic diversity in RNA virus quasispecies is controlled by host-virus interactions. J. Virol. 2001, 75, 6566–6571. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, L.; Esmael, A.; Duan, J.; Bian, X.; Jia, J.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D.; et al. Four novel botourmiaviruses co-infecting an isolate of the rice blast fungus Magnaporthe oryzae. Viruses 2020, 12, 1383. [Google Scholar] [CrossRef]
- Howitt, R.L.; Beever, R.E.; Pearson, M.N.; Forster, R.L. Genome characterization of a flexuous rod-shaped mycovirus, Botrytis virus X, reveals high amino acid identity to genes from plant ‘potex-like’ viruses. Arch. Virol. 2006, 151, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Miyanishi, M.; Ozaki, K.; Onoue, M.; Yoshida, K. Molecular characterization of a partitivirus from the plant pathogenic ascomycete Rosellinia necatrix. Arch. Virol. 2005, 150, 1069–1083. [Google Scholar] [CrossRef] [PubMed]
- Azhar, A.; Mu, F.; Huang, H.; Cheng, J.; Fu, Y.; Hamid, M.R.; Jiang, D.; Xie, J. A novel RNA virus related to sobemoviruses confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Viruses 2019, 11, 759. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Cheng, S.; Xiao, X.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Xie, J. Discovery of two mycoviruses by high-throughput sequencing and assembly of mycovirus-derived small silencing RNAs from a hypovirulent strain of Sclerotinia sclerotiorum. Front. Microbiol. 2019, 10, 1415. [Google Scholar] [CrossRef]
- Hillman, B.I.; Cai, G. The family Narnaviridae: Simplest of RNA viruses. Adv. Virus Res. 2013, 86, 149–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Mu, F.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D. A single ssRNA segment encoding RdRp is sufficient for replication, infection, and transmission of ourmia-like virus in fungi. Front. Microbiol. 2020, 11, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.M.; Mahtani, P.H.; Lee, K.C.; Yu, H.H.; Tan, Y.; Neo, K.K.; Chan, Y.; Wu, M.; Chng, C.G. Cymbidium mosaic potexvirus RNA: Complete nucleotide sequence and phylogenetic analysis. Arch. Virol. 1997, 142, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Alves, T.M.; de Novaes, Q.S.; de Paula, A.; Camelo-García, V.M.; Nagata, T.; Silva, J.M.F.; Rezende, J.A.M.; Kitajima, E.W. Near-complete genome sequence and biological properties of an allexivirus found in Senna rizzinii in Brazil. Arch. Virol. 2020, 165, 1463–1467. [Google Scholar] [CrossRef]
- Aguilar, J.M.; Hernández-Gallardo, M.D.; Cenis, J.L.; Lacasa, A.; Aranda, M.A. Complete sequence of the Pepino mosaic virus RNA genome. Arch. Virol. 2002, 147, 2009–2015. [Google Scholar] [CrossRef]
- Zuidema, D.; Linthorst, H.J.; Huisman, M.J.; Asjes, C.J.; Bol, J.F. Nucleotide sequence of narcissus mosaic virus RNA. J. Gen. Virol. 1989, 70, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Rustici, G.; Accotto, G.P.; Noris, E.; Masenga, V.; Luisoni, E.; Milne, R.G. Indian citrus ringspot virus: A proposed new species with some affinities to potex-, carla-, fovea- and allexiviruses. Arch. Virol. 2000, 145, 1895–1908. [Google Scholar] [CrossRef] [PubMed]
- Vaira, A.M.; Maroon-Lango, C.J.; Hammond, J. Molecular characterization of Lolium latent virus, proposed type member of a new genus in the family Flexiviridae. Arch. Virol. 2008, 153, 1263–1270. [Google Scholar] [CrossRef]
- Grisoni, M.; Marais, A.; Filloux, D.; Saison, A.; Faure, C.; Julian, C.; Theil, S.; Contreras, S.; Teycheney, P.Y.; Roumagnac, P.; et al. Two novel Alphaflexiviridae members revealed by deep sequencing of the Vanilla (Orchidaceae) virome. Arch. Virol. 2017, 162, 3855–3861. [Google Scholar] [CrossRef] [PubMed]
- Jelkmann, W.; Maiss, E.; Martin, R.R. The nucleotide sequence and genome organization of strawberry mild yellow edge-associated potexvirus. J. Gen. Virol. 1992, 73, 475–479. [Google Scholar] [CrossRef]
- Wylie, S.J.; Li, H.; Jones, M.G. Donkey orchid symptomless virus: A viral ’platypus’ from Australian terrestrial orchids. PLoS ONE 2013, 8, e79587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumi, S.; Matsumi, T.; Tsuneyoshi, T. Complete nucleotide sequences of garlic viruses A and C, members of the newly ratified genus Allexivirus. Arch. Virol. 1999, 144, 1819–1826. [Google Scholar] [CrossRef]
- Song, S.I.; Song, J.T.; Kim, C.H.; Lee, J.S.; Choi, Y.D. Molecular characterization of the garlic virus X genome. J. Gen. Virol. 1998, 79, 155–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Chen, J.; Adams, M.J. Molecular characterisation of a complex mixture of viruses in garlic with mosaic symptoms in China. Arch. Virol. 1999, 146, 1841–1853. [Google Scholar] [CrossRef] [PubMed]
- Kanyuka, K.V.; Vishnichenko, V.K.; Levay, K.E.; Kondrikov, D.Y.; Ryabov, E.V.; Zavriev, S.K. Nucleotide sequence of shallot virus X RNA reveals a 5′-proximal cistron closely related to those of potexviruses and a unique arrangement of the 3′-proximal cistrons. J. Gen. Virol. 1992, 73, 2553–2560. [Google Scholar] [CrossRef] [PubMed]
- Huisman, M.J.; Linthorst, H.J.; Bol, J.F.; Cornelissen, J.C. The complete nucleotide sequence of potato virus X and its homologies at the amino acid level with various plus-stranded RNA viruses. J. Gen. Virol. 1998, 69, 1789–1798. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through Metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, Z.; Hai, D.; Huang, H.; Cheng, J.; Fu, Y.; Lin, Y.; Jiang, D.; Xie, J. Mycoviromic analysis unveils complex virus composition in a hypovirulent strain of Sclerotinia sclerotiorum. J. Fungi 2022, 8, 649. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ to 3′) | Length (bp) | Description |
|---|---|---|---|
| AFV1F | ATAGGTTCGCTCAGCCTTTTG | 21 | For SsAFV1 detection |
| AFV1R | CAGCCCTCTACACCGCATT | 19 | |
| MV9F | TATCAGGATTCATACCGAGGCA | 22 | For SsMV9 detection |
| MV9R | CACCGACAAAGGAAAGAAGGAG | 22 | |
| MV17F | AGCAGAGTGGACCAGGCTATT | 21 | For SsMV17 detection |
| MV17R | TGTTCACCCTATCCCATCATTT | 22 | |
| OlV18F | TGTGACGGCTGAGAAGTTGAA | 21 | For SsOlV18 detection |
| OlV18R | TCCCATCCTCGTTGTCTGAAT | 21 | |
| OlV14F | GGAAGACGGCAGCAGCAAA | 19 | For SsOlV14 detection |
| OlV14R | TCGCCACTCCCAGAAAAGC | 19 | |
| PC2 | CCGAATTCCCGGGATCC | 17 | For nested PCR |
| 3AFV1L | CACTCAACGCTCACTTGCTC | 20 | For 3′ terminal-sequence cloning of SsAFV1 |
| 3AFV1S | GAACGACACCACCACAATGG | 20 | |
| 5AFV1L | GACCAGGGGATGTTCGCATA | 20 | For 5′ terminal sequence cloning of SsAFV1 |
| 5AFV1S | GCGTGTGAGTGTAATTGCGT | 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, T.; Lu, Z.; Li, H.; Duan, J.; Hai, D.; Lin, Y.; Xie, J.; Cheng, J.; Li, B.; Chen, T.; et al. Characterization of a Fungal Virus Representing a Novel Genus in the Family Alphaflexiviridae. Viruses 2023, 15, 339. https://doi.org/10.3390/v15020339
Ye T, Lu Z, Li H, Duan J, Hai D, Lin Y, Xie J, Cheng J, Li B, Chen T, et al. Characterization of a Fungal Virus Representing a Novel Genus in the Family Alphaflexiviridae. Viruses. 2023; 15(2):339. https://doi.org/10.3390/v15020339
Chicago/Turabian StyleYe, Ting, Zhongbo Lu, Han Li, Jie Duan, Du Hai, Yang Lin, Jiatao Xie, Jiasen Cheng, Bo Li, Tao Chen, and et al. 2023. "Characterization of a Fungal Virus Representing a Novel Genus in the Family Alphaflexiviridae" Viruses 15, no. 2: 339. https://doi.org/10.3390/v15020339
APA StyleYe, T., Lu, Z., Li, H., Duan, J., Hai, D., Lin, Y., Xie, J., Cheng, J., Li, B., Chen, T., Fu, Y., & Jiang, D. (2023). Characterization of a Fungal Virus Representing a Novel Genus in the Family Alphaflexiviridae. Viruses, 15(2), 339. https://doi.org/10.3390/v15020339