
Polymycovirus Infection Sensitizes Aspergillus fumigatus for Antifungal Effects of Nikkomycin Z


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
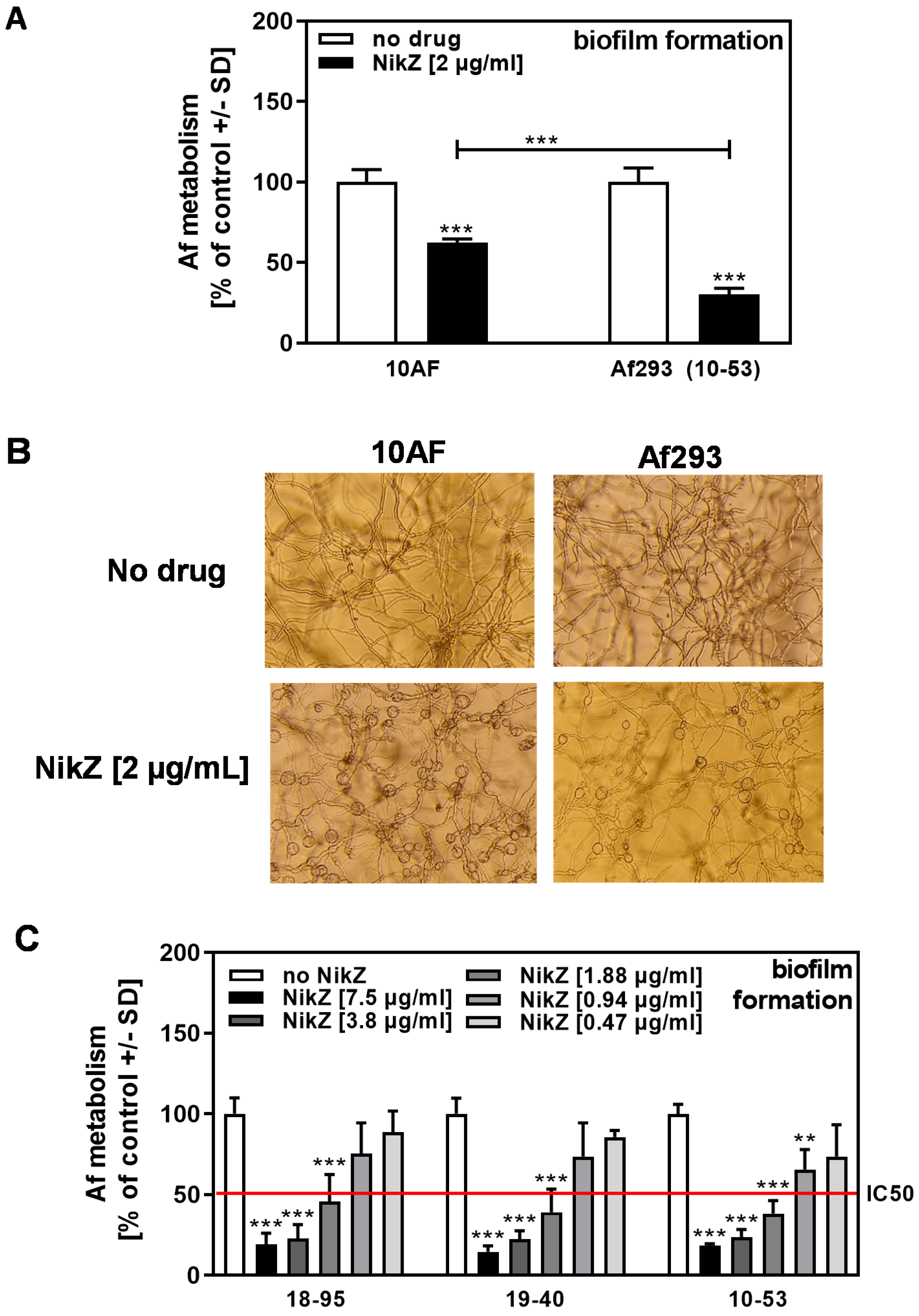
3.1. NikZ Dose-Dependently Interfered with Forming Biofilm Metabolism of Two A. fumigatus Reference Strains and Caused Structural Changes in Hyphae
3.2. Infected Af293 Is More Sensitive to NikZ Than Virus-Free Af293
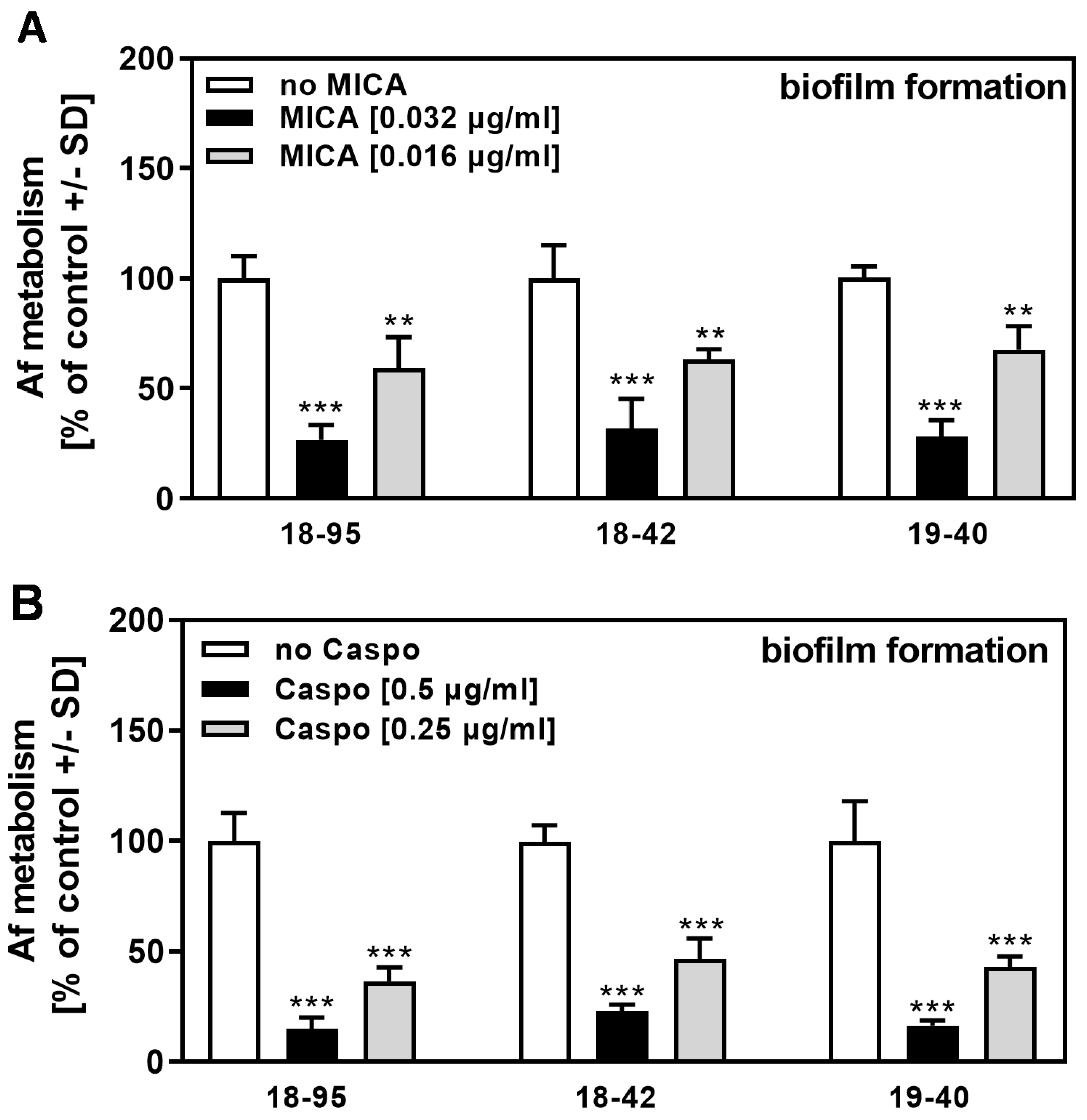
3.3. AfuPmV-1 Infection Does Not Sensitize A. fumigatus Biofilm to Conventional Drugs
3.4. Virus-Free Planktonic Af293 Is Less Sensitive to NikZ Than Infected Fungus
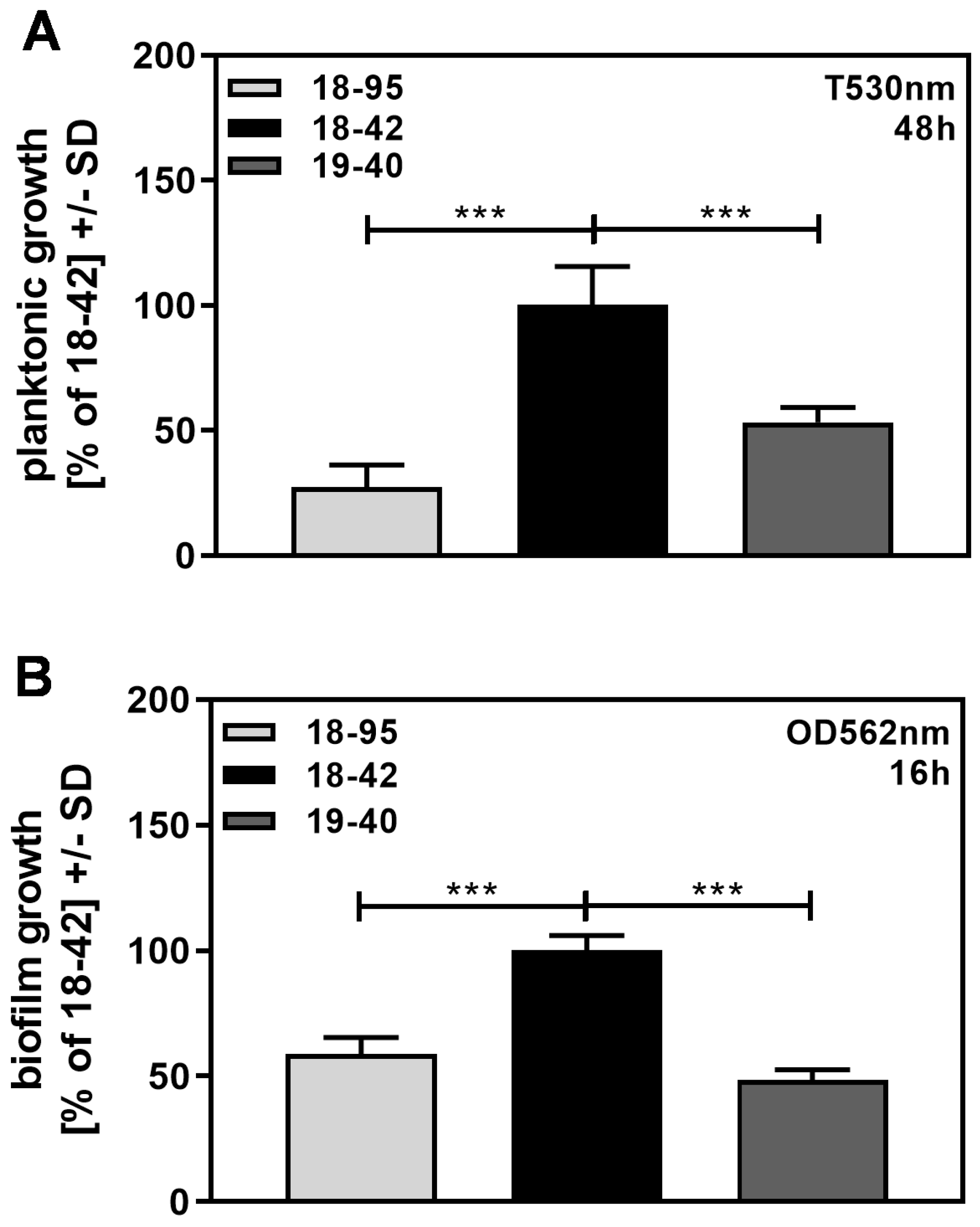
3.5. Increased Growth or Metabolism Does Not Explain Decreased Sensitivity of Virus-Free Aspergillus to NikZ
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kotta-Loizou, I.; Coutts, R.H.A. Mycoviruses in Aspergilli: A comprehensive review. Front. Microbiol. 2017, 8, 1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotta-Loizou, I. Mycoviruses and their role in fungal pathogenesis. Curr. Opin. Microbiol. 2021, 63, 10–18, Review. [Google Scholar] [CrossRef] [PubMed]
- Kanhayuwa, L.; Kotta-Loizou, I.; Özkan, S.; Gunning, A.P.; Coutts, R.H.A. A novel mycovirus from Aspergillus fumigatus contains four unique dsRNAs as its genome and is infectious as dsRNA. Proc. Natl. Acad. Sci. USA 2015, 112, 9100–9105. [Google Scholar] [CrossRef] [Green Version]
- Kotta-Loizou, I.; Coutts, R.H.A.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Polymycoviridae 2022. J. Gen. Virol. 2022, 103. [Google Scholar] [CrossRef] [PubMed]
- Kotta-Loizou, I.; Coutts, R.H.A. Studies on the virome of the entomopathogenic fungus Beauveria bassiana reveal novel dsRNA elements and mild hypervirulence. PLoS Pathog. 2017, 13, e1006183. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Shamsi, W.; Jamal, A.; Bhatti, M.F.; Kondo, H.; Suzuki, N. Hadaka virus 1: A capsidless eleven-segmented positive-sense single-stranded RNA virus from a phytopathogenic fungus, Fusarium oxysporum. MBio 2020, 11, e00450-20. [Google Scholar] [CrossRef]
- Jia, H.; Dong, K.; Zhou, L.; Wang, G.; Hong, N.; Jiang, D.; Xu, W. A dsRNA virus with filamentous viral particles. Nat. Commun. 2017, 8, 168. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Yuan, Y.; Mao, J.; Yang, Z.; Cao, Q.; Zhang, T.; Wang, S.; Liu, D. Characterization of two novel mycoviruses from Penicillium digitatum and the related fungicide resistance analysis. Sci. Rep. 2018, 8, 5513. [Google Scholar] [CrossRef] [Green Version]
- Nierman, W.C.; Pain, A.; Anderson, M.J.; Wortman, J.R.; Kim, H.S.; Arroyo, J.; Berriman, M.; Abe, K.; Archer, D.B.; Bermejo, C.; et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 2005, 438, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- Nazik, H.; Kotta-Loizou, I.; Sass, G.; Coutts, R.H.A.; Stevens, D.A. Virus infection of Aspergillus fumigatus compromises the fungus in intermicrobial competition. Viruses 2021, 13, 686. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.H.; Kotta-Loizou, I.; Palyzová, A.; Pluháček, T.; Coutts, R.H.A.; Stevens, D.A.; Havlíček, V. Freeing Aspergillus fumigatus of Polymycovirus infection renders it more resistant to competition with Pseudomonas aeruginosa due to altered iron-acquiring tactics. J. Fungi. 2021, 7, 497. [Google Scholar] [CrossRef]
- Nazik, H.; Sass, G.; Déziel, E.; Stevens, D.A. Aspergillus is inhibited by Pseudomonas aeruginosa volatiles. J. Fungi. 2020, 6, 118. [Google Scholar] [CrossRef]
- Stevens, D.A.; Kotta-Loizou, I.; Martinez, M.; Coutts, R.H.A.; Sass, G. Virus infection impairs fungal response to stress: Effect of salt. In Proceedings of the Program of the 10th Advances Against Aspergillosis and Mucormycosis Meeting, Virtual, 2–3 February 2022. Abstract No. 13. [Google Scholar]
- Denning, D.W.; Clemons, K.V.; Hanson, L.H.; Stevens, D.A.; Morrison, D.C.; Silverstein, R.; Bright, S.W.; Chen, T.-Y.; Flebbe, L.M.; Lei, M.-G. Restriction endonuclease analysis of total cellular DNA of Aspergillus fumigatus isolates of geographically and epidemiologically diverse origin. J. Infect. Dis. 1990, 162, 1151–1158. [Google Scholar] [CrossRef]
- Denning, D.W.; Stevens, D.A. Efficacy of cilofungin alone and in combination with amphotericin B in a murine model of disseminated aspergillosis. Antimicrob. Agents Chemother. 1991, 35, 1329–1333. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.A.; Penner, J.C.; Moss, R.B.; Haagensen, J.A.; Clemons, K.V.; Spormann, A.M.; Nazik, H.; Cohen, K.; Banaei, N.; Carolino, E.; et al. Inhibition of Aspergillus fumigatus and its biofilm by Pseudomonas aeruginosa is dependent on the source, phenotype and growth conditions of the bacterium. PLoS ONE 2015, 10, e0134692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scudiero, D.A.; Shoemaker, R.H.; Paull, K.D.; Monks, A.; Tierney, S.; Nofziger, T.H.; Currens, M.J.; Seniff, D.; Boyd, M.R. Evaluation of a soluble tetrazolium/formazan assay for cell growth and drug sensitivity in culture using human and other tumor cell lines. Cancer Res. 1988, 48, 4827–4833. [Google Scholar] [PubMed]
- Document M38-A2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008.
- Denning, D.W.; Hanson, L.H.; Perlman, A.M.; Stevens, D.A. In vitro susceptibility and synergy studies of Aspergillus species to conventional and new agents. Diagn. Microbiol. Infect. Dis. 1992, 15, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Kotta-Loizou, I. Diversity of mycoviruses in Aspergilli. In Encyclopedia of Virology, 4th ed.; Bamford, D.H., Mark Zuckerman, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 4, pp. 450–456. [Google Scholar]
- Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Discovery and characterization of novel Aspergillus fumigatus mycoviruses. PLoS ONE 2018, 13, e0200511. [Google Scholar] [CrossRef]
- Refos, J.M.; Vonk, A.G.; Eadie, K.; Lo-Ten-Foe, J.R.; Verbrugh, H.A.; van Diepeningen, A.D.; van de Sande, W.W.J. Doublestranded RNA mycovirus infection of Aspergillus fumigatus is not dependent on the genetic make-up of the host. PLoS ONE 2013, 8, e77381. [Google Scholar] [CrossRef] [Green Version]
- Bhatti, M.F.; Jamal, A.; Bignell, E.M.; Petrou, M.A.; Coutts, R.H.A. Incidence of dsRNA mycoviruses in a collection of Aspergillus fumigatus isolates. Mycopathologia 2012, 174, 323–326. [Google Scholar] [CrossRef]
- van de Sande, W.; Lo-Ten-Foe, J.R.; van Belkum, A.; Netea, M.G.; Kullberg, B.J.; Vonk, A.G. Mycoviruses: Future therapeutic agents of invasive fungal infections in humans? Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Takahashi-Nakaguchi, A.; Shishido, E.; Yahara, M.; Urayama, S.I.; Ninomiya, A.; Chiba, Y.; Sakai, K.; Hagiwara, D.; Chibana, H.; Moriyama, H.; et al. Phenotypic and molecular biological analysis of polymycovirus AfuPmV-1M from Aspergillus fumigatus: Reduced fungal virulence in a mouse infection model. Front. Microbiol. 2020, 11, 607795. [Google Scholar] [CrossRef]
- van de Sande, W.W.J.; Vonk, A.G. Mycovirus therapy for invasive pulmonary aspergillosis? Med. Mycol. 2019, 57, S179–S188. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, S.; Coutts, R.H.A. Aspergillus fumigatus mycovirus causes mild hypervirulent effect on pathogenicity when tested on Galleria mellonella. Fungal. Genet. Biol. 2015, 76, 20–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatti, M.F.; Jamal, A.; Petrou, M.A.; Cairns, T.C.; Bignell, E.M.; Coutts, R.H.A. The effects of dsRNA mycoviruses on growth and murine virulence of Aspergillus fumigatus. Fungal. Genet. Biol. 2011, 48, 1071–1075. [Google Scholar] [CrossRef]
- Ji, Q.; Li, B.; Chu, Y.; Wu, H.; Du, C.; Xu, Y.; Shen, Y.; Deng, J. Design, synthesis and biological evaluation of novel diazaspirodecanone derivatives containing piperidine-4-carboxamide as chitin synthase inhibitors and antifungal agents. Bioorg. Chem. 2021, 114, 105108. [Google Scholar] [CrossRef]
- Sathishkumar, Y.; Velmurugan, N.; Lee, H.M.; Rajagopal, K.; Im, C.K.; Lee, Y.S. Effect of low shear modeled microgravity on phenotypic and central chitin metabolism in the filamentous fungi Aspergillus niger and Penicillium chrysogenum. Antonie. Van Leeuwenhoek 2014, 106, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Ram, A.F.; Arentshorst, M.; Damveld, R.A.; vanKuyk, P.A.; Klis, F.M.; van den Hondel, C.A. The cell wall stress response in Aspergillus niger involves increased expression of the glutamine: Fructose-6-phosphate amidotransferase-encoding gene (gfaA) and increased deposition of chitin in the cell wall. Microbiology 2004, 150, 3315–3326. [Google Scholar] [CrossRef] [Green Version]
- Dichtl, K.; Samantaray, S.; Wagener, J. Cell wall integrity signalling in human pathogenic fungi. Cell Microbiol. 2016, 18, 1228–1238. [Google Scholar] [CrossRef]
- Rocha, M.C.; Fabri, J.H.T.M.; Simões, I.T.; Silva-Rocha, R.; Hagiwara, D.; da Cunha, A.F.; Goldman, G.H.; Cánovas, D.; Malavazi, I. The cell wall integrity pathway contributes to the early stages of Aspergillus fumigatus asexual development. Appl. Environ. Microbiol. 2020, 86, e02347-19. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | ATCC Number | CIMR Number | Origin | AfuPmV-1 Infection | Ref. |
|---|---|---|---|---|---|
| 10AF | 90240 | Virulent patient isolate | Not infected | [14,15] | |
| Af293 | MYA-4609 | 10–53 | USA | Naturally infected | [9] |
| Af293 | MYA-4609 | 18–95 | UK | Naturally infected | [9] |
| Af293 | MYA-4609 | 18–95B | UK | Naturally infected | [9] |
| Af293 | MYA-4609 | 18–95C | UK | Naturally infected | [9] |
| Af293 | 18–42 | UK | 18–95 cured | [3] | |
| Af293 | 18–42B | UK | 18–95 cured | [3] | |
| Af293 | 18–42C | UK | 18–95 cured | [3] | |
| Af293 | 19–40 | UK | 18–42 reinfected | [3] |
| Strain | MIC 50% | MIC 95% | MIC 100% | MFC |
|---|---|---|---|---|
| 18–95 | 2 | 8 | >128 | >128 |
| 18–42 | 4 | >128 | >128 | >128 |
| 19–40 | 2 | 16 | >128 | >128 |
| 10–53 | 2 | 8 | >128 | >128 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sass, G.; Kotta-Loizou, I.; Martinez, M.; Larwood, D.J.; Stevens, D.A. Polymycovirus Infection Sensitizes Aspergillus fumigatus for Antifungal Effects of Nikkomycin Z. Viruses 2023, 15, 197. https://doi.org/10.3390/v15010197
Sass G, Kotta-Loizou I, Martinez M, Larwood DJ, Stevens DA. Polymycovirus Infection Sensitizes Aspergillus fumigatus for Antifungal Effects of Nikkomycin Z. Viruses. 2023; 15(1):197. https://doi.org/10.3390/v15010197
Chicago/Turabian StyleSass, Gabriele, Ioly Kotta-Loizou, Marife Martinez, David J. Larwood, and David A. Stevens. 2023. "Polymycovirus Infection Sensitizes Aspergillus fumigatus for Antifungal Effects of Nikkomycin Z" Viruses 15, no. 1: 197. https://doi.org/10.3390/v15010197
APA StyleSass, G., Kotta-Loizou, I., Martinez, M., Larwood, D. J., & Stevens, D. A. (2023). Polymycovirus Infection Sensitizes Aspergillus fumigatus for Antifungal Effects of Nikkomycin Z. Viruses, 15(1), 197. https://doi.org/10.3390/v15010197