

Host Desmin Interacts with RABV Matrix Protein and Facilitates Virus Propagation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Plasmids, and Virus
2.2. GST-Pulldown Assay
2.3. Immunoprecipitation Assay
2.4. Transfection/Infection Assays
2.5. siRNA Knockdown
2.6. Quantitative RT-PCR
2.7. Indirect Immunofluorescence Assay
2.8. In Vivo Studies
2.9. Immunohistochemistry
2.10. Statistical Analysis
3. Results
3.1. Identification of Desmin as a Host Interactor with RABV Matrix Protein
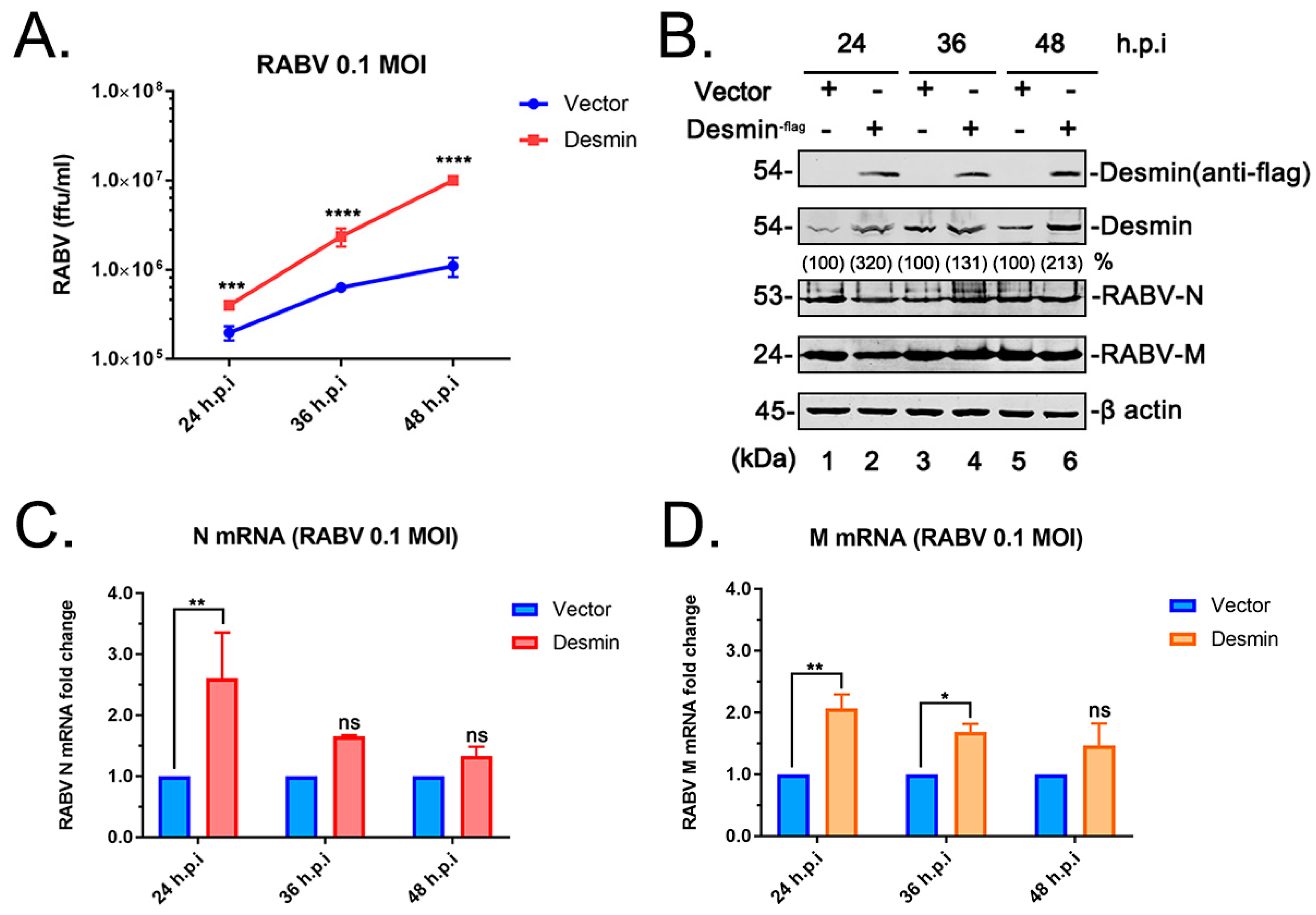
3.2. Desmin Facilitates RABV Infection
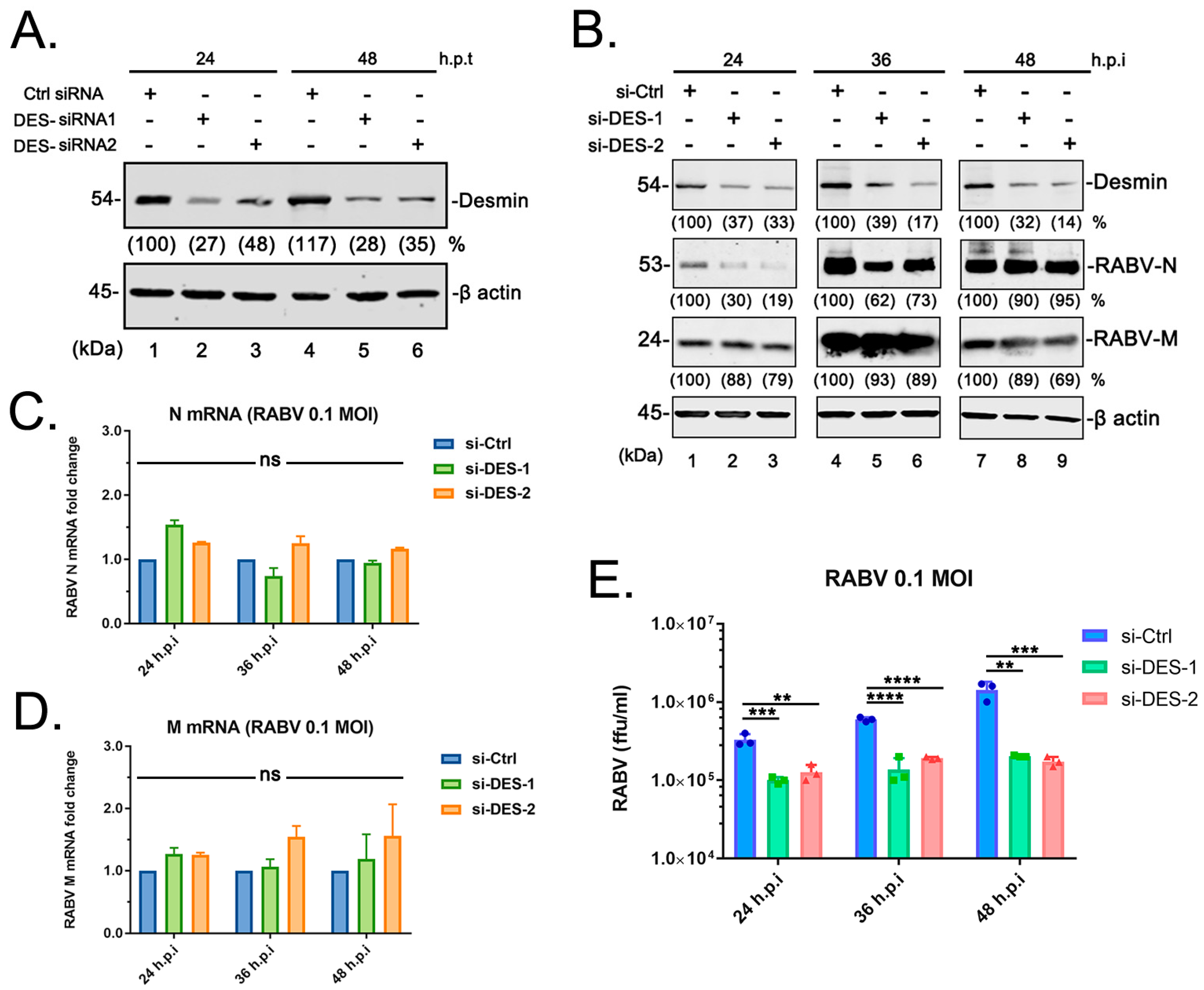
3.3. Suppression of Endogenous Desmin Impairs RABV Infection
3.4. RABV Matrix Protein Localizes to the Desmin-Formed Intermediate Filament Bundles
3.5. RABV Infection Enhances Desmin Expression in Mouse Brain
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, C.R.; Streicker, D.G.; Schnell, M.J. The spread and evolution of rabies virus: Conquering new frontiers. Nat. Rev. Microbiol. 2018, 16, 241–255. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Expert Consultation on Rabies: Third Report; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Smith, T.G.; Wu, X.; Franka, R.; Rupprecht, C.E. Design of future rabies biologics and antiviral drugs. Adv. Virus Res. 2011, 79, 345–363. [Google Scholar]
- Du Pont, V.; Plemper, R.K.; Schnell, M.J. Status of antiviral therapeutics against rabies virus and related emerging lyssaviruses. Curr. Opin. Virol. 2019, 35, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Okumura, A.; Harty, R.N. Rabies virus assembly and budding. Adv. Virus Res. 2011, 79, 23–32. [Google Scholar]
- Harty, R.N.; Paragas, J.; Sudol, M.; Palese, P. A proline-rich motif within the matrix protein of vesicular stomatitis virus and rabies virus interacts with WW domains of cellular proteins: Implications for viral budding. J. Virol. 1999, 73, 2921–2929. [Google Scholar] [CrossRef]
- Wirblich, C.; Tan, G.S.; Papaneri, A.; Godlewski, P.J.; Orenstein, J.M.; Harty, R.N.; Schnell, M.J. PPEY motif within the rabies virus (RV) matrix protein is essential for efficient virion release and RV pathogenicity. J. Virol. 2008, 82, 9730–9738. [Google Scholar] [CrossRef]
- Komarova, A.V.; Real, E.; Borman, A.M.; Brocard, M.; England, P.; Tordo, N.; Hershey, J.W.; Kean, K.M.; Jacob, Y. Rabies virus matrix protein interplay with eIF3, new insights into rabies virus pathogenesis. Nucleic Acids Res. 2007, 35, 1522–1532. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, Y.; Zhang, Q.; Wu, Y.; Zhang, B.; Mo, M.; Tian, Q.; Zhao, J.; Mei, M.; Guo, X. The Deoptimization of Rabies Virus Matrix Protein Impacts Viral Transcription and Replication. Viruses 2019, 12, 4. [Google Scholar] [CrossRef]
- Liu, X.; Li, F.; Zhang, J.; Wang, L.; Wang, J.; Wen, Z.; Wang, Z.; Shuai, L.; Wang, X.; Ge, J.; et al. The ATPase ATP6V1A facilitates rabies virus replication by promoting virion uncoating and interacting with the viral matrix protein. J. Biol. Chem. 2021, 296, 100096. [Google Scholar] [CrossRef]
- Ben Khalifa, Y.; Luco, S.; Besson, B.; Sonthonnax, F.; Archambaud, M.; Grimes, J.M.; Larrous, F.; Bourhy, H. The matrix protein of rabies virus binds to RelAp43 to modulate NF-kappaB-dependent gene expression related to innate immunity. Sci. Rep. 2016, 6, 39420. [Google Scholar] [CrossRef] [PubMed]
- Besson, B.; Sonthonnax, F.; Duchateau, M.; Ben Khalifa, Y.; Larrous, F.; Eun, H.; Hourdel, V.; Matondo, M.; Chamot-Rooke, J.; Grailhe, R.; et al. Regulation of NF-kappaB by the p105-ABIN2-TPL2 complex and RelAp43 during rabies virus infection. PLoS Pathog. 2017, 13, e1006697. [Google Scholar] [CrossRef] [PubMed]
- Sonthonnax, F.; Besson, B.; Bonnaud, E.; Jouvion, G.; Merino, D.; Larrous, F.; Bourhy, H. Lyssavirus matrix protein cooperates with phosphoprotein to modulate the Jak-Stat pathway. Sci. Rep. 2019, 9, 12171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zan, J.; Liu, J.; Zhou, J.W.; Wang, H.L.; Mo, K.K.; Yan, Y.; Xu, Y.B.; Liao, M.; Su, S.; Hu, R.L.; et al. Rabies virus matrix protein induces apoptosis by targeting mitochondria. Exp. Cell Res. 2016, 347, 83–94. [Google Scholar] [CrossRef]
- Kojima, I.; Onomoto, K.; Zuo, W.; Ozawa, M.; Okuya, K.; Naitou, K.; Izumi, F.; Okajima, M.; Fujiwara, T.; Ito, N.; et al. The Amino Acid at Position 95 in the Matrix Protein of Rabies Virus Is Involved in Antiviral Stress Granule Formation in Infected Cells. J. Virol. 2022, 96, e0081022. [Google Scholar] [CrossRef] [PubMed]
- Kojima, I.; Izumi, F.; Ozawa, M.; Fujimoto, Y.; Okajima, M.; Ito, N.; Sugiyama, M.; Masatani, T. Analyses of cell death mechanisms related to amino acid substitution at position 95 in the rabies virus matrix protein. J. Gen. Virol. 2021, 102. [Google Scholar] [CrossRef]
- Snider, N.T.; Omary, M.B. Post-translational modifications of intermediate filament proteins: Mechanisms and functions. Nat. Rev. Mol. Cell. Biol. 2014, 15, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Duan, M.; Wang, X.; Gao, J.; Guan, Z.; Zhang, M. Early events in rabies virus infection-Attachment, entry, and intracellular trafficking. Virus Res. 2019, 263, 217–225. [Google Scholar] [CrossRef]
- Zandi, F.; Khalaj, V.; Goshadrou, F.; Meyfour, A.; Gholami, A.; Enayati, S.; Mehranfar, M.; Rahmati, S.; Kheiri, E.V.; Badie, H.G.; et al. Rabies virus matrix protein targets host actin cytoskeleton: A protein-protein interaction analysis. Pathog. Dis. 2021, 79, ftaa075. [Google Scholar] [CrossRef]
- Liu, X.; Nawaz, Z.; Guo, C.; Ali, S.; Naeem, M.A.; Jamil, T.; Ahmad, W.; Siddiq, M.U.; Ahmed, S.; Asif Idrees, M.; et al. Rabies Virus Exploits Cytoskeleton Network to Cause Early Disease Progression and Cellular Dysfunction. Front Vet. Sci. 2022, 9, 889873. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.B.; Lu, Z.L.; Wei, X.K.; Zhong, T.Z.; Zhong, Y.Z.; Ouyang, L.X.; Luo, Y.; Xing, X.W.; Liao, F.; Peng, K.K.; et al. Viperin inhibits rabies virus replication via reduced cholesterol and sphingomyelin and is regulated upstream by TLR4. Sci. Rep. 2016, 6, 30529. [Google Scholar] [CrossRef]
- Tang, H.B.; Pan, Y.; Wei, X.K.; Lu, Z.L.; Lu, W.; Yang, J.; He, X.X.; Xie, L.J.; Zeng, L.; Zheng, L.F.; et al. Re-emergence of rabies in the Guangxi province of Southern China. PLoS Negl. Trop. Dis. 2014, 8, e3114. [Google Scholar] [CrossRef]
- Frank, E.D.; Tuszynski, G.P.; Warren, L. Localization of vimentin and desmin in BHK21/C13 cells and in baby hamster kidney. Exp. Cell Res. 1982, 139, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Pollin, R.; Granzow, H.; Kollner, B.; Conzelmann, K.K.; Finke, S. Membrane and inclusion body targeting of lyssavirus matrix proteins. Cell. Microbiol. 2013, 15, 200–212. [Google Scholar] [CrossRef]
- Zhao, J.; Zeng, Z.; Chen, Y.; Liu, W.; Chen, H.; Fu, Z.F.; Zhao, L.; Zhou, M. Lipid Droplets Are Beneficial for Rabies Virus Replication by Facilitating Viral Budding. J. Virol. 2022, 96, e0147321. [Google Scholar] [CrossRef] [PubMed]
- Lafon, M. Evasive strategies in rabies virus infection. Adv. Virus Res. 2011, 79, 33–53. [Google Scholar] [PubMed]
- Feige, L.; Zaeck, L.M.; Sehl-Ewert, J.; Finke, S.; Bourhy, H. Innate Immune Signaling and Role of Glial Cells in Herpes Simplex Virus- and Rabies Virus-Induced Encephalitis. Viruses 2021, 13, 2364. [Google Scholar] [CrossRef]
- Schnell, M.J.; McGettigan, J.P.; Wirblich, C.; Papaneri, A. The cell biology of rabies virus: Using stealth to reach the brain. Nat. Rev. Microbiol. 2010, 8, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Gao, J.; Zhang, M.; Zhang, D.; Duan, M.; Guan, Z.; Guo, Y. Dynein- and kinesin- mediated intracellular transport on microtubules facilitates RABV infection. Vet. Microbiol. 2021, 262, 109241. [Google Scholar] [CrossRef]
- Bauer, A.; Nolden, T.; Nemitz, S.; Perlson, E.; Finke, S. A Dynein Light Chain 1 Binding Motif in Rabies Virus Polymerase L Protein Plays a Role in Microtubule Reorganization and Viral Primary Transcription. J. Virol. 2015, 89, 9591–9600. [Google Scholar] [CrossRef]
- Zan, J.; Liu, S.; Sun, D.N.; Mo, K.K.; Yan, Y.; Liu, J.; Hu, B.L.; Gu, J.Y.; Liao, M.; Zhou, J.Y. Rabies Virus Infection Induces Microtubule Depolymerization to Facilitate Viral RNA Synthesis by Upregulating HDAC6. Front. Cell. Infect. Microbiol. 2017, 7, 146. [Google Scholar] [CrossRef]
- Zan, J.; An, S.T.; Mo, K.K.; Zhou, J.W.; Liu, J.; Wang, H.L.; Yan, Y.; Liao, M.; Zhou, J.Y. Rabies virus inactivates cofilin to facilitate viral budding and release. Biochem. Biophys. Res. Commun. 2016, 477, 1045–1050. [Google Scholar] [CrossRef]
- Milner, D.J.; Weitzer, G.; Tran, D.; Bradley, A.; Capetanaki, Y. Disruption of muscle architecture and myocardial degeneration in mice lacking desmin. J. Cell Biol. 1996, 134, 1255–1270. [Google Scholar] [CrossRef] [Green Version]
- Paulin, D.; Li, Z. Desmin: A major intermediate filament protein essential for the structural integrity and function of muscle. Exp. Cell. Res. 2004, 301, 1–7. [Google Scholar] [CrossRef]
- Joanne, P.; Hovhannisyan, Y.; Bencze, M.; Daher, M.T.; Parlakian, A.; Toutirais, G.; Gao-Li, J.; Lilienbaum, A.; Li, Z.; Kordeli, E.; et al. Absence of Desmin Results in Impaired Adaptive Response to Mechanical Overloading of Skeletal Muscle. Front. Cell Dev. Biol. 2021, 9, 662133. [Google Scholar] [CrossRef]
- Goebel, H.H. Desmin-related neuromuscular disorders. Muscle Nerve 1995, 18, 1306–1320. [Google Scholar] [CrossRef]
- Eiber, N.; Fröb, F.; Schowalter, M.; Thiel, C.; Clemen, C.S.; Schröder, R.; Hashemolhosseini, S. Lack of Desmin in Mice Causes Structural and Functional Disorders of Neuromuscular Junctions. Front. Mol. Neurosci. 2020, 13, 567084. [Google Scholar] [CrossRef]
- Lewis, P.; Lentz, T.L. Rabies virus entry into cultured rat hippocampal neurons. J. Neurocytol. 1998, 27, 559–573. [Google Scholar] [CrossRef]
- Stein, L.T.; Rech, R.R.; Harrison, L.; Brown, C.C. Immunohistochemical study of rabies virus within the central nervous system of domestic and wildlife species. Vet. Pathol. 2010, 47, 630–633. [Google Scholar] [CrossRef]
- Chung, B.-M.; Rotty, J.D.; Coulombe, P.A. Networking galore: Intermediate filaments and cell migration. Curr. Opin. Cell Biol. 2013, 25, 600–612. [Google Scholar] [CrossRef]
- Margiotta, A.; Bucci, C. Role of Intermediate Filaments in Vesicular Traffic. Cells 2016, 5, 20. [Google Scholar] [CrossRef]
- Etienne-Manneville, S. Cytoplasmic Intermediate Filaments in Cell Biology. Annu. Rev. Cell Dev. Biol. 2018, 34, 1–28. [Google Scholar] [CrossRef]
- Geisler, F.; Leube, R.E. Epithelial Intermediate Filaments: Guardians against Microbial Infection? Cells 2016, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Hamirally, S.; Kamil, J.P.; Ndassa-Colday, Y.M.; Lin, A.J.; Jahng, W.J.; Baek, M.C.; Noton, S.; Silva, L.A.; Simpson-Holley, M.; Knipe, D.M.; et al. Viral mimicry of Cdc2/cyclin-dependent kinase 1 mediates disruption of nuclear lamina during human cytomegalovirus nuclear egress. PLoS Pathog. 2009, 5, e1000275. [Google Scholar] [CrossRef]
- Milbradt, J.; Webel, R.; Auerochs, S.; Sticht, H.; Marschall, M. Novel mode of phosphorylation-triggered reorganization of the nuclear lamina during nuclear egress of human cytomegalovirus. J. Biol. Chem. 2010, 285, 13979–13989. [Google Scholar] [CrossRef]
- Cortese, M.; Lee, J.Y.; Cerikan, B.; Neufeldt, C.J.; Oorschot, V.M.J.; Köhrer, S.; Hennies, J.; Schieber, N.L.; Ronchi, P.; Mizzon, G.; et al. Integrative Imaging Reveals SARS-CoV-2-Induced Reshaping of Subcellular Morphologies. Cell. Host Microbe 2020, 28, 853–866.e5. [Google Scholar] [CrossRef]
- Patteson, A.E.; Vahabikashi, A.; Goldman, R.D.; Janmey, P.A. Mechanical and Non-Mechanical Functions of Filamentous and Non-Filamentous Vimentin. BioEssays News Rev. Mol. Cell. Dev. Biol. 2020, 42, e2000078. [Google Scholar] [CrossRef]
- Amraei, R.; Xia, C.; Olejnik, J.; White, M.R.; Napoleon, M.A.; Lotfollahzadeh, S.; Hauser, B.M.; Schmidt, A.G.; Chitalia, V.; Mühlberger, E.; et al. Extracellular vimentin is an attachment factor that facilitates SARS-CoV-2 entry into human endothelial cells. Proc. Natl. Acad. Sci. USA 2022, 119, e2113874119. [Google Scholar] [CrossRef]
- Nédellec, P.; Vicart, P.; Laurent-Winter, C.; Martinat, C.; Prévost, M.C.; Brahic, M. Interaction of Theiler’s virus with intermediate filaments of infected cells. J. Virol. 1998, 72, 9553–9560. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Liu, Y.; Li, M.; Zhu, J.; Li, X.; Luo, T.R.; Liang, J. Host Desmin Interacts with RABV Matrix Protein and Facilitates Virus Propagation. Viruses 2023, 15, 434. https://doi.org/10.3390/v15020434
Zhang W, Liu Y, Li M, Zhu J, Li X, Luo TR, Liang J. Host Desmin Interacts with RABV Matrix Protein and Facilitates Virus Propagation. Viruses. 2023; 15(2):434. https://doi.org/10.3390/v15020434
Chicago/Turabian StyleZhang, Wen, Yuming Liu, Mengru Li, Jian Zhu, Xiaoning Li, Ting Rong Luo, and Jingjing Liang. 2023. "Host Desmin Interacts with RABV Matrix Protein and Facilitates Virus Propagation" Viruses 15, no. 2: 434. https://doi.org/10.3390/v15020434
APA StyleZhang, W., Liu, Y., Li, M., Zhu, J., Li, X., Luo, T. R., & Liang, J. (2023). Host Desmin Interacts with RABV Matrix Protein and Facilitates Virus Propagation. Viruses, 15(2), 434. https://doi.org/10.3390/v15020434