
Dynamic of Mayaro Virus Transmission in Aedes aegypti, Culex quinquefasciatus Mosquitoes, and a Mice Model
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viral Strain
2.2. Mosquitoes and Animals
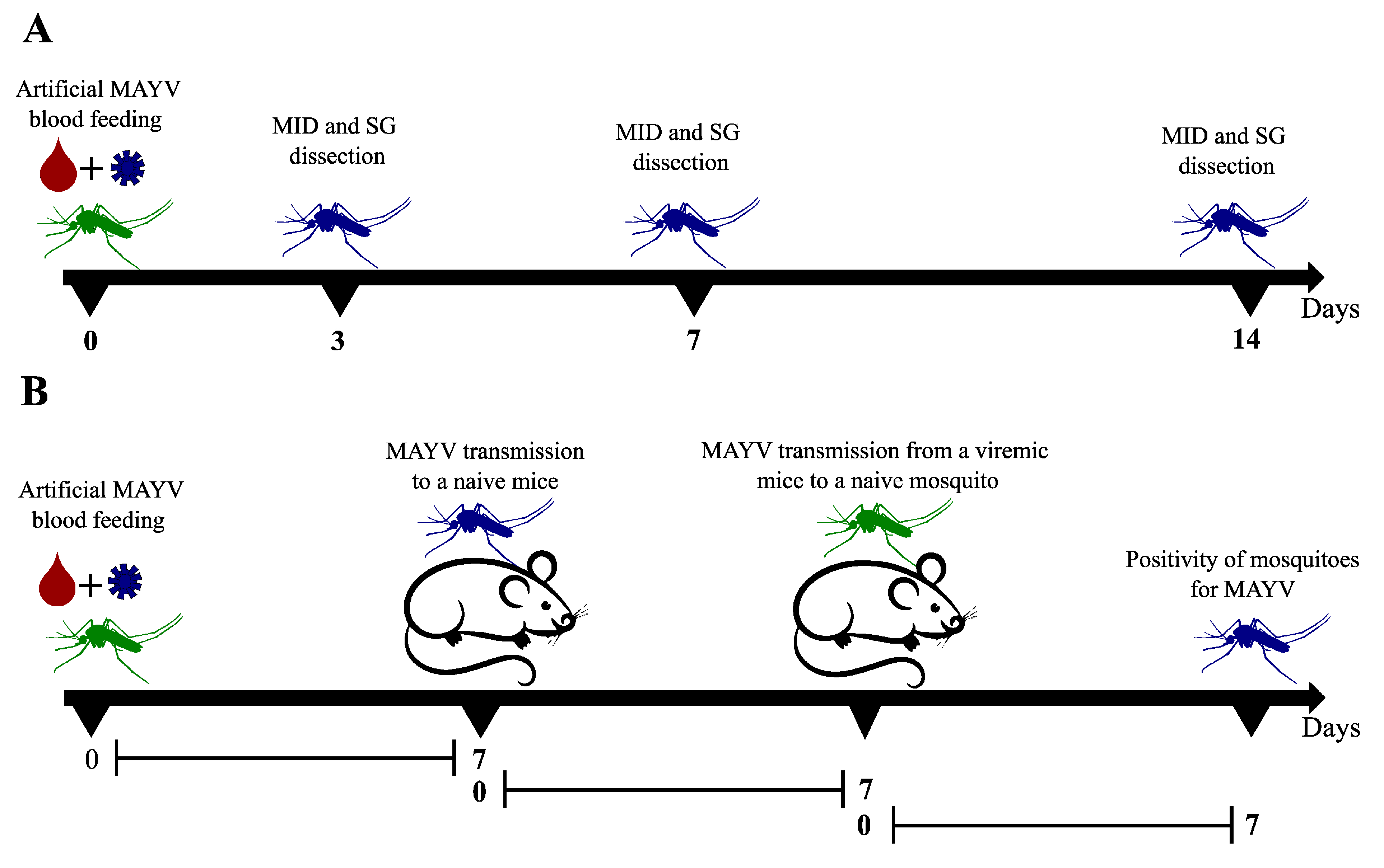
2.3. Mosquitoes Artificial Feeding and Vector Competence Evaluation
2.4. MAYV Transmission Cycle Model in Mice
2.5. RNA Isolation and RT-qPCR
2.6. Plaque Assay
2.7. Data Analysis
2.8. Ethic Statement
3. Results
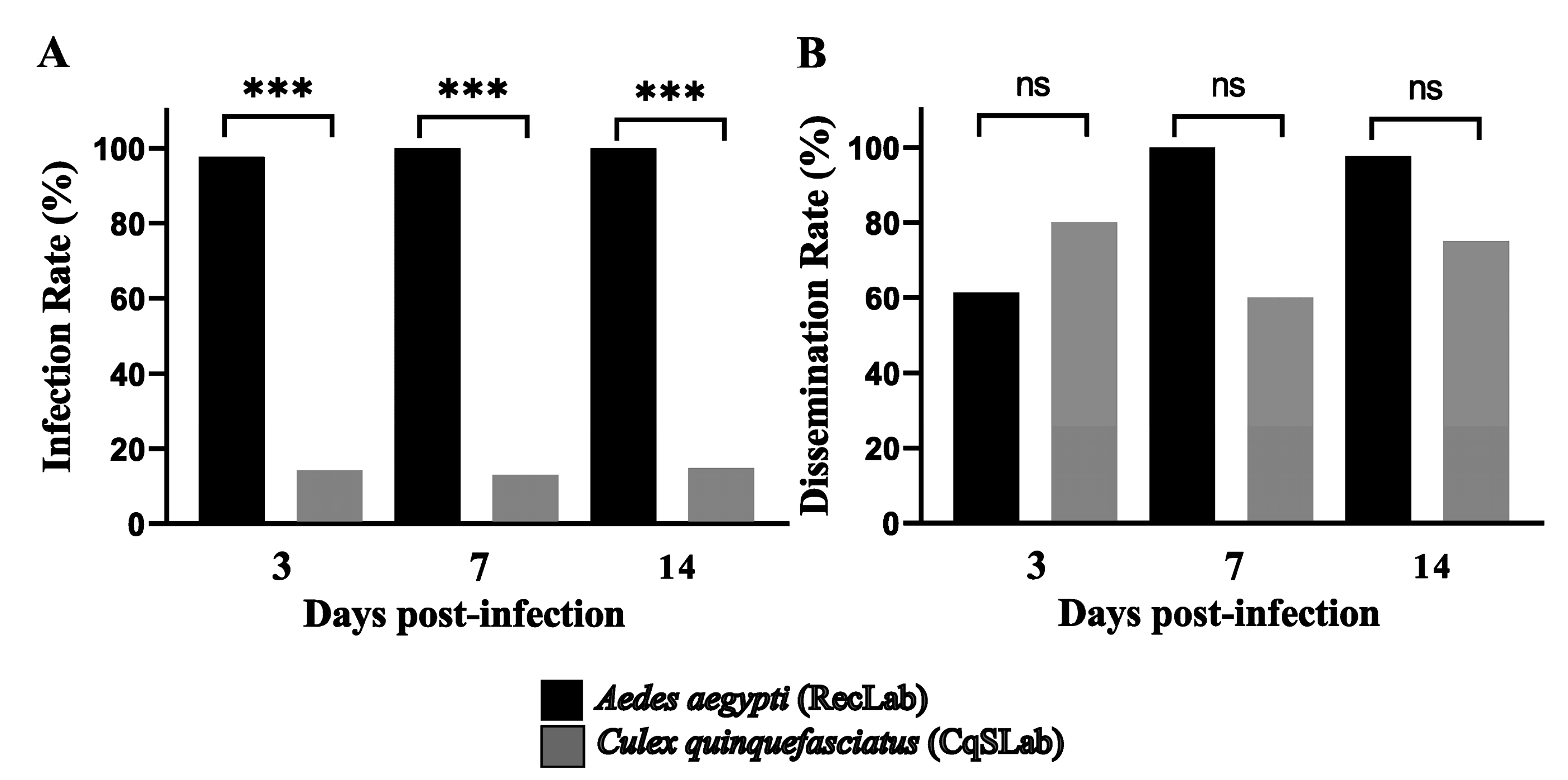
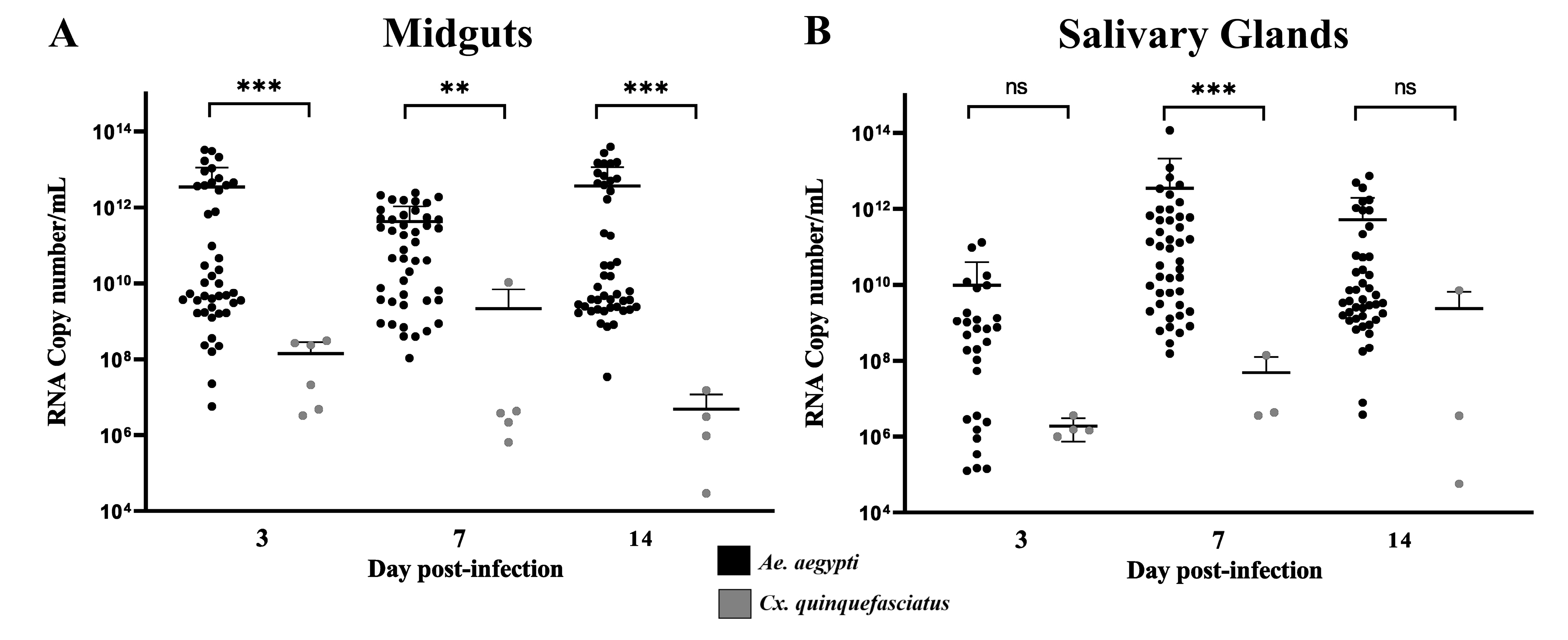
3.1. Vector Competence Assays
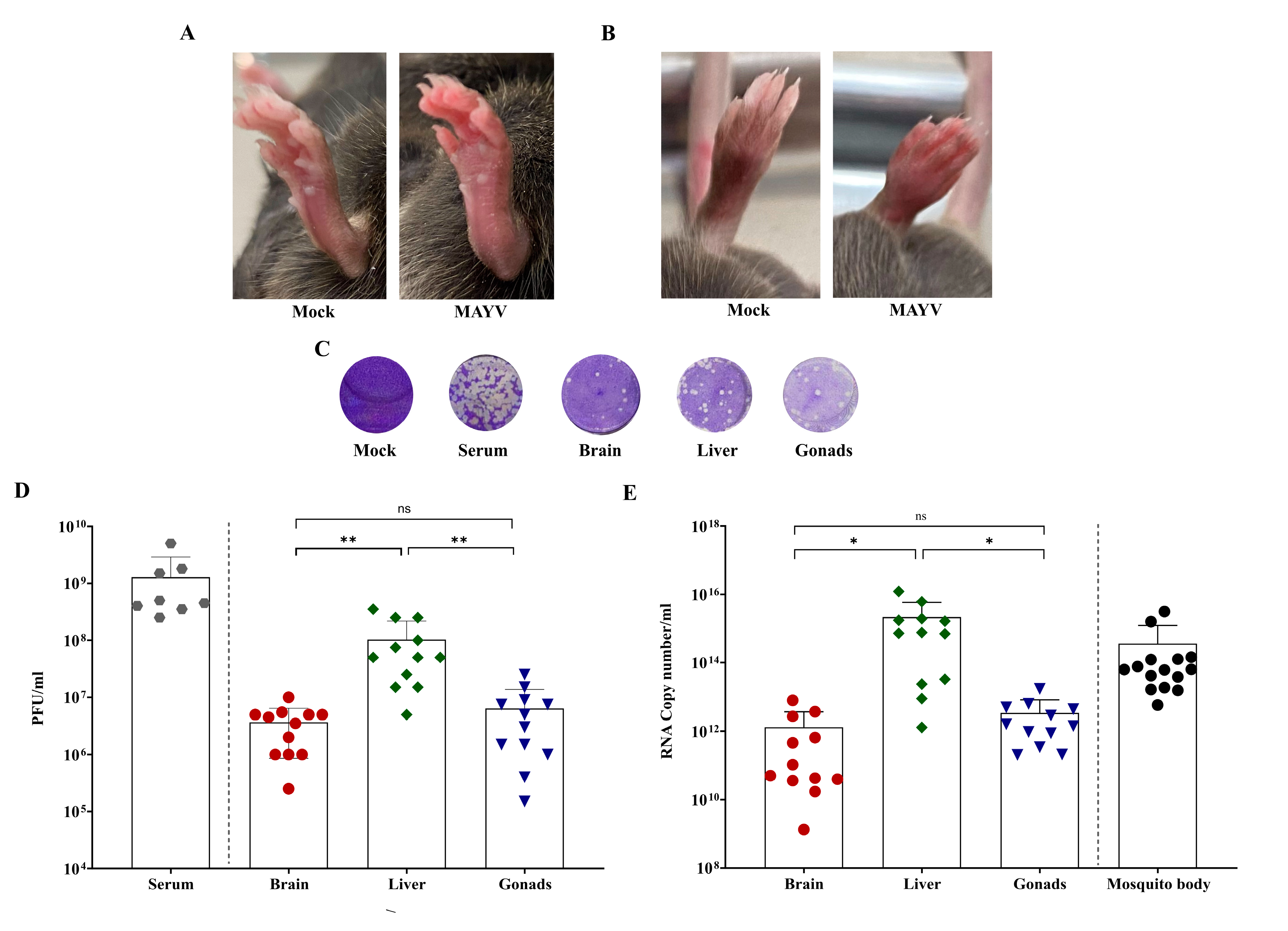
3.2. Mayaro Virus Transmission Cycle in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, C.R.; Downs, W.G.; Wattley, G.H.; Ahin, N.W.; Reese, A.A. Mayaro Virus: A New Human Disease Agent. II. Isolation from Blood of Patients in Trinidad, B.W.I. Am. J. Trop. Med. Hyg. 1957, 6, 1012–1016. [Google Scholar] [CrossRef]
- Schaeffer, M.; Gajdusek, D.C.; Lema, A.B.; Eichenwald, H. Epidemic Jungle Fevers Among Okinawan Colonists in the Bolivian Rain Forest: I. Epidemiology. Am. J. Trop. Med. Hyg. 1959, 8, 372–396. [Google Scholar] [CrossRef]
- Carvalho, V.L.; Azevedo, R.S.S.; Carvalho, V.L.; Azevedo, R.S.; Henriques, D.F.; Cruz, A.C.R.; Vasconcelos, P.F.C.; Martins, L.C. Arbovirus Outbreak in a Rural Region of the Brazilian Amazon. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2022, 150–151, 105155. [Google Scholar] [CrossRef]
- Mutricy, R.; Matheus, S.; Mosnier, É.; Martinez-Lorenzi, E.; De Laval, F.; Nacher, M.; Niemetzky, F.; Naudion, P.; Djossou, F.; Rousset, D.; et al. Mayaro Virus Infection in French Guiana, a Cross Sectional Study 2003-2019. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2022, 99, 105243. [Google Scholar] [CrossRef]
- Julia da Silva Pessoa Vieira, C.; José Ferreira da Silva, D.; Rigotti Kubiszeski, J.; Ceschini Machado, L.; Pena, L.J.; Vieira de Morais Bronzoni, R.; da Luz Wallau, G. The Emergence of Chikungunya ECSA Lineage in a Mayaro Endemic Region on the Southern Border of the Amazon Forest. Trop. Med. Infect. Dis. 2020, 5, 105. [Google Scholar] [CrossRef]
- Lima, W.G.; Pereira, R.S.; da Cruz Nizer, W.S.; Brito, J.C.M.; Godói, I.P.; Cardoso, V.N.; Fernandes, S.O.A.; Ferreira, J.M.S. Rate of Exposure to Mayaro Virus (MAYV) in Brazil between 1955 and 2018: A Systematic Review and Meta-Analysis. Arch. Virol. 2021, 166, 347–361. [Google Scholar] [CrossRef]
- Aguilar-Luis, M.A.; del Valle-Mendoza, J.; Sandoval, I.; Silva-Caso, W.; Mazulis, F.; Carrillo-Ng, H.; Tarazona-Castro, Y.; Martins-Luna, J.; Aquino-Ortega, R.; Peña-Tuesta, I.; et al. A Silent Public Health Threat: Emergence of Mayaro Virus and Co-Infection with Dengue in Peru. BMC Res. Notes 2021, 14, 29. [Google Scholar] [CrossRef]
- Mourão, M.P.G.; Bastos, M.d.S.; de Figueiredo, R.P.; Gimaque, J.B.L.; dos Santos Galusso, E.; Kramer, V.M.; de Oliveira, C.M.C.; Naveca, F.G.; Figueiredo, L.T.M. Mayaro Fever in the City of Manaus, Brazil, 2007–2008. Vector Borne Zoonotic Dis. 2012, 12, 42–46. [Google Scholar] [CrossRef]
- Arenívar, C.; Rodríguez, Y.; Rodríguez-Morales, A.J.; Anaya, J.-M. Osteoarticular Manifestations of Mayaro Virus Infection. Curr. Opin. Rheumatol. 2019, 31, 512–516. [Google Scholar] [CrossRef]
- Diagne, C.T.; Bengue, M.; Choumet, V.; Hamel, R.; Pompon, J.; Missé, D. Mayaro Virus Pathogenesis and Transmission Mechanisms. Pathogens 2020, 9, 738. [Google Scholar] [CrossRef]
- Secretaria de Vigilância em Saúde. Boletim de Vigilância Laboratorial Dos Arbovírus. Bol. Epidemiol. 2021, 52, 18. [Google Scholar]
- Celone, M.; Okech, B.; Han, B.A.; Forshey, B.M.; Anyamba, A.; Dunford, J.; Rutherford, G.; Mita-Mendoza, N.K.; Estallo, E.L.; Khouri, R.; et al. A Systematic Review and Meta-Analysis of the Potential Non-Human Animal Reservoirs and Arthropod Vectors of the Mayaro Virus. PLoS Negl. Trop. Dis. 2021, 15, e0010016. [Google Scholar] [CrossRef]
- Galindo, P.; Srihongse, S. Transmission of Arboviruses to Hamsters by the Bite of Naturally Infected Culex (Melanoconion) Mosquitoes. Am. J. Trop. Med. Hyg. 1967, 16, 525–530. [Google Scholar] [CrossRef]
- Pauvolid-Corrêa, A.; Juliano, R.S.; Campos, Z.; Velez, J.; Nogueira, R.M.R.; Komar, N.; Pauvolid-Corrêa, A.; Juliano, R.S.; Campos, Z.; Velez, J.; et al. Neutralising Antibodies for Mayaro Virus in Pantanal, Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, P.F.C.; Travassos da Rosa, A.P.A.; Rodrigues, S.G.; Travassos da Rosa, E.S.; Dégallier, N.; Travassos da Rosa, J.F.S. Inadequate Management of Natural Ecosystem in the Brazilian Amazon Region Results in the Emergence and Reemergence of Arboviruses. Cad. Saúde Pública 2001, 17, S155–S164. [Google Scholar] [CrossRef]
- Diop, F.; Alout, H.; Diagne, C.T.; Bengue, M.; Baronti, C.; Hamel, R.; Talignani, L.; Liegeois, F.; Pompon, J.; Vargas, R.E.M.; et al. Differential Susceptibility and Innate Immune Response of Aedes Aegypti and Aedes Albopictus to the Haitian Strain of the Mayaro Virus. Viruses 2019, 11, 924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, T.N.; Carvalho, F.D.; Mendonça, S.F.D.; Rocha, M.N.; Moreira, L.A. Vector Competence of Aedes Aegypti, Aedes Albopictus, and Culex Quinquefasciatus Mosquitoes for Mayaro Virus. PLoS Negl. Trop. Dis. 2020, 14, e0007518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiggins, K.; Eastmond, B.; Alto, B.W. Transmission Potential of Mayaro Virus in Florida Aedes Aegypti and Aedes Albopictus Mosquitoes. Med. Vet. Entomol. 2018, 32, 436–442. [Google Scholar] [CrossRef] [Green Version]
- da Silva Neves, N.A.; da Silva Ferreira, R.; Morais, D.O.; Pavon, J.A.R.; de Pinho, J.B.; Slhessarenko, R.D. Chikungunya, Zika, Mayaro, and Equine Encephalitis Virus Detection in Adult Culicinae from South Central Mato Grosso, Brazil, during the Rainy Season of 2018. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2022, 53, 63–70. [Google Scholar] [CrossRef] [PubMed]
- de Curcio, J.S.; Salem-Izacc, S.M.; Pereira Neto, L.M.; Nunes, E.B.; Anunciação, C.E.; de Paule Silveira-Lacerda, E. Detection of Mayaro Virus in Aedes Aegypti Mosquitoes Circulating in Goiânia-Goiás-Brazil. Microbes Infect. 2022, 24, 104948. [Google Scholar] [CrossRef] [PubMed]
- Maia, L.M.S.; Bezerra, M.C.F.; Costa, M.C.S.; Souza, E.M.; Oliveira, M.E.B.; Ribeiro, A.L.M.; Miyazaki, R.D.; Slhessarenko, R.D. Natural Vertical Infection by Dengue Virus Serotype 4, Zika Virus and Mayaro Virus in Aedes (Stegomyia) Aegypti and Aedes (Stegomyia) Albopictus. Med. Vet. Entomol. 2019, 33, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, M.J.; de Souza, W.M.; de Castro-Jorge, L.A.; de Carvalho, R.V.H.; Castro, Í.d.A.; de Almeida, L.G.N.; Consonni, S.R.; Zamboni, D.S.; Figueiredo, L.T.M. Chikungunya Virus Exposure Partially Cross-Protects against Mayaro Virus Infection in Mice. J. Virol. 2021, 95, e0112221. [Google Scholar] [CrossRef]
- Figueiredo, L.T.M. Human Urban Arboviruses Can Infect Wild Animals and Jump to Sylvatic Maintenance Cycles in South America. Front. Cell. Infect. Microbiol. 2019, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.; Herrero, L.J.; Rudd, P.A.; Mahalingam, S. Mouse Models of Alphavirus-Induced Inflammatory Disease. J. Gen. Virol. 2015, 96, 221–238. [Google Scholar] [CrossRef]
- Nguyet, M.N.; Duong, T.H.K.; Trung, V.T.; Nguyen, T.H.Q.; Tran, C.N.B.; Long, V.T.; Dui, L.T.; Nguyen, H.L.; Farrar, J.J.; Holmes, E.C.; et al. Host and Viral Features of Human Dengue Cases Shape the Population of Infected and Infectious Aedes Aegypti Mosquitoes. Proc. Natl. Acad. Sci. USA 2013, 110, 9072–9077. [Google Scholar] [CrossRef] [Green Version]
- Shaw, W.R.; Catteruccia, F. Vector Biology Meets Disease Control: Using Basic Research to Fight Vector-Borne Diseases. Nat. Microbiol. 2019, 4, 20–34. [Google Scholar] [CrossRef]
- Agboli, E.; Leggewie, M.; Altinli, M.; Schnettler, E. Mosquito-Specific Viruses-Transmission and Interaction. Viruses 2019, 11, E873. [Google Scholar] [CrossRef] [Green Version]
- Caluwé, L.D.; Ariën, K.K.; Bartholomeeusen, K. Host Factors and Pathways Involved in the Entry of Mosquito-Borne Alphaviruses. Trends Microbiol. 2021, 29, 634–647. [Google Scholar] [CrossRef]
- Wu, V.Y.; Chen, B.; Christofferson, R.; Ebel, G.; Fagre, A.C.; Gallichotte, E.N.; Sweeny, A.R.; Carlson, C.J.; Ryan, S.J. A Minimum Data Standard for Vector Competence Experiments. Sci. Data 2022, 9, 634. [Google Scholar] [CrossRef] [PubMed]
- de Melo-Santos, M.A.V.; Sanches, E.G.; de Jesus, F.J.; Regis, L. Evaluation of a New Tablet Formulation Based on Bacillus Thuringiensis Sorovar. Israelensis for Larvicidal Control of Aedes Aegypti. Mem. Inst. Oswaldo Cruz 2001, 96, 859–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorim, L.B.; Helvecio, E.; de Oliveira, C.M.F.; Ayres, C.F.J. Susceptibility Status of Culex Quinquefasciatus (Diptera: Culicidae) Populations to the Chemical Insecticide Temephos in Pernambuco, Brazil. Pest Manag. Sci. 2013, 69, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Chevillon, C.; Albright, R.G.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Scott, T.W. Genetic Specificity and Potential for Local Adaptation between Dengue Viruses and Mosquito Vectors. BMC Evol. Biol. 2009, 9, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, P.P.; Guedes, D.R.D.; Melo-Santos, M.A.V.; Cordeiro, M.T.; Acioli, R.V.; Batista, C.A.V.; Gonçalves, L.S.M.; Souza, M.F.M.; Araújo, Y.V.; Magalhães, F.J.R.; et al. Vector Surveillance for Dengue Virus Detection in the Archipelago of Fernando de Noronha, Brazil. J. Med. Entomol. 2016, 53, 613–619. [Google Scholar] [CrossRef]
- Guedes, D.R.; Paiva, M.H.; Donato, M.M.; Barbosa, P.P.; Krokovsky, L.; Rocha, S.W.D.S.; Saraiva, K.L.; Crespo, M.M.; Rezende, T.M.; Wallau, G.L.; et al. Zika Virus Replication in the Mosquito Culex Quinquefasciatus in Brazil. Emerg. Microbes Infect. 2017, 6, e69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naveca, F.G.; do Nascimento, V.A.; de Souza, V.C.; Nunes, B.T.D.; Rodrigues, D.S.G.; da Costa Vasconcelos, P.F. Multiplexed Reverse Transcription Real-Time Polymerase Chain Reaction for Simultaneous Detection of Mayaro, Oropouche, and Oropouche-like Viruses. Mem. Inst. Oswaldo Cruz 2017, 112, 510–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.Y.; Thay, C.H.; Tin, T.C.; Devi, S. Rapid Detection, Serotyping and Quantitation of Dengue Viruses by TaqMan Real-Time One-Step RT-PCR. J. Virol. Methods 2006, 138, 123–130. [Google Scholar] [CrossRef]
- Kantor, A.M.; Lin, J.; Wang, A.; Thompson, D.C.; Franz, A.W.E. Infection Pattern of Mayaro Virus in Aedes Aegypti (Diptera: Culicidae) and Transmission Potential of the Virus in Mixed Infections With Chikungunya Virus. J. Med. Entomol. 2019, 56, 832–843. [Google Scholar] [CrossRef]
- Long, K.C.; Ziegler, S.A.; Thangamani, S.; Hausser, N.L.; Kochel, T.J.; Higgs, S.; Tesh, R.B. Experimental Transmission of Mayaro Virus by Aedes Aegypti. Am. J. Trop. Med. Hyg. 2011, 85, 750–757. [Google Scholar] [CrossRef] [Green Version]
- Dieme, C.; Ciota, A.T.; Kramer, L.D. Transmission Potential of Mayaro Virus by Aedes Albopictus, and Anopheles Quadrimaculatus from the USA. Parasit. Vectors 2020, 13, 613. [Google Scholar] [CrossRef]
- Secundino, N.F.C.; Chaves, B.A.; Orfano, A.S.; Silveira, K.R.D.; Rodrigues, N.B.; Campolina, T.B.; Nacif-Pimenta, R.; Villegas, L.E.M.; Silva, B.M.; Lacerda, M.V.G.; et al. Zika Virus Transmission to Mouse Ear by Mosquito Bite: A Laboratory Model That Replicates the Natural Transmission Process. Parasit. Vectors 2017, 10, 346. [Google Scholar] [CrossRef] [Green Version]
- Hibl, B.M.; Dailey Garnes, N.J.M.; Kneubehl, A.R.; Vogt, M.B.; Spencer Clinton, J.L.; Rico-Hesse, R.R. Mosquito-Bite Infection of Humanized Mice with Chikungunya Virus Produces Systemic Disease with Long-Term Effects. PLoS Negl. Trop. Dis. 2021, 15, e0009427. [Google Scholar] [CrossRef]
- Weger-Lucarelli, J.; Carrau, L.; Levi, L.I.; Rezelj, V.; Vallet, T.; Blanc, H.; Boussier, J.; Megrian, D.; Coutermarsh-Ott, S.; LeRoith, T.; et al. Host Nutritional Status Affects Alphavirus Virulence, Transmission, and Evolution. PLOS Pathog. 2019, 15, e1008089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín-Lopez, A.; Calvo-Pinilla, E.; Moreno, S.; Utrilla-Trigo, S.; Nogales, A.; Brun, A.; Fikrig, E.; Ortego, J. Modeling Arboviral Infection in Mice Lacking the Interferon Alpha/Beta Receptor. Viruses 2019, 11, E35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constant, L.E.C.; Rajsfus, B.F.; Carneiro, P.H.; Sisnande, T.; Mohana-Borges, R.; Allonso, D. Overview on Chikungunya Virus Infection: From Epidemiology to State-of-the-Art Experimental Models. Front. Microbiol. 2021, 12, 744164. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.M.; Dias, R.S.; de Oliveira, M.D.; Costa, I.C.T.A.; Fernandes, L.d.S.; Pessoa, C.R.; da Matta, S.L.P.; Costa, V.V.; Souza, D.G.; da Silva, C.C.; et al. Animal Model of Arthritis and Myositis Induced by the Mayaro Virus. PLoS Negl. Trop. Dis. 2019, 13, e0007375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, R.L.; Rossi, S.L.; Bergren, N.A.; Plante, K.S.; Weaver, S.C. The Role of Innate versus Adaptive Immune Responses in a Mouse Model of O’Nyong-Nyong Virus Infection. Am. J. Trop. Med. Hyg. 2013, 88, 1170–1179. [Google Scholar] [CrossRef] [Green Version]
- Nem de Oliveira Souza, I.; Frost, P.S.; França, J.V.; Nascimento-Viana, J.B.; Neris, R.L.S.; Freitas, L.; Pinheiro, D.J.L.L.; Nogueira, C.O.; Neves, G.; Chimelli, L.; et al. Acute and Chronic Neurological Consequences of Early-Life Zika Virus Infection in Mice. Sci. Transl. Med. 2018, 10, eaar2749. [Google Scholar] [CrossRef] [Green Version]
- Christian, K.M.; Song, H.; Ming, G.-L. Pathophysiology and Mechanisms of Zika Virus Infection in the Nervous System. Annu. Rev. Neurosci. 2019, 42, 249–269. [Google Scholar] [CrossRef] [Green Version]
- Grubaugh, N.D.; Fauver, J.R.; Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.; Murrieta, R.A.; Gendernalik, A.; Smith, D.R.; Brackney, D.E.; Ebel, G.D. Mosquitoes Transmit Unique West Nile Virus Populations during Each Feeding Episode. Cell Rep. 2017, 19, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Kramer, L.D. Insights into Arbovirus Evolution and Adaptation from Experimental Studies. Viruses 2010, 2, 2594–2617. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Dpi | MID + | IR | Cq Range | SG + | DR | Cq Range |
|---|---|---|---|---|---|---|---|
| Ae. aegypti RecLab | 3 | 44/45 | 97.7% | 17.8–31.5 | 27/44 | 61.3% | 20.2–37.6 |
| 7 | 45/45 | 100% | 17–25.5 | 45/45 | 100% | 17–32.2 | |
| 14 | 45/45 | 100% | 17.5–31.7 | 44/45 | 97.7% | 20–35.1 | |
| 134/135 | 116/134 | ||||||
| Cx. quinquefasciatus CqSLab | 3 | 5/35 | 14.28% | 29–36.8 | 4/5 | 80% | 35.8–37.8 |
| 7 | 5/38 | 13.1% | 31–36.7 | 3/5 | 60% | 35.2–38 | |
| 14 | 4/27 | 14.81 | 35–37.5 | 3/4 | 75% | 25.2–37.3 | |
| 14/100 | 10/14 |
| Species | Group | Mice | Sex | Assay | Clinical Score | Dpi | Viremia (PFU/mL) |
|---|---|---|---|---|---|---|---|
| Ae. aegypti RecLab | Control | 1 | M | 1 | 0 | 3 | N.A. |
| 2 | M | 1 | 0 | 3 | N.A. | ||
| 3 | F | 2 | 0 | 4 | N.A. | ||
| 4 | F | 2 | 0 | 4 | N.A. | ||
| 5 | F | 3 | 0 | 3 | N.A. | ||
| 6 | F | 3 | 0 | 3 | N.A. | ||
| Test | 1 | M | 1 | 5 | 3 | 4 × 108 | |
| 2 | M | 1 | 7 | 3 | 1.5 × 109 | ||
| 3 | M | 1 | 5 | 3 | 5 × 109 | ||
| 4 | M | 1 | 5 | 3 | 1.8 × 109 | ||
| 5 | F | 2 | Mortality | 4 | N.A. | ||
| 6 | F | 2 | Mortality | 4 | N.A. | ||
| 7 | F | 2 | Mortality | 4 | N.A. | ||
| 8 | F | 2 | Mortality | 4 | N.A. | ||
| 9 | F | 3 | 7 | 3 | 4.5 × 108 | ||
| 10 | F | 3 | 7 | 3 | 2.5 × 108 | ||
| 11 | F | 3 | 5 | 3 | 5 × 108 | ||
| 12 | F | 3 | 5 | 3 | 3.5 × 108 | ||
| Cx. quinquefasciatus CqSLab | Control | 1 | F | 1 | 0 | 7 | N.A. |
| 2 | F | 1 | 0 | 7 | N.A. | ||
| 3 | M | 2 | 0 | 7 | N.A. | ||
| 4 | M | 2 | 0 | 7 | N.A. | ||
| Test | 1 | F | 1 | 0 | 7 | N.A. | |
| 2 | F | 1 | 0 | 7 | N.A. | ||
| 3 | F | 1 | 0 | 7 | N.A. | ||
| 4 | F | 1 | 0 | 7 | N.A. | ||
| 5 | M | 2 | 0 | 7 | N.A. | ||
| 6 | M | 2 | 0 | 7 | N.A. | ||
| 7 | M | 2 | 0 | 7 | N.A. | ||
| 8 | M | 2 | 0 | 7 | N.A. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krokovsky, L.; Lins, C.R.B.; Guedes, D.R.D.; Wallau, G.d.L.; Ayres, C.F.J.; Paiva, M.H.S. Dynamic of Mayaro Virus Transmission in Aedes aegypti, Culex quinquefasciatus Mosquitoes, and a Mice Model. Viruses 2023, 15, 799. https://doi.org/10.3390/v15030799
Krokovsky L, Lins CRB, Guedes DRD, Wallau GdL, Ayres CFJ, Paiva MHS. Dynamic of Mayaro Virus Transmission in Aedes aegypti, Culex quinquefasciatus Mosquitoes, and a Mice Model. Viruses. 2023; 15(3):799. https://doi.org/10.3390/v15030799
Chicago/Turabian StyleKrokovsky, Larissa, Carlos Ralph Batista Lins, Duschinka Ribeiro Duarte Guedes, Gabriel da Luz Wallau, Constância Flávia Junqueira Ayres, and Marcelo Henrique Santos Paiva. 2023. "Dynamic of Mayaro Virus Transmission in Aedes aegypti, Culex quinquefasciatus Mosquitoes, and a Mice Model" Viruses 15, no. 3: 799. https://doi.org/10.3390/v15030799
APA StyleKrokovsky, L., Lins, C. R. B., Guedes, D. R. D., Wallau, G. d. L., Ayres, C. F. J., & Paiva, M. H. S. (2023). Dynamic of Mayaro Virus Transmission in Aedes aegypti, Culex quinquefasciatus Mosquitoes, and a Mice Model. Viruses, 15(3), 799. https://doi.org/10.3390/v15030799