
The Others: A Systematic Review of the Lesser-Known Arboviruses of the Insular Caribbean

 , and
, and
Abstract
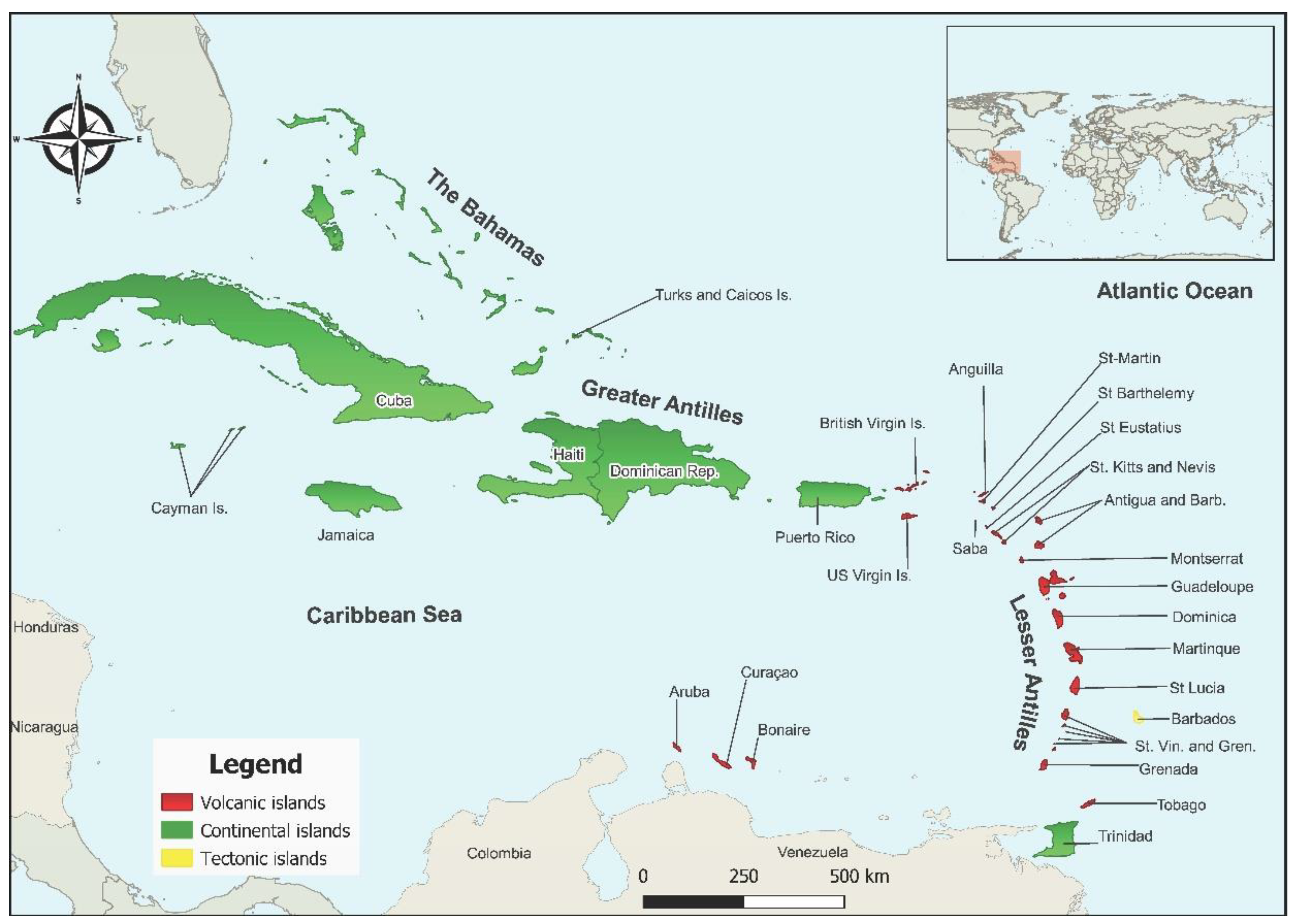
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Selection Criteria
2.3. Screening and Data Extraction
3. Results
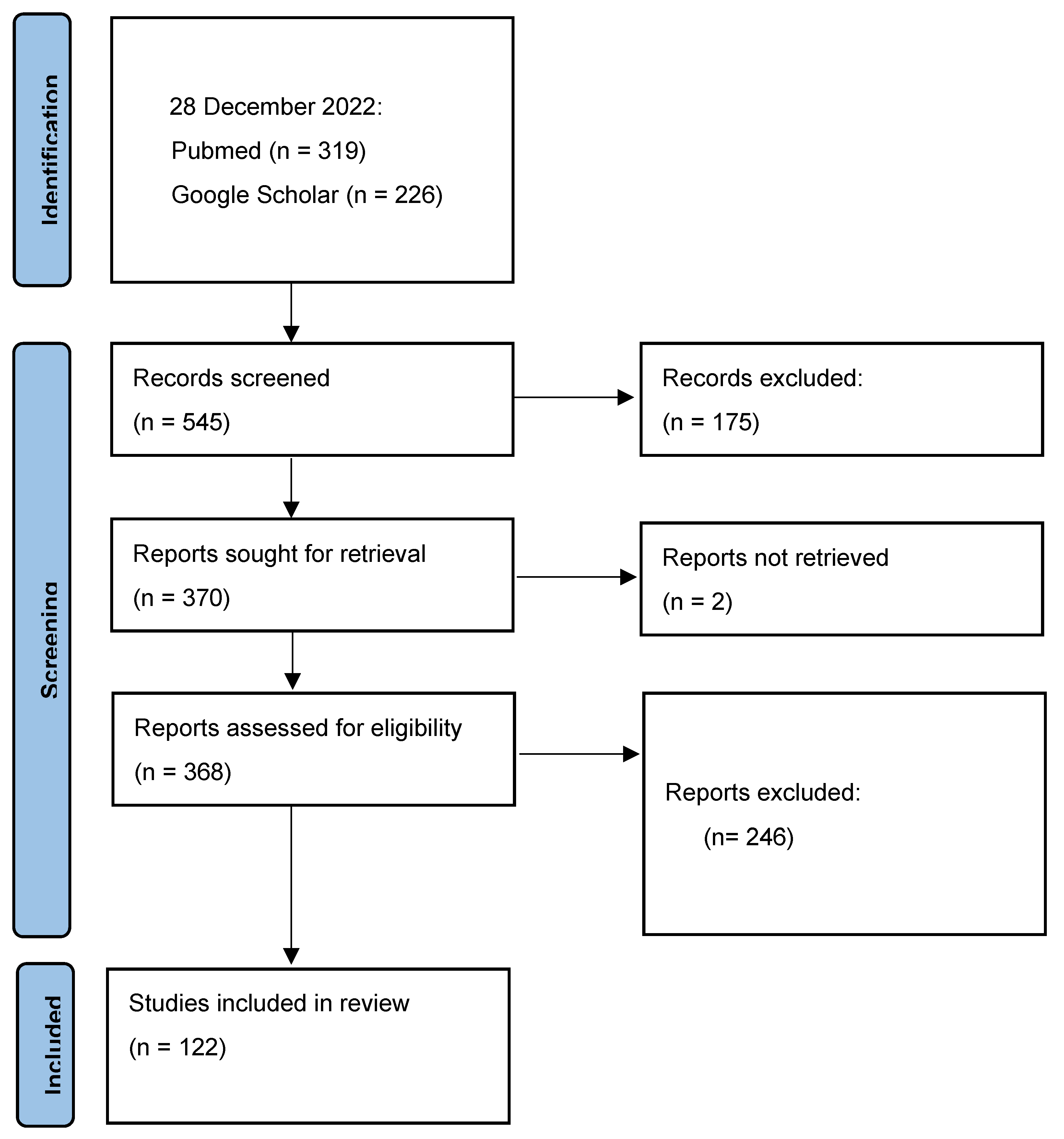
3.1. Study Selection
3.2. Sample Characteristics
3.3. Lesser-Known Arboviruses Identified from the Insular Caribbean
3.3.1. Asfaviridae
African Swine Fever Virus (ASFV)
3.3.2. Flaviviridae
Ilhéus Virus (ILHV)
Japanese Encephalitis Virus (JEV)
St Louis Encephalitis Virus (SLEV)
Spondweni Virus (SPONV)
West Nile Virus (WNV)
3.3.3. Nairoviridae
Estero Real Virus (ERV)
Hughes Virus (HUGV)
Soldado Virus (SOLV)
3.3.4. Peribunyaviridae
Bimiti Virus (BIMV)
Bushbush Virus (BSBV)
Cache Valley Virus (CVV)
Caraparú like Virus (CARV)
Catú Virus (CATUV)
Guamá Virus (GMAV)
Kairi Virus (KRIV)
Lukuni Virus (LUKV)
Manzanilla Virus (MANV)
Melao Virus (MELV)
Moriche Virus (MORV)
Nepuyo Virus (NEPUV)
Oriboca Virus (ORIV)
Oropouche Virus (OROV)
Pacui Virus (PACV)
Restan Virus (RESV)
Triniti Virus (TNTV)
Turlock Virus (TURV)
Wyeomyia Virus (WYOV)
3.3.5. Phenuiviridae
Itaporanga Virus (ITPV)
3.3.6. Rhabdoviridae
Aruac Virus (ARUV)
Cocal Virus (COCV)
3.3.7. Sedoreoviridae
Bluetongue Virus (BTV)
Epizootic Hemorrhagic Disease Virus (EHDV)
Ieri Virus (IERIV)
Wad Medani Virus (WMV)
3.3.8. Togaviridae
Eastern Equine Encephalitis Virus (EEEV)
Madariaga Virus
Mayaro Virus (MAYV)
Mucambo Virus (MUCV)
Una Virus (UNAV)
Venezuelan Equine Encephalitis Virus (VEEV)
Western Equine Encephalitis Virus (WEEV)

{kind=link}
{kind=link}
| Arbovirus | Island | Date | Source | References |
|---|---|---|---|---|
| Flaviviridae | ||||
| Ilhéus virus | Trinidad | 1954 | Mixed pool of mosquitoes, Psorophora spp., | [74] |
| 1955 | Mixed pool of mosquitoes, Homo sapiens | [50,74] | ||
| 1956 | Mixed pool of mosquitoes, Psorophora ferox | [74] | ||
| 1966 | Homo sapiens | [50] | ||
| 1967 | Homo sapiens | [50] | ||
| St Louis encephalitis virus | Haiti | 1955 | Butorides virescens | [85] |
| Jamaica | 1962 | Culex nigripalpus, Mimus polyglottos | [87,89] | |
| 1963 | Culex nigripalpus | [88] | ||
| Trinidad | 1955 | Culex coronator, Culex caudelli, Psorophora ferox | [93] | |
| 1956 | Leptotila verreauxi | [94] | ||
| 1958 | Ramphocelus carbo, Turdus fumigatus, Manacus manacus, mosquitoes, Homo sapiens | [95,96] | ||
| Pipra erythrocephola, mosquitoes | [95] | |||
| 1960–1962 | Aedes scapularis, Culex caudelli, Culex coronator, Culex declarator, Culex nigripalpus, Culex spissipes, Culex taeniopus, Psorophora ferox | [95] | ||
| Spondweni virus | Haiti | 2016 | Culex quinquefasciatus a | [30] |
| West Nile virus | Puerto Rico | 2006 | Gallus gallus, Culex nigripalpus, Culex bahamensis | [104] |
| Nairoviridae | ||||
| Estero real virus | Cuba | 1980 | Ornithodoros tadaridae | [46] |
| Hughes virus | Cuba | 1979 | Ornithodoros denmarki | [115] |
| 1980 | Ornithodoros denmarki | [116] | ||
| Trinidad | 1962 | Ornithodoros capensis, Ornithodoros denmarki, Sterna fuscata | [114] | |
| Soldado virus | Trinidad | 1963 | Ornithodoros capensis | [47] |
| Peribunyaviridae | ||||
| Bimiti virus | Trinidad | 1955 | Culex spp., Culex spissipes | [31] |
| 2007–2009 | Culex portesi | [32] | ||
| Bushbush virus | Trinidad | 1959 | Culex amazonensis | [33] |
| Cache Valley virus b | Jamaica | 1962 | Anopheles grabhami | [35] |
| 1963 | Aedes taeniorhynchus | [35] | ||
| 1965 | Anopheles grabhamii, Aedes taeniorhynchus | [35,36] | ||
| Trinidad | 1958 | Aedes scapularis | [34] | |
| Carapurú like virus | Trinidad | 1959–1964 | Culex portesi, Culex accelerans, Culex amazonensis, Culex coronator, Culex spissipes, Culex nigripalpus, Culex vomerifer, Wyeomyia medioalbipes | [37] |
| 2007–2009 | Culex portesi, Culex vomerifer, Culex pedroi | [32] | ||
| Catú virus | Trinidad | 1960–1964 | Zygodontomys brevicauda, Oryzomys laticeps, Culex portesi, Culex amazonensis, Culex crybda | [119] |
| 1971 | Homo sapiens | [51] | ||
| Guama virus | Trinidad | 1960–1963 | Heteromys anomalus, sentinel mice, Oryzomys, Zygodontomys, Heteromys, Proechimys, Culex amazonensis, Culex portesi, Mansonia spp. | [37] |
| Kairi virus | Trinidad | 1955 | Aedes scapularis, Psorophora ferox, Culex spissipes, Wyeomyia aporonoma | [38] |
| 1958 | Wyeomyia ypsipola | [38] | ||
| Lukuni virus | Trinidad | 1955 | Aedes scapularis | [33] |
| Manzanilla virus | Trinidad | 1954 | Alouatta seniculus insularis | [121] |
| Melao virus | Haiti | 2014 | Homo sapiens | [52] |
| Trinidad | 1955 | Aedes scapularis | [122] | |
| Moriche virus | Trinidad | 1964 | Culex amazonensis | [39] |
| Nepuyo virus | Trinidad | 1957–1958 | Culex accelerans | [123] |
| Oriboca virus | Trinidad | 1962–1963 | Culex portesi, Sentinel mice | [37] |
| 2007–2009 | Culex portesi | [32] | ||
| Oropouche virus | Haiti Trinidad | 2014 1955 | Homo sapiens a Homo sapiens | [52] [53] |
| 1960 | Mansonia venezuelensis b | [53] | ||
| Pacui virus | Trinidad | 1961 | Zygodontomys brevicauda, Oryzomya laticeps | [39] |
| Restan virus | Trinidad | 1963 | Culex portesi | [40] |
| Triniti virus | Trinidad | 1955 | Mixed pool containing Trichoprosopon digitatum, Trichoprosopon theobaldi, Trichoprosopon longipes | [41] |
| Turlock virus | Trinidad | 1963 | Culex declarator | [42] |
| Wyeomyia virus | Trinidad | 1953–1966 | Aedes scapularis, Culex amazonensis, Limatus durhamii, Limatus flavisetosus, Trichoprosopon longipes, Psorophora albipes, Psorophora ferox | [43] |
| 2007–2009 | Trichoprosopon digitatum, Wyeomyia spp. | [32] | ||
| Phenuiviridae | ||||
| Itaporanga virus | Trinidad | Not stated | Culex eastor | [44] |
| Rhabdoviridae | ||||
| Aruac virus | Trinidad | 1955 | Trichoprosopon theobaldi, Psorophora ferox | [45] |
| Cocal virus | Trinidad | 1961 | Orzomys laticeps velutinus, Culex portesi, Heteromys anomalis | [128] |
| Sedoreoviridae | ||||
| Bluetongue virus | Barbados | 1987–1992 | Ovine | [49,133] |
| Dominican Republic | 1987–1992 | Bovine | [49,133] | |
| Guadeloupe | 2006 | Bovine | [135] | |
| Jamaica | 1987–1992 | Bovine | [49,133] | |
| Martinique | 2006 | Bovine | [132] | |
| Puerto Rico | 1987–1992 | Bovine, Culicoides pusillus | [49,133] | |
| Trinidad | 1987–1992 | Bovine | [49,133] | |
| 2017 | Bovine | [138] | ||
| Epizootic haemorrhagic virus | Puerto Rico | 1987–1992 | Culicoides pusillus | [49] |
| Trinidad | 2019 | Bovine | [140] | |
| Ieri virus | Trinidad | 1955 | Psorophora albipes, Psorophora ferox | [33] |
| Wad Medani virus | Jamaica | 1965 | Amblyomma cajennense | [48] |
| Togaviridae | ||||
| Eastern equine encephalitis virus | Cuba | 1943 | Equine | [145] |
| 1953 | Equine | [143] | ||
| 1969–1970 | Equine | [144] | ||
| 1970 | Columba livia | [146] | ||
| 1972 | Equine, Aedes taeniorhynchus, Cyclura macleaya, Egretta thula, Butorides virescens, Guara alba, Buteogallus gunlachi, Tringa solitaria, Saurothera merlini, Tolmarchus caudifasciatus, Corvus nasicus | [143] | ||
| 1980 | Capromys pilorides, Dumetella carolinensis | [147] | ||
| 1949 | Equine | [158] | ||
| Dominican Republic | 1978 | Equine | [148] | |
| Jamaica | 1962 | Equine, Homo sapiens | [150] | |
| Trinidad | 1959 | Culex nigripalpus, Culex taeniopus | [153] | |
| 1970 | Homo sapiens | [55] | ||
| Madariaga virus | Haiti | 2015 | Homo sapiens | [54] |
| Mayaro virus | Haiti | 2014 | Homo sapiens | [57] |
| 2015 | Homo sapiens | [58] | ||
| Trinidad | 1954 | Homo sapiens | [56] | |
| 1957 | Mansonia venezuelensis c | [155] | ||
| Una virus | Trinidad | 1959 | Psorophora ferox | [39] |
| 1960 | Culex ybarmis, Sentinel mice | [39] | ||
| 1962 | Aedes serratus | [39] | ||
| Venezuelan equine encephalitis | Trinidad | 1943 | Mansonia titillans, Equine | [157] |
| 1959–1963 | Culex portesi, Aedes serratus, Culex amazonensis, Limatus flavisetosus, Mansonia, titillans, Mansonia venezuelensis c, Psorophora ferox, Wyeomyia medioalbipes, Orozymus, Zygodontomys, Heteromys, Sentinel mice | [37] | ||
| Mucambo virus | Trinidad | 2007–2009 | Culex portesi | [32] |
| Western equine encephalitis | Cuba | 1970 | Columba livia | [144] |
4. Discussion
4.1. Distribution of Arboviruses in the Caribbean
4.2. Resurgence of Arboviral Threats
4.3. The Need for Increased Regional Capacity
4.4. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization; UNICEF/UNDP/World Bank/World Health Organization Special Programme for Research and Training in Tropical Diseases. Training in Tropical Diseases. In Global Vector Control Response 2017–2030; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization. Arboviruses and Human Disease. In Report of a WHO Scientific Group; World Health Organization Technical Report Series; World Health Organization: Geneva, Switzerland, 1967; Volume 369, pp. 1–84. [Google Scholar]
- Cobo, F. Imported Infectious Diseases: The Impact in Developed Countries; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Centers for Disease Control and Prevention. Arbovirus Catalog. Available online: https://wwwn.cdc.gov/arbocat/ (accessed on 13 August 2022).
- UNEP; Villasol, A.; Beltran, J. Caribbean Islands, GIWA Regional Assessment 4, Global International Waters Assessment; Fortnam, M., Blime, P., Eds.; University of Kalmar: Kalmar, Sweden, 2004. [Google Scholar]
- Heileman, S. Thematic report for the insular Caribbean sub-region. In Proceedings of the CLME Synthesis Workshop, Barbados, February 2007; Available online: https://www.clmeproject.org/phaseone/dcenter/CLME%20Insular%20Caribbean%20Thematic%20Report%20draft%2016%20Feb%2007%5B1%5D.pdf (accessed on 13 August 2022).
- Maxwell, J.C. Geology of Tobago, British West Indies. Geol. Soc. Am. Bull. 1948, 59, 801–854. [Google Scholar] [CrossRef]
- Liang, G.; Gao, X.; Gould, E.A. Factors responsible for the emergence of arboviruses; strategies, challenges and limitations for their control. Emerg. Microbes Infect. 2015, 4, 1–5. [Google Scholar] [CrossRef]
- Pfeffer, M.; Dobler, G. Emergence of zoonotic arboviruses by animal trade and migration. Parasites Vectors 2010, 3, 1–15. [Google Scholar] [CrossRef]
- Tabachnick, W.J. Evolutionary genetics and arthropod-borne disease: The yellow fever mosquito. Am. Entomol. 1991, 37, 14–26. [Google Scholar] [CrossRef]
- Reiter, P. Aedes albopictus and the world trade in used tires, 1988–1995: The shape of things to come? J. Am. Mosq. Control Assoc. 1998, 14, 83–94. [Google Scholar]
- Emmanouil, M.; Evangelidou, M.; Papa, A.; Mentis, A. Importation of dengue, Zika and chikungunya infections in Europe: The current situation in Greece. New Microbes New Infect. 2020, 35, 100663. [Google Scholar] [CrossRef]
- Grossi, P.A.; Percivalle, E.; Campanini, G.; Sarasini, A.; Premoli, M.; Zavattoni, M.; Girello, A.; Dalla Gasperina, D.; Balsamo, M.L.; Baldanti, F. An autochthonous sexually transmitted Zika virus infection in Italy 2016. New Microbiol. 2018, 41, 80–82. [Google Scholar]
- Llagonne-Barets, M.; Icard, V.; Leparc-Goffart, I.; Prat, C.; Perpoint, T.; André, P.; Ramière, C. A case of Mayaro virus infection imported from French Guiana. J. Clin. Virol. 2016, 77, 66–68. [Google Scholar] [CrossRef]
- Norman, F.F.; Henríquez-Camacho, C.; Díaz-Menendez, M.; Chamorro, S.; Pou, D.; Molina, I.; Goikoetxea, J.; Rodríguez-Guardado, A.; Calabuig, E.; Crespillo, C. Imported Arbovirus Infections in Spain, 2009–2018. Emerg. Infect. Dis. 2020, 26, 658. [Google Scholar] [CrossRef]
- Ginnetti, J. Modelling displacement. Forced Migr. Rev. 2015, 10. [Google Scholar]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Aedes aegypti—A review. Mem. Inst. Oswaldo Cruz 2013, 108, 11–17. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef]
- Service, M.W. Agricultural development and arthropod-borne diseases: A review. Rev. Saude Publica 1991, 25, 165–178. [Google Scholar] [CrossRef]
- Stephenson, T.S.; Vincent, L.A.; Allen, T.; Van Meerbeeck, C.J.; McLean, N.; Peterson, T.C.; Taylor, M.A.; Aaron-Morrison, A.P.; Auguste, T.; Bernard, D. Changes in extreme temperature and precipitation in the Caribbean region, 1961–2010. Int. J. Climatol. 2014, 34, 2957–2971. [Google Scholar] [CrossRef]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef]
- Mordecai, E.A.; Caldwell, J.M.; Grossman, M.K.; Lippi, C.A.; Johnson, L.R.; Neira, M.; Rohr, J.R.; Ryan, S.J.; Savage, V.; Shocket, M.S. Thermal biology of mosquito-borne disease. Ecol. Lett. 2019, 22, 1690–1708. [Google Scholar] [CrossRef]
- Hanson, S.M.; Craig, G.B., Jr. Cold acclimation, diapause, and geographic origin affect cold hardiness in eggs of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 1994, 31, 192–201. [Google Scholar] [CrossRef]
- Shelton, R.M. The effect of temperatures on development of eight mosquito species. Mosq. News 1973, 33, 1–12. [Google Scholar]
- Allicock, O.M.; Lemey, P.; Tatem, A.J.; Pybus, O.G.; Bennett, S.N.; Mueller, B.A.; Suchard, M.A.; Foster, J.E.; Rambaut, A.; Carrington, C.V. Phylogeography and population dynamics of dengue viruses in the Americas. Mol. Biol. Evol. 2012, 29, 1533–1543. [Google Scholar] [CrossRef]
- Mowatt, L.; Jackson, S.T. Chikungunya in the Caribbean: An Epidemic in the Making. Infect. Dis. Ther. 2014, 3, 63–68. [Google Scholar] [CrossRef]
- Francis, L.; Hunte, S.A.; Valadere, A.M.; Polson-Edwards, K.; Asin-Oostburg, V.; Hospedales, C.J. Zika virus outbreak in 19 English- and Dutch-speaking Caribbean countries and territories, 2015–2016. Rev. Panam. Salud Publica = Pan Am. J. Public Health 2018, 42, e120. [Google Scholar] [CrossRef]
- Scott, H.H. A History of Tropical Medicine. Based on the Fitzpatrick Lectures Delivered before the Royal College of Physicians of London 1937–1938; Edward Arnold & Co.: London, UK, 1939. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- White, S.K.; Lednicky, J.A.; Okech, B.A.; Morris, J.G., Jr.; Dunford, J.C. Spondweni virus in field-caught Culex quinquefasciatus mosquitoes, Haiti, 2016. Emerg. Infect. Dis. 2018, 24, 1765. [Google Scholar] [CrossRef]
- Spence, L.; Anderson, C.R.; Aitken, T.H.; Downs, W.G. Bimiti virus, a new agent isolated from Trinidadian mosquitoes. Am. J. Trop. Med. Hyg. 1962, 11, 414–417. [Google Scholar] [CrossRef]
- Auguste, A.J.; Adams, A.P.; Arrigo, N.C.; Martinez, R.; Travassos da Rosa, A.P.; Adesiyun, A.A.; Chadee, D.D.; Tesh, R.B.; Carrington, C.V.; Weaver, S.C. Isolation and characterization of sylvatic mosquito-borne viruses in Trinidad: Enzootic transmission and a new potential vector of Mucambo virus. Am. J. Trop. Med. Hyg. 2010, 83, 1262–1265. [Google Scholar] [CrossRef]
- Spence, L.; Anderson, C.R.; Aitken, T.H.; Downs, W.G. Bushbush, Ieri and Lukuni viruses, three unrelated new agents isolated from Trinidadian forest mosquitoes. Proc. Soc. Exp. Biol. Med. 1967, 125, 45–50. [Google Scholar] [CrossRef]
- Downs, W.G.; Spence, L.; Aitken, T.H.; Whitman, L. Cache Valley virus, isolated from a Trinidadian mosquito, Aedes scapularis. West Indian Med. J. 1961, 10, 13–15. [Google Scholar]
- Belle, E.A.; Grant, L.S.; Griffiths, B.B. The isolation of Cache Valley virus from mosquitoes in Jamaica. West Indian Med. J. 1966, 15, 217–220. [Google Scholar]
- Belkin, J.N.; Heinemann, S.J.; Page, W.A. The Culicidae of Jamaica (Mosquito studies. XXI). Contrib. Am. Entomol. Inst. 1970, 6, 1–319. [Google Scholar]
- Jonkers, A.H.; Spence, L.; Downs, W.G.; Aitken, T.H.; Worth, C.B. Arbovirus studies in Bush Bush Forest, Trinidad, W.I., September 1959–December 1964. VI. Rodent-associated viruses (VEE and agents of groups C and Guamá): Isolations and further studies. Am. J. Trop. Med. Hyg. 1968, 17, 285–298. [Google Scholar] [CrossRef]
- Aitken, T.H.; Spence, L.; Manuel, R. Virus Transmission Studies with Tranidadian Mosquitoes IV. Kairi Virus. J. Med. Entomol. 1964, 1, 50–52. [Google Scholar] [CrossRef]
- Jonkers, A.H.; Spence, L.; Downs, W.G.; Aitken, T.H.; Tikasingh, E.S. Arbovirus studies in Bush Bush forest, Trinidad, W.I., September 1959–December 1964. V. Virus isolations. Am. J. Trop. Med. Hyg. 1968, 17, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Jonkers, A.H.; Metselaar, D.; de Andrade, A.H.; Tikasingh, E.S. Restan virus, a new group C arbovirus from Trinidad and Surinam. Am. J. Trop. Med. Hyg. 1967, 16, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Spence, L.; Anderson, C.R.; Aitken, T.H.; Downs, W.G. Triniti VIRUS, a new agent isolated from trinidadian mosquitoes. Am. J. Trop. Med. Hyg. 1964, 13, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Tikasingh, E.S.; Aitken, T.H.; Butcher, L.; Gonzalez, O. Epidemiologic investigations relating to a case of eastern equine encephalitis in a Trinidadian horse. West Indian Med. J. 1968, 17, 90–95. [Google Scholar]
- Aitken, T.H.; Spence, L.; Jonkers, A.H.; Anderson, C.R. Wyeomyia-virus isolations in Trinidad, West Indies. Am. J. Trop. Med. Hyg. 1968, 17, 886–888. [Google Scholar] [CrossRef]
- Scherer, W.F. International Catalogue of Arboviruses Including Certain Other Viruses of Vertebrates 2nd edition, edited by Trygve O. Berge. v + 789 pages. U. S. Department of Health, Education, and Welfare, Public Health Service, Publication No. (CDC) 75-8301. 1975. No price. Am. J. Trop. Med. Hyg. 1976, 25, 204–205. [Google Scholar]
- Spence, L.; Anderson, C.R.; Aitken, T.H.; Downs, W.G. Aruac virus, a new agent isolated from Trinidadian mosquitoes. Am. J. Trop. Med. Hyg. 1966, 15, 231–234. [Google Scholar] [CrossRef]
- Málková, D.; Holubová, J.; Cerný, V.; Daniel, M.; Fernández, A.; de la Cruz, J.; Herrera, M.; Calisher, C.H. Estero real virus: A new virus isolated from argasid ticks Ornithodoros tadaridae in Cuba. Acta Virol. 1985, 29, 247–250. [Google Scholar]
- Jonkers, A.H.; Casals, J.; Aitken, T.H.; Spence, L. Soldado virus, a new agent from Trinidadian Ornithodoros ticks. J. Med. Entomol. 1973, 10, 517–519. [Google Scholar] [CrossRef]
- Belle, E.A.; King, S.D.; Griffiths, B.B.; Grant, L.S. Epidemiological investigation for arboviruses in Jamaica, West Indies. Am. J. Trop. Med. Hyg. 1980, 29, 667–675. [Google Scholar] [CrossRef]
- Mo, C.; Thompson, L.; Homan, E.; Oviedo, M.; Greiner, E.; Gonzalez, J.; Saenz, M. Bluetongue virus isolations from vectors and ruminants in Central America and the Caribbean. Interamerican Bluetongue Team. Am. J. Vet. Res. 1994, 55, 211–215. [Google Scholar]
- Spence, L.; Anderson, C.R.; Downs, W.G. Isolation of Ilheus virus from human beings in Trinidad, West Indies. Trans. R Soc. Trop. Med. Hyg. 1962, 56, 504–509. [Google Scholar] [CrossRef]
- Tikasingh, E.S.; Ardoin, P.; Williams, M.C. First isolation of Catu virus from a human in Trinidad. Trop. Geogr. Med. 1974, 26, 414–416. [Google Scholar]
- Elbadry, M.A.; Durães-Carvalho, R.; Blohm, G.M.; Stephenson, C.J.; Loeb, J.C.; White, S.K.; Telisma, T.; Chavannes, S.; De Beau Rochars, V.M.; Salemi, M. Orthobunyaviruses in the Caribbean: Melao and Oropouche virus infections in school children in Haiti in 2014. PLoS Negl. Trop. Dis. 2021, 15, e0009494. [Google Scholar] [CrossRef]
- Anderson, C.R.; Spence, L.; Downs, W.G.; Aitken, T.H. Oropouche virus: A new human disease agent from Trinidad, West Indies. Am. J. Trop. Med. Hyg. 1961, 10, 574–578. [Google Scholar] [CrossRef]
- Lednicky, J.A.; White, S.K.; Mavian, C.N.; El Badry, M.A.; Telisma, T.; Salemi, M.; OKech, B.A.; De Beau Rochars, V.M.; Morris, J.G., Jr. Emergence of Madariaga virus as a cause of acute febrile illness in children, Haiti, 2015–2016. PLoS Negl. Trop. Dis. 2019, 13, e0006972. [Google Scholar] [CrossRef]
- Corniou, B.; Ardoin, P.; Bartholomew, C.; Ince, W. First isolation of a South American strain of Eastern Equine virus from a case of encephalitis in Trinidad. Trop. Geogr. Med. 1972, 24, 162–167. [Google Scholar]
- Anderson, C.R.; Downs, W.G.; Wattley, G.H.; Ahin, N.W.; Reese, A.A. Mayaro virus: A new human disease agent. II. Isolation from blood of patients in Trinidad, B.W.I. Am. J. Trop. Med. Hyg. 1957, 6, 1012–1016. [Google Scholar] [CrossRef]
- White, S.K.; Mavian, C.; Elbadry, M.A.; De Beau Rochars, V.M.; Paisie, T.; Telisma, T.; Salemi, M.; Lednicky, J.A.; Morris, J.G., Jr. Detection and phylogenetic characterization of arbovirus dual-infections among persons during a chikungunya fever outbreak, Haiti 2014. PLoS Negl. Trop. Dis. 2018, 12, e0006505. [Google Scholar] [CrossRef]
- Lednicky, J.; De Rochars, V.M.; Elbadry, M.; Loeb, J.; Telisma, T.; Chavannes, S.; Anilis, G.; Cella, E.; Ciccozzi, M.; Okech, B.; et al. Mayaro Virus in Child with Acute Febrile Illness, Haiti, 2015. Emerg. Infect. Dis. 2016, 22, 2000–2002. [Google Scholar] [CrossRef]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M.; Ictv Report, C. ICTV Virus Taxonomy Profile: Asfarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef]
- Stegniy, B.; Gerilovich, A.; Buzun, A.; Bisiuk, I.; Kucheryavenko, R.; De Pérez León, A. African Swine Fever: Background, Present Time, and Prospects; ST Druk: Kyiv, Ukraine, 2015; pp. 1–237. [Google Scholar]
- Alexander, F.C. Experiences with African swine fever in Haiti. Ann. N. Y. Acad. Sci. 1992, 653, 251–256. [Google Scholar] [CrossRef]
- Botija, C.S. African Swine Fever: New Developments; Office International des Epizooties: Paris, France, 1982. [Google Scholar]
- Danzetta, M.L.; Marenzoni, M.L.; Iannetti, S.; Tizzani, P.; Calistri, P.; Feliziani, F. African Swine Fever: Lessons to Learn From Past Eradication Experiences. A Systematic Review. Front. Vet. Sci. 2020, 7, 296. [Google Scholar] [CrossRef]
- USDA. USDA Statement on Confirmation of African Swine Fever in the Dominican Republic. Available online: https://www.aphis.usda.gov/aphis/newsroom/news/sa_by_date/sa-2021/asf-confirm (accessed on 12 September 2022).
- USDA. USDA Statement on Confirmation of African Swine Fever in Haiti. Available online: https://www.aphis.usda.gov/aphis/newsroom/stakeholder-info/sa_by_date/sa-2021/sa-09/asf-haiti (accessed on 12 September 2022).
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef]
- Downs, W.G.; Delpeche, K.A.; Uttley, K. Arthropod-Borne Encephalitis Viruses in the West Indies Area. Part IV. A Serological Survey of Antigua, WI. West Indian Med. J. 1963, 12, 109–116. [Google Scholar]
- Weiland, H.; Williams, M.; Hull, B. Serologic survey of dengue and other arboviruses in Curaçao and Aruba, 1973. Bull. Pan Am. Health Organ. 1978, 12, 134–142. [Google Scholar]
- Downs, W.G.; Spence, L.; Borghans, J. Arthropod-Borne Encephalitis Viruses in theWest Indies Area. Part V. A Serological Survey of Curaçao, NWI. Trop. Geogr. Med. 1963, 15, 237–242. [Google Scholar]
- Downs, W.G.; Anderson, C.R. Arthropod-borne encephalitic viruses in the West Indies area. Part I: A serological survey of Grenada, WI. West Indian Med. J. 1959, 8, 101–109. [Google Scholar]
- Downs, W.G.; Spence, L. Arthropod-Borne Encephalitis Viruses in the West Indies Area. Part VII. A Serological Survey of St. Lucia, WI. West Indian Med. J. 1964, 13, 25–32. [Google Scholar]
- Downs, W.G.; Spence, L. Arthropod-Borne Encephalitis Viruses in the West Indies Area. Part VI. A Serological Survey of St. Vincent, WI. West Indian Med. J. 1963, 12, 148–155. [Google Scholar]
- Anderson, C.R.; Downs, W.G.; Theiler, M. Neutralizing antibodies against certain viruses in the sera of residents of Trinidad, B.W.I. Am. J. Trop. Med. Hyg. 1956, 5, 626–641. [Google Scholar] [CrossRef]
- Anderson, C.R.; Aitken, T.H.G.; Downs, W.G. The Isolation of Ilhéus Virus from Wild Caught Forest Mosquitoes in Trinidad. Am. J. Trop. Med. Hyg. 1956, 5, 621–625. [Google Scholar] [CrossRef]
- Verlinde, J.; Molron, J.; Wyler, R. Antibodies that neutralize neurotropic arthropod-borne viruses in residents of Curaçao and Indonesia. Doc. Med. Geogr. Trop. 1955, 7, 94–96. [Google Scholar]
- Downs, W.G.; Anderson, C.R.; Delpeche, K.A.; Byer, M.A. Arthropod-Borne Encephalitis Viruses in the West Indies Area. Part II. A Serological Survey of Barbados, WI. West Indian Med. J. 1962, 11, 117–122. [Google Scholar]
- Tikasingh, E.; Spence, L.; Jonkers, A.H.; Green, A. Arthropod-borne encephalitis viruses in the West Indies area. 8. A serological survey of montserrat. West Indian Med. J. 1966, 15, 112–117. [Google Scholar]
- Komar, O.; Robbins, M.B.; Contreras, G.G.; Benz, B.W.; Klenk, K.; Blitvich, B.J.; Marlenee, N.L.; Burkhalter, K.L.; Beckett, S.; Gonzálvez, G. West Nile virus survey of birds and mosquitoes in the Dominican Republic. Vector-Borne Zoonotic Dis. 2005, 5, 120–126. [Google Scholar] [CrossRef]
- Dupuis, A.P.; Marra, P.P.; Reitsma, R.; Jones, M.J.; Louie, K.L.; Kramer, L.D. Serologic evidence for West Nile virus transmission, Puerto Rico and Cuba. Am. J. Trop. Med. Hyg. 2005, 73, 474–476. [Google Scholar] [CrossRef]
- Dupuis; Alan, P.; Peter, P.; Marra; Kramer., L.D. Serologic evidence of west nile virus transmission, Jamaica, West Indies. Emerg. Infect Dis. 2003, 9, 860–863. [Google Scholar] [CrossRef]
- Komar, N.; Bessoff, K.; Diaz, A.; Amador, M.; Young, G.; Seda, R.; Perez, T.; Hunsperger, E. Avian hosts of West Nile virus in Puerto Rico. Vector-Borne Zoonotic Dis. 2012, 12, 47–54. [Google Scholar] [CrossRef]
- Pelegrino, J.L.; Suarez, M.; Guzman, M.G.; Vazquez, S.; Benitez, N.R. Vigilancia de las encefalitis de San Luis, equina del este y equina del oeste en la provincia Ciego de Avila. Rev. Cuba. Med. Trop. 1996, 48, 109–113. [Google Scholar]
- Pupo, M.; Guzmán, M.G.; Fernández, R.; Llop, A.; Dickinson, F.O.; Pérez, D.; Cruz, R.; González, T.; Estévez, G.; González, H. West Nile virus infection in humans and horses, Cuba. Emerg. Infect. Dis. 2006, 12, 1022. [Google Scholar] [CrossRef]
- Vázquez Ramudo, S.; Guzmán Tirado, M.G.; Monteagudo Borges, R. Serologic study of arbovirus in 2 localities of the Juventud island. Rev. Cuba. Med. Trop. 1989, 41, 362–370. [Google Scholar]
- Stamm, D.D. Susceptibility of bird populations to eastern, western, and St.Louis encephalitis viruses. In Proceedings of the XIII International Ornithological Congress, Ithaca, NY, USA, 17–24 June 1963; pp. 591–603. [Google Scholar]
- McLean, R.; Trevino, H.; Sather, G. Prevalence of selected zoonotic diseases in vertebrates from Haiti, 1972. J. Wildl. Dis. 1979, 15, 327–330. [Google Scholar]
- Belle, E.A.; Grant, L.S.; Page, W.A. The Isolation of St. Louis Encephalitis Virus from Culex nigripalpus Mosquitoes in Jamaica. Am. J. Trop. Med. Hyg. 1964, 13, 452–454. [Google Scholar] [CrossRef]
- Sudia, W.D.; Coleman, P.; Grant, L.S. The Recurrence of St. Louis Encephalitis Virus in Culex nigripalpus Mosquitoes in Jamaica, 1963. Mosq. News 1966, 26, 39–42. [Google Scholar]
- Ventura, A.K. St.Louis encephalitis virus in Jamaican birds. Am. J. Trop. Med. Hyg. 1965, 14, 297–303. [Google Scholar] [CrossRef]
- Sigel, M.; Beasley, A. Studies on Dengue Fever. Tex. Rep. Biol. Med. 1959, 17, 618–623. [Google Scholar]
- Downs, W.; Grant, L. Arthropod-Borne Encephalitis Viruses in the West Indies Area. Part III. A Serological Survey of Jamaica, WI. West Indian Med. J. 1962, 11, 253–264. [Google Scholar]
- Irvine, R.A.; Grant, L.S.; Belle, E.A. Acute viral encephalitis in Jamaica. Am. J. Trop. Med. Hyg. 1963, 12, 916–923. [Google Scholar] [CrossRef]
- Anderson, C.R.; Aitken, T.H.; Downs, W.G.; Spence, L. The isolation of St. Louis virus from Trinidad mosquitoes. Am. J. Trop. Med. Hyg. 1957, 6, 688–692. [Google Scholar] [CrossRef]
- Downs, W.G.; Anderson, C.R.; Casals, J. The isolation of St. Louis virus from a nestling bird in Trinidad, British West Indies. Am. J. Trop. Med. Hyg. 1957, 6, 693–696. [Google Scholar] [CrossRef]
- Aitken, T.H.; Downs, W.G.; Spence, L.; Jonkers, A.H. St.Louis Encephalitis Virus Isolations in Trinidad, West Indies, 1953–1962. Am. J. Trop. Med. Hyg. 1964, 13, 450–451. [Google Scholar] [CrossRef]
- Spence, L.; Downs, W.G.; Boyd, C. Isolation of St. Louis encephalitis virus from the blood of a child in Trinidad, W.I. West Indian Med. J. 1959, 8, 195–198. [Google Scholar]
- Thompson, N.; Auguste, A.; Travassos da Rosa, A.; Carrington, C.; Blitvich, B.; Chadee, D.; Tesh, R.; Weaver, S.; Adesiyun, A. Seroepidemiology of selected alphaviruses and flaviviruses in bats in Trinidad. Zoonoses Public Health 2015, 62, 53–60. [Google Scholar] [CrossRef]
- Thompson, N.N.; Auguste, A.J.; Coombs, D.; Blitvich, B.J.; Carrington, C.V.; da Rosa, A.P.T.; Wang, E.; Chadee, D.D.; Drebot, M.A.; Tesh, R.B. Serological evidence of flaviviruses and alphaviruses in livestock and wildlife in Trinidad. Vector-Borne Zoonotic Dis. 2012, 12, 969–978. [Google Scholar] [CrossRef]
- ProMED. West Nile Viruse, human—Cayman Islands. Available online: https://promedmail.org/promed-post/?id=2200830 (accessed on 2 August 2022).
- Centers for Disease Control and Prevention. West Nile Virus Activity United States—2001. MMWR 2002, 51, 497–520. [Google Scholar]
- Centers for Disease Control and Prevention. Detection of West Nile virus in blood donations—Puerto Rico, 2007. MMWR 2008, 57, 577–580. [Google Scholar]
- Beatty, M.E.; Hunsperger, E.; Long, E.; Schürch, J.; Jain, S.; Colindres, R.; Lerebours, G.; Bernard, Y.-M.; Dobbins, J.G.; Brown, M. Mosquitoborne infections after Hurricane Jeanne, Haiti, 2004. Emerg. Infect. Dis. 2007, 13, 308. [Google Scholar] [CrossRef]
- Weppelmann, T.A.; Burne, A.; von Fricken, M.E.; Elbadry, M.A.; De Rochars, M.B.; Boncy, J.; Okech, B.A. A tale of two Flaviviruses: A Seroepidemiological study of dengue virus and West Nile virus transmission in the Ouest and Sud-Est departments of Haiti. Am. J. Trop. Med. Hyg. 2017, 96, 135. [Google Scholar] [CrossRef]
- Barrera, R.; Hunsperger, E.; Muñoz-Jordán, J.L.; Amador, M.; Diaz, A.; Smith, J.; Bessoff, K.; Beltran, M.; Vergne, E.; Verduin, M. First isolation of West Nile virus in the Caribbean. Am. J. Trop. Med. Hyg. 2008, 78, 666–668. [Google Scholar] [CrossRef]
- Hunsperger, E.A.; McElroy, K.L.; Bessoff, K.; Colón, C.; Barrera, R.; Muñoz-Jordán, J.L. West Nile virus from blood donors, vertebrates, and mosquitoes, Puerto Rico, 2007. Emerg. Infect. Dis. 2009, 15, 1298. [Google Scholar] [CrossRef]
- Anthony, S.J.; Garner, M.M.; Palminteri, L.; Navarrete-Macias, I.; Sanchez-Leon, M.D.; Briese, T.; Daszak, P.; Lipkin, W.I. West Nile virus in the British Virgin Islands. EcoHealth 2014, 11, 255–257. [Google Scholar] [CrossRef]
- Pupo-Antúnez, M.; Cabrera Rodriguez, V.; Vázquez Mojena, Y.; Drebot, M.; Andonova, M.; Dickinson Meneses, F.; Fuentes Gonzalez, O.; Pérez Rodriguez, A.; Santos Montero, P. Estudio serológico en localidades cubanas con infecciones confirmadas al virus del Nilo Occidental. Rev. Cuba. Med. Trop. 2011, 63, 227–230. [Google Scholar]
- Komar, O.; Robbins, M.B.; Klenk, K.; Blitvich, B.J.; Marlenee, N.L.; Burkhalter, K.L.; Gubler, D.J.; Gonzálvez, G.; Peña, C.J.; Peterson, A.T. West Nile virus transmission in resident birds, Dominican Republic. Emerg. Infect. Dis. 2003, 9, 1299. [Google Scholar] [CrossRef]
- Quirin, R.; Salas, M.; Zientara, S.; Zeller, H.; Labie, J.; Murri, S.; Lefrançois, T.; Petitclerc, M.; Martinez, D. West nile virus, Guadeloupe. Emerg. Infect. Dis. 2004, 10, 706. [Google Scholar] [CrossRef]
- Lefrançois, T.; Blitvich, B.J.; Pradel, J.; Molia, S.; Vachiéry, N.; Pallavicini, G.; Marlenee, N.L.; Zientara, S.; Petitclerc, M.; Martinez, D. West Nile virus surveillance, Guadeloupe, 2003–2004. Emerg. Infect. Dis. 2005, 11, 1100. [Google Scholar] [CrossRef]
- Bolfa, P.; Jeon, I.; Loftis, A.; Leslie, T.; Marchi, S.; Sithole, F.; Beck, C.; Lecollinet, S.; Zientara, S.; Hans, A. Detection of West Nile Virus and other common equine viruses in three locations from the Leeward Islands, West Indies. Acta Trop. 2017, 174, 24–28. [Google Scholar] [CrossRef]
- Komar, N.; Clark, G.G. West Nile virus activity in Latin America and the Caribbean. Rev. Panam. Salud Publica 2006, 19, 112–117. [Google Scholar] [CrossRef]
- Garrison, A.R.; Alkhovsky, S.V.; Avšič-Županc, T.; Bente, D.A.; Bergeron, É.; Burt, F.; Di Paola, N.; Ergünay, K.; Hewson, R.; Kuhn, J.H.; et al. ICTV Virus Taxonomy Profile: Nairoviridae. J. Gen. Virol. 2020, 101, 798–799. [Google Scholar] [CrossRef]
- Aitken, T.H.; Jonkers, A.H.; Tikasingh, E.S.; Worth, C.B. Hughes virus from Trinidadian ticks and terns. J. Med. Entomol. 1968, 5, 501–503. [Google Scholar] [CrossRef]
- Danielová, V.; Marhoul, Z.; Dusbábek, F.; Ryba, J.; Fernández, A.; De La Cruz, J.; Abreu, R.; Herrera, M.; Rodriquez, P.; Cantelar, N. Isolation of Hughes virus from ticks in Cuba. Acta Virol. 1982, 26, 186–189. [Google Scholar]
- Cerný, V.; Málkova, D. Results of the Czechoslovak-Cuban cooperation concerning investigations of ticks and arboviruses transmitted by ticks. Folia Parasitol. 1989, 36, 93–96. [Google Scholar]
- Hughes, H.R.; Adkins, S.; Alkhovskiy, S.; Beer, M.; Blair, C.; Calisher, C.H.; Drebot, M.; Lambert, A.J.; de Souza, W.M.; Marklewitz, M.; et al. ICTV Virus Taxonomy Profile: Peribunyaviridae. J. Gen. Virol. 2020, 101, 1–2. [Google Scholar] [CrossRef]
- Groseth, A.; Vine, V.; Weisend, C.; Guevara, C.; Watts, D.; Russell, B.; Tesh, R.B.; Ebihara, H. Maguari Virus Associated with Human Disease. Emerg. Infect. Dis. 2017, 23, 1325–1331. [Google Scholar] [CrossRef]
- Spence, L.; Jonkers, A.H.; Grant, L.S. Arboviruses in the Caribbean Islands. Prog. Med. Virol. 1968, 10, 415–486. [Google Scholar]
- Anderson, C.R.; Aitken, T.H.; Spence, L.P.; Downs, W.G. Kairi virus, a new virus from Trinidadian forest mosquitoes. Am. J. Trop. Med. Hyg. 1960, 9, 70–72. [Google Scholar] [CrossRef]
- Anderson, C.R.; Spence, L.P.; Downs, W.G.; Aitken, T.H. Manzanilla virus: A new virus isolated from the blood of a howler monkey in Trinidad, W.I. Am. J. Trop. Med. Hyg. 1960, 9, 78–80. [Google Scholar] [CrossRef]
- Spence, L.; Anderson, C.R.; Aitken, T.H.; Downs, W.G. Melao virus, a new agent isolated from Trinidadian mosquitoes. Am. J. Trop. Med. Hyg. 1962, 11, 687–690. [Google Scholar] [CrossRef]
- Spence, L.; Anderson, C.R.; Aitken, T.H.; Downs, W.G. Nepuyo virus, a new group C agent isolated in Trinidad and Brazil. I. Isolation and properties of the Trinidadian strain. Am. J. Trop. Med. Hyg. 1966, 15, 71–74. [Google Scholar] [CrossRef]
- Shope, R.E.; Whitman, L. Nepuyo virus, a new group C agent isolated in Trinidad and Brazil. II. Serological studies. Am. J. Trop. Med. Hyg. 1966, 15, 772–774. [Google Scholar] [CrossRef]
- Price, J.L. Serological evidence of infection of Tacaribe virus and arboviruses in Trinidadian bats. Am. J. Trop. Med. Hyg. 1978, 27, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Leventhal, S.S.; Wilson, D.; Feldmann, H.; Hawman, D.W. A Look into Bunyavirales Genomes: Functions of Non-Structural (NS) Proteins. Viruses 2021, 13, 314. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Freitas-Astúa, J.; Bejerman, N.; Blasdell, K.R.; Breyta, R.; Dietzgen, R.G.; Fooks, A.R.; Kondo, H.; Kurath, G.; Kuzmin, I.V.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae 2022. J. Gen. Virol. 2022, 103, 001689. [Google Scholar] [CrossRef] [PubMed]
- Jonkers, A.H.; Shope, R.E.; Aitken, T.H.; Spence, L. Cocal Virus, a New Agent in Trinidad Related to Vesicular Stomatitis Virus, Type Indiana. Am. J. Vet. Res. 1964, 25, 236–242. [Google Scholar]
- Matthijnssens, J.; Attoui, H.; Bányai, K.; Brussaard, C.; Danthi, P.; del Vas, M.; Dermody, T.; Duncan, R.; Fāng, Q.; Johne, R. ICTV Virus Taxonmy Profile: Sedoreoviridae. J. Gen. Virol. 2022, 103, 001782. [Google Scholar] [CrossRef]
- Greiner, E.C.; Mo, C.L.; Homan, E.J.; Gonzalez, J.; Oviedo, M.T.; Thompson, L.H.; Gibbs, E.P. Epidemiology of bluetongue in Central America and the Caribbean: Initial entomological findings. Regional Bluetongue Team. Med. Vet. Entomol. 1993, 7, 309–315. [Google Scholar] [CrossRef]
- Martínez, N.; Alfonso, A.; Barrera, M. Primer reporte de bovinos seroreactores al virus lengua azul en Cuba. Rev. Salud Anim. 2011, 33, 131–133. [Google Scholar]
- MacLachlan, N.J.; Zientara, S.; Stallknecht, D.E.; Boone, J.D.; Goekjian, V.H.; Sailleau, C.; Balasuriya, U.B. Phylogenetic comparison of the S10 genes of recent isolates of bluetongue virus from the United States and French Martinique Island. Virus Res. 2007, 129, 236–240. [Google Scholar] [CrossRef]
- Gibbs, E.P.; Homan, E.J.; Mo, C.L.; Greiner, E.C.; Gonzalez, J.; Thompson, L.H.; Oveido, M.T.; Walton, T.E.; Yuill, T.M. Epidemiology of bluetongue viruses in the American tropics. Regional Bluetongue Team. Ann. N. Y. Acad. Sci. 1992, 653, 243–250. [Google Scholar] [CrossRef]
- Gibbs, E.P.; Greiner, E.C.; Alexander, F.C.; King, T.H.; Roach, C.J. Serological survey of ruminant livestock in some countries of the Caribbean region and South America for antibody to bluetongue virus. Vet. Rec. 1983, 113, 446–448. [Google Scholar] [CrossRef]
- Legisa, D.M.; Gonzalez, F.N.; Dus Santos, M.J. Bluetongue virus in South America, Central America and the Caribbean. Virus Res. 2014, 182, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, H.; Pearson, J.; Klingsporn, A. Bluetongue in cattle: A serologic survey of slaughter cattle in the United States. Am. J. Vet. Res. 1981, 42, 1057. [Google Scholar] [PubMed]
- Reuter, J.D.; Nelson, S.L. Hematologic parameters and viral status for zika, chikungunya, bluetongue, and epizootic hemorrhagic disease in white-tailed deer (Odocoileus virginianus) on St. John, US Virgin Islands. J. Wildl. Dis. 2018, 54, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Brown-Joseph, T.; Batten, C.; Harrup, L.E.; Frost, L.; Flannery, J.; Hicks, H.; Ramkissoon, V.; Ramdeen, R.; Carrington, C.V.; Oura, C.A.L. Bluetongue virus infection in naïve cattle: Identification of circulating serotypes and associated Culicoides biting midge species in Trinidad. Vet. Microbiol. 2017, 211, 1–5. [Google Scholar] [CrossRef]
- Gumm, I.; Taylor, W.; Roach, C.; Alexander, F.; Greiner, E.; Gibbs, E. Serological survey of ruminants in some Caribbean and South American countries for type-specific antibody to bluetongue and epizootic haemorrhagic disease viruses. Vet. Rec. 1984, 114, 635. [Google Scholar] [CrossRef]
- Brown-Joseph, T.; Rajko-Nenow, P.; Hicks, H.; Sahadeo, N.; Harrup, L.E.; Carrington, C.V.; Batten, C.; Oura, C.A.L. Identification and characterization of epizootic hemorrhagic disease virus serotype 6 in cattle co-infected with bluetongue virus in Trinidad, West Indies. Vet. Microbiol. 2019, 229, 1–6. [Google Scholar] [CrossRef]
- Chen, R.; Mukhopadhyay, S.; Merits, A.; Bolling, B.; Nasar, F.; Coffey, L.L.; Powers, A.; Weaver, S.C.; Ictv Report, C. ICTV Virus Taxonomy Profile: Togaviridae. J. Gen. Virol. 2018, 99, 761–762. [Google Scholar] [CrossRef]
- Guilbride, P.D.L. Veterinary Public Health. The Importance of Animal Disease to Public Health in the Caribbean with special reference to Jamaica. Part III. Virus Infections. West Indian Med. J. 1953, 2, 11–36. [Google Scholar]
- Berezin, V. Characteristics of the ecology of the eastern equine encephalomyelitis virus in the Republic of Cuba. Vopr. Virusol. 1977, Jan 1, 62–70. [Google Scholar]
- Fernandez, A.; Cantelar, N. Desarrollo de epizootías y epidemias de encefalomielitis americanas (EEA) en Cuba, 1918–1972. Rev. Cub. Hig. Epid. 1981, 19, 340–345. [Google Scholar]
- Arenas, R. Aporte al estudio de la Encefalomielitis-infecciosa equina. Bol. Col. Vet. Nac. 1946, 5, 3–11. [Google Scholar]
- Soler Nodarse, M.; Mena Portales, J.; Barrio, G.d. Identificación de una cepa de encefalomielitis equina del este (EEE) aislada de una paloma Columba livia domestica. Rev. Cuba. Med. Trop. 1985, 37, 12–18. [Google Scholar]
- Morier Díaz, L.; Fernández Martínez, A.; Fernández Llanes, R.; Soler Nodarse, M.; de Cantelar Francisco, N. [Ecological considerations in areas of the Isla de la Juventud with possible arbovirus circulation]. Rev. Cuba. Med. Trop. 1984, 36, 187–193. [Google Scholar]
- Calisher, C.H.; Levy-Koenig, E.; Mitchell, C.J.; Cabrera, P.F.A.; Cuevas, L.; Pearson, J.E. Eastern equine encephalitis in the Dominican Republic, 1978. Bull. Pan Am. Health Organ. 1979, 13, 1979. [Google Scholar]
- Hart, K.L.; Keen, D.; Belle, E.A. An Outbreak of Eastern Equine Encephalomyelitis in Jamaica, West Indies. Am. J. Trop. Med. Hyg. 1964, 13, 331–334. [Google Scholar] [CrossRef]
- Belle, E.A.; Grant, L.S.; Thorburn, M.J. An outbreak of eastern equine encephalomyelitis in Jamaica. Am. J. Trop. Med. Hyg. 1964, 13, 335–341. [Google Scholar] [CrossRef]
- Rowe, M.; King, D. Eastern equine encephalitis (EEE) antibody status in St Thomas, Jamaica, 1975. West Indian Med. J. 1976, 25, 146–152. [Google Scholar]
- Griffiths, B.B.; King, S.D.; Anglin, A.; Heslop, O. The role of chickens in the epidemiology of eastern equine encephalomyelitis virus in Jamaica. Res. Vet. Sci. 1978, 25, 200–203. [Google Scholar] [CrossRef]
- Downs, W.G.; Aitken, T.H.; Spence, L. Eastern equine encephalitis virus isolated from Culex nigripalpus in Trinidad. Science 1959, 130, 1471. [Google Scholar] [CrossRef]
- Downs, W.G.; Anderson, C.R. Distribution of Immunity to Mayaro Virus Infection in the West Indies. West Indian Med. J. 1958, 7, 190–194. [Google Scholar]
- Aitken, T.H.; Downs, W.G.; Anderson, C.R.; Spence, L.; Casals, J. Mayaro virus isolated from a Trinidadian mosquito, Mansonia venezuelensis. Science 1960, 131, 986. [Google Scholar] [CrossRef]
- Blohm, G.; Elbadry, M.A.; Mavian, C.; Stephenson, C.; Loeb, J.; White, S.; Telisma, T.; Chavannes, S.; De Rochar, V.M.B.; Salemi, M. Mayaro as a Caribbean traveler: Evidence for multiple introductions and transmission of the virus into Haiti. Int. J. Infect. Dis. 2019, 87, 151–153. [Google Scholar] [CrossRef]
- Tigertt, W.D.; Berge, T.O.; Downs, W.G. Studies on the virus of Venezuelan equine encephalomyelitis in Trinidad, W.I. II. Serological status of human beings, 1954–1958. Am. J. Trop. Med. Hyg. 1962, 11, 835–840. [Google Scholar] [CrossRef]
- Eklund, C.M.; Brennan, J.M.; Bell, J.F. Final report to the Pan American Sanitary Bureau regarding the 1948–1949 outbreak of eastern equine encephalitis in the Dominican Republic. Bol. Oficina Sanit. Panam. (OSP) 1950, 29, 493–516. [Google Scholar]
- Fischer, C.; Jo, W.K.; Haage, V.; Moreira-Soto, A.; de Oliveira Filho, E.F.; Drexler, J.F. Challenges towards serologic diagnostics of emerging arboviruses. Clin. Microbiol. Infect. 2021, 27, 1221–1229. [Google Scholar] [CrossRef]
- Aitken, T.H.; Spence, L.; Jonkers, A.H.; Downs, W.G. A 10-year survey of Trinidadian arthropods for natural virus infections (1953–1963). J. Med. Entomol. 1969, 6, 207–215. [Google Scholar] [CrossRef]
- Pagán-Jiménez, J.R.; Rodríguez-Ramos, R.; Reid, B.A.; van den Bel, M.; Hofman, C.L. Early dispersals of maize and other food plants into the Southern Caribbean and Northeastern South America. Quat. Sci. Rev. 2015, 123, 231–246. [Google Scholar] [CrossRef]
- Walter Reed Biosytematics Unit. Systematic Catelog of Culicidae. Available online: http://www.mosquitocatalog.org/ (accessed on 25 October 2022).
- Basu, A.K.; Basu, M.; Adesiyun, A.A. A review on ticks (Acari: Ixodoidea: Ixodidae, Argasidae), associated pathogens and diseases of Trinidad and Tobago. Acarologia 2012, 52, 39–50. [Google Scholar] [CrossRef]
- Camus, E.; Barre, N. Vector situation of tick-borne diseases in the Caribbean Islands. Vet. Parasitol. 1995, 57, 167–176. [Google Scholar] [CrossRef]
- Gondard, M.; Cabezas-Cruz, A.; Charles, R.A.; Vayssier-Taussat, M.; Albina, E.; Moutailler, S. Ticks and Tick-Borne Pathogens of the Caribbean: Current Understanding and Future Directions for More Comprehensive Surveillance. Front. Cell. Infect. Microbiol. 2017, 7, 490. [Google Scholar] [CrossRef]
- Lorenz, C.; Chiaravalloti-Neto, F. Why are there no human West Nile virus outbreaks in South America? Lancet Reg. Health–Am. 2022, 12, 100276. [Google Scholar] [CrossRef]
- Pan American Health Organization. The feasibility of eradicating Aedes aegypti in the Americas. Rev. Panam. Salud Publica = Pan Am. J. Public Health 1997, 1, 68–72. [Google Scholar] [CrossRef]
- Nelson, M.J. Aedes Aegypti: Biology and Ecology; PNSP/86-64; Pan American Health Organizationpan American Sanitary Bureau Reglonal Office of the World Health Organization: Washington, DC, USA, 1986. [Google Scholar]
- Guedes, R.N.C.; Beins, K.; Navarro Costa, D.; Coelho, G.E.; Bezerra, H. Patterns of insecticide resistance in Aedes aegypti: Meta-analyses of surveys in Latin America and the Caribbean. Pest Manag. Sci. 2020, 76, 2144–2157. [Google Scholar] [CrossRef]
- Rawlins, S.C.; Wan, J.O. Resistance in some Caribbean populations of Aedes aegypti to several insecticides. J. Am. Mosq. Control Assoc. 1995, 11, 59–65. [Google Scholar]
- Alphey, L.; Andreasen, M. Dominant lethality and insect population control. Mol. Biochem. Parasitol. 2002, 121, 173–178. [Google Scholar] [CrossRef]
- Dahmana, H.; Mediannikov, O. Mosquito-borne diseases emergence/resurgence and how to effectively control it biologically. Pathogens 2020, 9, 310. [Google Scholar] [CrossRef]
- Achee, N.L.; Bangs, M.J.; Farlow, R.; Killeen, G.F.; Lindsay, S.; Logan, J.G.; Moore, S.J.; Rowland, M.; Sweeney, K.; Torr, S.J.; et al. Spatial repellents: From discovery and development to evidence-based validation. Malar. J. 2012, 11, 164. [Google Scholar] [CrossRef]
- Chávez, J.H.; Silva, J.R.; Amarilla, A.A.; Figueiredo, L.T.M. Domain III peptides from flavivirus envelope protein are useful antigens for serologic diagnosis and targets for immunization. Biologicals 2010, 38, 613–618. [Google Scholar] [CrossRef]
- Yuan, L.; Huang, X.Y.; Liu, Z.Y.; Zhang, F.; Zhu, X.L.; Yu, J.Y.; Ji, X.; Xu, Y.P.; Li, G.; Li, C.; et al. A single mutation in the prM protein of Zika virus contributes to fetal microcephaly. Science 2017, 358, 933–936. [Google Scholar] [CrossRef]
- Brault, A.C.; Huang, C.Y.; Langevin, S.A.; Kinney, R.M.; Bowen, R.A.; Ramey, W.N.; Panella, N.A.; Holmes, E.C.; Powers, A.M.; Miller, B.R. A single positively selected West Nile viral mutation confers increased virogenesis in American crows. Nat. Genet. 2007, 39, 1162–1166. [Google Scholar] [CrossRef]
- Moudy, R.M.; Meola, M.A.; Morin, L.-L.L.; Ebel, G.D.; Kramer, L.D. A newly emergent genotype of West Nile virus is transmitted earlier and more efficiently by Culex mosquitoes. Am. J. Trop. Med. Hyg. 2007, 77, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Pan American Health Organization. PAHO Warns of the Complex Situation of Dengue in Latin America and the Caribbean. Available online: https://www3.paho.org/hq/index.php?option=com_content&view=article&id=15365:paho-warns-of-the-complex-situation-of-dengue-in-latin-america-and-the-caribbean&Itemid=135&lang=en#gsc.tab=0 (accessed on 11 October 2022).
- Diagne, C.T.; Bengue, M.; Choumet, V.; Hamel, R.; Pompon, J.; Missé, D. Mayaro Virus Pathogenesis and Transmission Mechanisms. Pathogens 2020, 9, 738. [Google Scholar] [CrossRef] [PubMed]
- Files, M.A.; Hansen, C.A.; Herrera, V.C.; Schindewolf, C.; Barrett, A.D.T.; Beasley, D.W.C.; Bourne, N.; Milligan, G.N. Baseline mapping of Oropouche virology, epidemiology, therapeutics, and vaccine research and development. Npj Vaccines 2022, 7, 38. [Google Scholar] [CrossRef]
- Pereira, T.N.; Virginio, F.; Souza, J.I.; Moreira, L.A. Emergent Arboviruses: A Review About Mayaro virus and Oropouche orthobunyavirus. Front. Trop. Dis. 2021, 2, 737436. [Google Scholar] [CrossRef]
- Alarcón-Elbal, P.M.; Rodríguez-Sosa, M.A.; Newman, B.; Sutton, W. The first record of Aedes vittatus (Diptera: Culicidae) in the Dominican Republic: Public health implications of a potential invasive mosquito species in the Americas. J. Med. Entomol. 2020, 57, 2016–2021. [Google Scholar] [CrossRef]
- Díaz-Martínez, I.; Diéguez-Fernández, L.; Santana-Águila, B.; de la Paz, E.M.A.; Ruiz-Domínguez, D.; Alarcón-Elbal, P.M. Nueva introducción de Aedes vittatus (diptera: Culicidae) en la región centro-oriental de cuba: Caracterización ecológica y relevancia médica. Interam. J. Med. Health 2021, 4. [Google Scholar]
- Pagac, B.B.; Spring, A.R.; Stawicki, J.R.; Dinh, T.L.; Lura, T.; Kavanaugh, M.D.; Pecor, D.B.; Justi, S.A.; Linton, Y.-M. Incursion and establishment of the Old World arbovirus vector Aedes (Fredwardsius) vittatus (Bigot, 1861) in the Americas. Acta Trop. 2021, 213, 105739. [Google Scholar] [CrossRef]
- Ali, I.; Mundle, M.; Anzinger, J.J.; Sandiford, S.L. Tiger in the sun: A report of Aedes albopictus in Jamaica. Acta Trop. 2019, 199, 105112. [Google Scholar] [CrossRef]


| Country | Asfaviridae | Flaviviridae | Nairoviridae | Peribunyaviridae | Phenuiviridae | Rhabdoviridae | Sedoreoviridae | Togaviridae | ||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ASFV | ILHV | JEV | SLEV | SPONV | WNV | ERV | HUGV | SOLV | BIMV | BSBV | CVV a | CARV | CATUV | GMAV | KRIV | LUKV | MANV | MELV | MORV | NEPUV | ORIV | OROV | PACV | RESV | TNTV | TURV | WYOV | ITPV | ARUV | COCV | BTV | EHDV | IERIV | WMV | EEEV | MADV b | MAYV | MUCV c | UNAV | VEEV | WEEV | |
| Antigua and Barbuda | X § | X § | X | X | ||||||||||||||||||||||||||||||||||||||
| Aruba | X § | |||||||||||||||||||||||||||||||||||||||||
| Bahamas | X | X | ||||||||||||||||||||||||||||||||||||||||
| Barbados | X § | X | X | |||||||||||||||||||||||||||||||||||||||
| British Virgin Islands | X | |||||||||||||||||||||||||||||||||||||||||
| Cayman Islands | X | |||||||||||||||||||||||||||||||||||||||||
| Cuba | X | X | X | X | X | X | X | X | ||||||||||||||||||||||||||||||||||
| Curaçao | X § | X § | X | X § | ||||||||||||||||||||||||||||||||||||||
| Dominican Republic | X | X | X | X | X | |||||||||||||||||||||||||||||||||||||
| Grenada | X § | X § | X | X | ||||||||||||||||||||||||||||||||||||||
| Guadeloupe | X | X | X | |||||||||||||||||||||||||||||||||||||||
| Haiti | X | X | X | X | X | X | X | X | X | X | ||||||||||||||||||||||||||||||||
| Jamaica | X | X | X | X | X | X | X | |||||||||||||||||||||||||||||||||||
| Martinique | X | X | ||||||||||||||||||||||||||||||||||||||||
| Montserrat | X § | |||||||||||||||||||||||||||||||||||||||||
| Puerto Rico | X | X | X | |||||||||||||||||||||||||||||||||||||||
| St. Eustatius | X | |||||||||||||||||||||||||||||||||||||||||
| St. Kitts and Nevis | X | X | X | |||||||||||||||||||||||||||||||||||||||
| St Lucia | X § | X | X | |||||||||||||||||||||||||||||||||||||||
| St. Vin. and the Gren. | X § | X § | ||||||||||||||||||||||||||||||||||||||||
| Trinidad and Tobago | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | |||||||
| USVI | X | X | ||||||||||||||||||||||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, I.; Alarcόn-Elbal, P.M.; Mundle, M.; Noble, S.A.A.; Oura, C.A.L.; Anzinger, J.J.; Sandiford, S.L. The Others: A Systematic Review of the Lesser-Known Arboviruses of the Insular Caribbean. Viruses 2023, 15, 843. https://doi.org/10.3390/v15040843
Ali I, Alarcόn-Elbal PM, Mundle M, Noble SAA, Oura CAL, Anzinger JJ, Sandiford SL. The Others: A Systematic Review of the Lesser-Known Arboviruses of the Insular Caribbean. Viruses. 2023; 15(4):843. https://doi.org/10.3390/v15040843
Chicago/Turabian StyleAli, Inshan, Pedro M. Alarcόn-Elbal, Marcia Mundle, Simmoy A. A. Noble, Chris A. L. Oura, Joshua J. Anzinger, and Simone L. Sandiford. 2023. "The Others: A Systematic Review of the Lesser-Known Arboviruses of the Insular Caribbean" Viruses 15, no. 4: 843. https://doi.org/10.3390/v15040843
APA StyleAli, I., Alarcόn-Elbal, P. M., Mundle, M., Noble, S. A. A., Oura, C. A. L., Anzinger, J. J., & Sandiford, S. L. (2023). The Others: A Systematic Review of the Lesser-Known Arboviruses of the Insular Caribbean. Viruses, 15(4), 843. https://doi.org/10.3390/v15040843