
Orthopoxvirus Circulation in an Endemic Area in Brazil: Investigation of Infections in Small Mammals during an Absence of Outbreaks

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
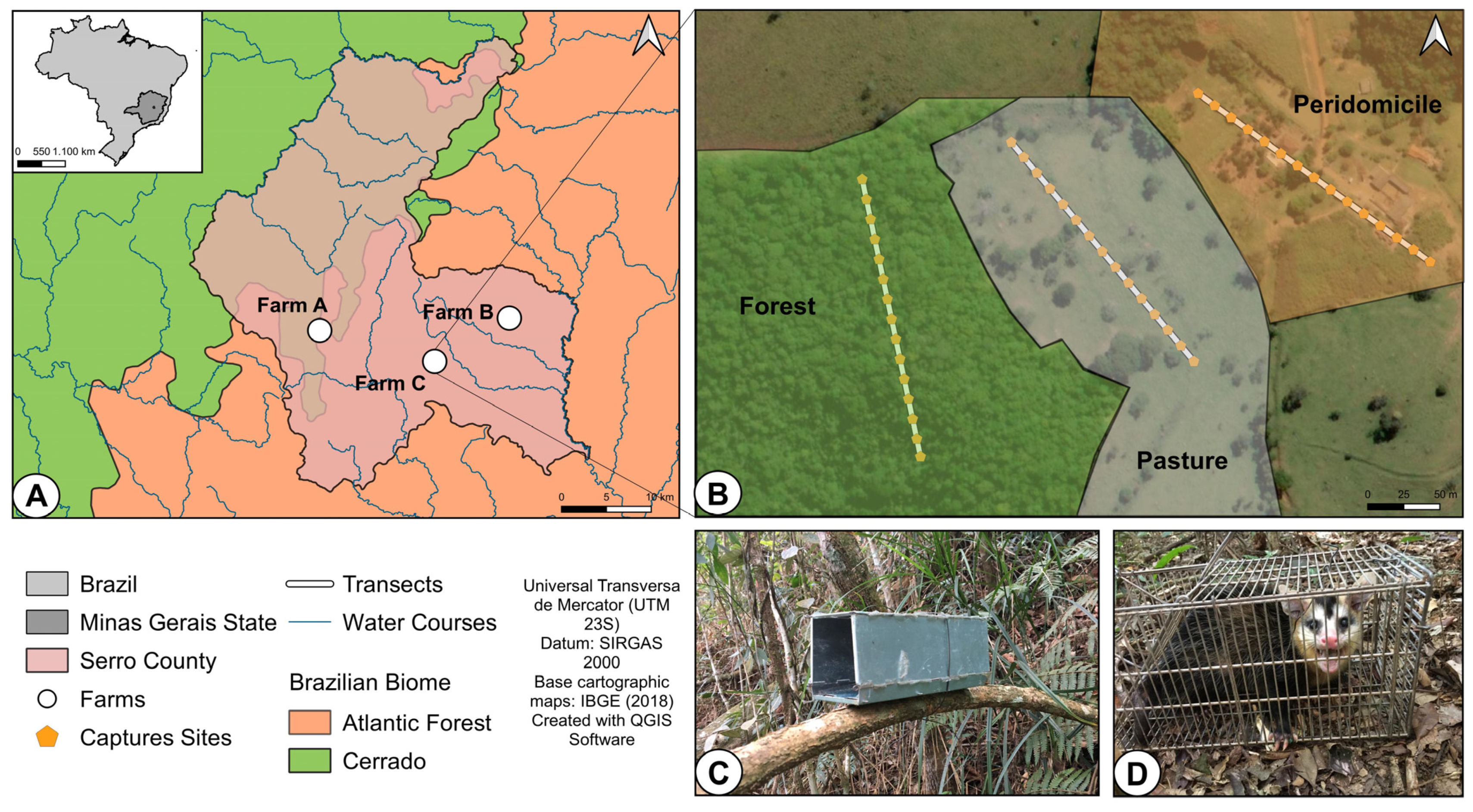
2.1. Small Mammal Sampling Procedures
2.2. Laboratory Analysis
3. Results
3.1. Animal Collection
3.2. Laboratory Findings
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fenner, F.; Henderson, D.A.; Arita, I.; Jezek, Z.; Ladnyi, I.D.; World Health Organization. History of International Public Health. Smallpox and Its Eradication, 6th ed.; World Health Organization: Geneva, Switzerland, 1988; pp. 209–314.
- Kirby, T. WHO celebrates 40 years since eradication of smallpox. Lancet Infect. Dis. 2020, 20, 174. [Google Scholar] [CrossRef] [PubMed]
- Ferrier, A.; Frenois-Veyrat, G.; Schvoerer, E.; Henard, S.; Jarjaval, F.; Drouet, I.; Timera, H.; Boutin, L.; Mosca, E.; Peyrefitte, C.; et al. Fatal Cowpox virus Infection in Human Fetus, France, 2017. Emerg. Infect. Dis. 2021, 27, 2570–2577. [Google Scholar] [CrossRef] [PubMed]
- Durski, K.N.; Mccollum, A.M.; Nakazawa, Y.; Petersen, B.W.; Reynolds, M.; Briand, S.; Djingarey, M.H.; Olson, V.; Damon, I.K.; Khalakdina, A. Emergence of Monkeypox—West and Central Africa, 1970-2017. MMWR. Morb. Mortal. Wkly. Rep. 2018, 67, 306–310. [Google Scholar] [CrossRef]
- Damaso, C.R.A.; Esposito, J.J.; Condit, R.C.; Moussatché, N. An Emergent Poxvirus from Humans and Cattle in Rio de Janeiro State: Cantagalo Virus May Derive from Brazilian Smallpox Vaccine. Virology 2000, 277, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Trindade, G.D.S.; Da Fonseca, F.G.; Marques, J.T.; Nogueira, M.; Mendes, L.C.N.; Borges, A.S.; Peiró, J.R.; Pituco, E.M.; Bonjardim, C.A.; Ferreira, P.C.P.; et al. Araçatuba Virus: A Vaccinia-like Virus Associated with Infection in Humans and Cattle. Emerg. Infect. Dis. 2003, 9, 155–160. [Google Scholar] [CrossRef]
- Lobato, Z.I.P.; Trindade, G.S.; Frois, M.C.M.; Ribeiro, E.B.T.; Dias, G.R.C.; Teixeira, B.M.; Lima, F.A.; Almeida, G.M.F.; Kroon, E.G. Outbreak of exantemal disease caused by Vaccinia virus in human and cattle in Zona da Mata region, Minas Gerais. Arq. Bras. De Med. Vet. E Zootec. 2005, 57, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Schatzmayr, H.G.; De Lemos, E.R.S.; Mazur, C.; Schubach, A.; Majerowicz, S.; Rozental, T.; Schubach, T.M.P.; Bustamante, M.C.; Barth, O.M. Detection of poxvirus in cattle associated with human cases in the State of Rio de Janeiro: Preliminary report. Memórias Do Inst. Oswaldo Cruz 2000, 95, 625–627. [Google Scholar] [CrossRef] [Green Version]
- Matos, A.C.D.; Rehfeld, I.S.; Guedes, M.I.M.C.; Lobato, Z.I.P. Bovine Vaccinia: Insights into the Disease in Cattle. Viruses 2018, 10, 120. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.T.; Oliveira, G.; Afonso, J.A.B.; Souto, R.J.C.; De Mendonça, C.L.; Dantas, A.F.M.; Abrahao, J.S.; Kroon, E.G. An Update on the Known Host Range of the Brazilian Vaccinia Virus: An Outbreak in Buffalo Calves. Front. Microbiol. 2019, 9, 3327. [Google Scholar] [CrossRef] [Green Version]
- Kroon, E.G.; Mota, B.E.F.; Abrahão, J.S.; da Fonseca, F.G.; Trindade, G.D.S. Zoonotic Brazilian Vaccinia Virus: From field to therapy. Antivir. Res. 2011, 92, 150–163. [Google Scholar] [CrossRef] [PubMed]
- da Costa, P.S.P.M.; Oliveira, J.S.; Domingos, I.J.D.S.; e Silva, P.H.B.; Dutra, A.G.S.; Amaral, C.D.; Abrahão, J.S.; Pereira, V.B.R.; Kroon, E.G.; Costa, G.B.; et al. Circulation of Vaccinia Virus in Southern and Southeastern wildlife, Brazil. Transbound. Emerg. Dis. 2020, 67, 1781–1785. [Google Scholar] [CrossRef]
- Peres, M.G.; Bacchiega, T.S.; Appolinário, C.M.; Vicente, A.F.; Mioni, M.S.R.; Ribeiro, B.L.D.; Fonseca, C.R.S.; Pelícia, V.C.; Ferreira, F.; Abrahão, J.S.; et al. Vaccinia Virus in Feces and Urine of Wild Rodents from São Paulo State, Brazil. Viruses 2018, 10, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, I.A.; Reynolds, M.G.; McCollum, A.M.; Figueiredo, P.O.; Ambrosio, L.L.D.; Vieira, F.N.; Costa, G.B.; Matos, A.C.D.; Almeida, V.M.D.A.; Ferreira, P.C.P.; et al. Serological Evidence of Orthopoxvirus Circulation Among Equids, Southeast Brazil. Front. Microbiol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, J.M.A.D.P.; Borges, I.A.; Trindade, G.D.S.; Kroon, E.G.; Cruvinel, T.M.D.A.; Peres, M.G.; Megid, J. Exposure of free-ranging capybaras (Hydrochoerus hydrochaeris) to the Vaccinia Virus. Transbound. Emerg. Dis. 2019, 67, 481–485. [Google Scholar] [CrossRef]
- Miranda, J.B.; Borges, I.A.; Campos, S.P.S.; Vieira, F.N.; De Ázara, T.M.F.; Marques, F.A.; Costa, G.B.; Luis, A.P.M.F.; De Oliveira, J.S.; Ferreira, P.C.P.; et al. Serologic and Molecular Evidence of Vaccinia Virus Circulation among Small Mammals from Different Biomes, Brazil. Emerg. Infect. Dis. 2017, 23, 931–938. [Google Scholar] [CrossRef]
- Peres, M.G.; Bacchiega, T.S.; Appolinário, C.M.; Vicente, A.F.; Mioni, M.D.S.R.; Ribeiro, B.L.D.; Fonseca, C.R.S.; Pelícia, V.C.; Ferreira, F.; Oliveira, G.P.; et al. Vaccinia Virus in Blood Samples of Humans, Domestic and Wild Mammals in Brazil. Viruses 2018, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- D’Anunciação, L.; Guedes, M.I.M.; Oliveira, T.L.; Rehfeld, I.; Bonjardim, C.A.; Ferreira, P.P.; Trindade, G.D.S.; Lobato, Z.P.; Kroon, E.G.; Abrahão, J.S. Filling one more gap: Experimental evidence of horizontal transmission of Vaccinia Virus between bovines and rodents. Vector-Borne Zoonotic Dis. 2012, 12, 61–64. [Google Scholar] [CrossRef]
- Chantrey, J.; Meyer, H.; Baxby, D.; Begon, M.; Bown, K.J.; Hazel, S.M.; Jones, T.; Montgomery, W.I.; Bennett, M. Cowpox: Reservoir hosts and geographic range. Epidemiol. Infect. 1999, 122, 455–460. [Google Scholar] [CrossRef] [Green Version]
- Khodakevich, L.; Jezek, Z.; Kinzanzka, K. Isolation of Monkeypox virus from wild squirrel infected in nature. Lancet 1986, 327, 98–99. [Google Scholar] [CrossRef]
- Hutson, C.L.; Nakazawa, Y.J.; Self, J.; Olson, V.A.; Regnery, R.L.; Braden, Z.; Weiss, S.; Malekani, J.; Jackson, E.; Tate, M.; et al. Laboratory Investigations of African Pouched Rats (Cricetomys gambianus) as a Potential Reservoir Host Species for Monkeypox virus. PLoS Negl. Trop. Dis. 2015, 9, e0004013. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, M.G.; Davidson, W.B.; Curns, A.T.; Conover, C.S.; Huhn, G.; Davis, J.P.; Wegner, M.; Croft, D.R.; Newman, A.; Obiesie, N.N.; et al. Spectrum of Infection and Risk Factors for Human Monkeypox, United States, 2003. Emerg. Infect. Dis. 2007, 13, 1332–1339. [Google Scholar] [CrossRef]
- Essbauer, S.; Pfeffer, M.; Meyer, H. Zoonotic poxviruses. Vet. Microbiol. 2010, 140, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Abrahão, J.S.; Guedes, M.I.M.; Trindade, G.S.; Fonseca, F.G.; Campos, R.K.; Mota, B.F.; Lobato, Z.I.P.; Silva-Fernandes, A.T.; Rodrigues, G.O.L.; Lima, L.S.; et al. One More Piece in the VACV Ecological Puzzle: Could Peridomestic Rodents Be the Link between Wildlife and Bovine Vaccinia Outbreaks in Brazil? PLoS ONE 2009, 4, e7428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peres, M.G.; Bacchiega, T.S.; Appolinário, C.M.; Vicente, A.F.; Allendorf, S.D.; Antunes, J.M.A.P.; Moreira, S.A.; Legatti, E.; Fonseca, C.R.; Pituco, E.M.; et al. Serological study of Vaccinia virus reservoirs in areas with and without official reports of outbreaks in cattle and humans in São Paulo, Brazil. Arch. Virol. 2013, 158, 2433–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trindade, G.D.S.; Emerson, G.L.; Sammons, S.; Frace, M.; Govil, D.; Mota, B.E.F.; Abrahão, J.S.; De Assis, F.L.; Olsen-Rasmussen, M.; Goldsmith, C.S.; et al. Serro 2 Virus Highlights the Fundamental Genomic and Biological Features of a Natural Vaccinia virus Infecting Humans. Viruses 2016, 8, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assis, F.L.; Borges, I.A.; Ferreira, P.C.P.; Bonjardim, C.A.; Trindade, G.D.S.; Lobato, Z.; Guedes, M.I.M.; Mesquita, V.; Kroon, E.; Abrahão, J.S. Group 2 Vaccinia virus, Brazil. Emerg. Infect. Dis. 2012, 18, 2035–2038. [Google Scholar] [CrossRef]
- Costa, G.B.; Augusto, L.T.S.; Leite, J.A.; Ferreira, P.C.P.; Bonjardim, C.A.; Abrahão, J.S.; Kroon, E.G.; Moreno, E.C.; Trindade, G.D.S. Seroprevalence of Orthopoxvirus in rural Brazil: Insights into anti-OPV immunity status and its implications for emergent zoonotic OPV. Virol. J. 2016, 13, 121. [Google Scholar] [CrossRef] [Green Version]
- Brazilian Institute for Geography and Statistics—IBGE. Serro. 2017. Available online: https://cidades.ibge.gov.br/brasil/mg/serro/panorama (accessed on 1 August 2022). (In Portuguese)
- Prefeitura do Serro: Aspectos Gerais. Available online: https://www.serro.mg.gov.br/portal/servicos/1001/aspectos-gerais (accessed on 1 August 2022). (In Portuguese)
- Colli, G.R.; Vieira, C.R.; Dianese, J.C. Biodiversity and conservation of the Cerrado: Recent advances and old challenges. Biodivers. Conserv. 2020, 29, 1465–1475. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J.; Hirota, M.M. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conserv. 2009, 142, 1141–1153. [Google Scholar] [CrossRef]
- Mills, J.N.; Yates, T.L.; Childs, J.E.; Parmenter, R.; Ksiazek, T.G.; Rollin, P.E.; Peters, C.J. Guidelines for Working with Rodents Potentially Infected with Hantavirus. J. Mammal. 1995, 76, 716–722. [Google Scholar] [CrossRef] [Green Version]
- Kroon, E.G.; Abrahão, J.S.; Trindade, G.D.S.; Oliveira, G.P.; Luiz, A.P.M.F.; Costa, G.B.; Lima, M.T.; Calixto, R.S.; de Oliveira, D.B.; Drumond, B.P. Natural Vaccinia virus Infection: Diagnosis, Isolation, and Characterization. Curr. Protoc. Microbiol. 2016, 42, 14A.5.1–14A.5.43. [Google Scholar] [CrossRef]
- Trindade, G.D.S.; Li, Y.; Olson, V.A.; Emerson, G.; Regnery, R.L.; da Fonseca, F.G.; Kroon, E.G.; Damon, I. Real-time PCR assay to identify variants of Vaccinia virus: Implications for the diagnosis of bovine vaccinia in Brazil. J. Virol. Methods 2008, 152, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Olson, V.A.; Laue, T.; Laker, M.T.; Damon, I.K. Detection of Monkeypox virus with real-time PCR assays. J. Clin. Virol. 2006, 36, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Newman, F.K.; Frey, S.E.; Blevins, T.P.; Mandava, M.; Bonifacio, A., Jr.; Yan, L.; Belshe, R.B. Improved Assay To Detect Neutralizing Antibody following Vaccination with Diluted or Undiluted Vaccinia (Dryvax) Vaccine. J. Clin. Microbiol. 2003, 41, 3154–3157. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 7039–7044. [Google Scholar] [CrossRef] [Green Version]
- Meerburg, B.G.; Singleton, G.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef]
- Wolfs, T.F.; Wagenaar, J.A.; Niesters, H.; Osterhaus, A. Rat-to-Human Transmission of Cowpox Infection. Emerg. Infect. Dis. 2002, 8, 1495–1496. [Google Scholar] [CrossRef]
- Campe, H.; Zimmermann, P.; Glos, K.; Bayer, M.; Bergemann, H.; Dreweck, C.; Graf, P.; Weber, B.K.; Meyer, H.; Büttner, M.; et al. Cowpox Virus Transmission from Pet Rats to Humans, Germany. Emerg. Infect. Dis. 2009, 15, 777–780. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Update: Multistate outbreak of monkeypox—Illinois, Indiana, Kansas, Missouri, Ohio, and Wisconsin, 2003. MMWR Morb. Mortal. Wkly Rep. 2003, 52, 561–564. [Google Scholar]
- Paglia, A.P.; Fonseca, G.A.B.d.; Rylands, A.B.; Herrmann, G.; Aguiar, L.M.S.; Chiarello, A.G.; Leite, Y.L.R.; Costa, L.P.; Siciliano, S.; Kierulff, M.C.M.; et al. Lista anotada dos mamíferos do Brasil [In Portuguese]. In Occasional Papers in Conservation Biology; Conservação Internacional: Belo Horizonte, Brazil, 2012. [Google Scholar]
- Bonvicino, J.A.; Oliveira, P.S.D. Guia dos Roedores do Brasil, com Chaves Para Gêneros Baseadas em Caracteres Externos; Centro Pan-Americano de Febre Aftosa—Pan American Health Organization/World Health Organization: Rio de Janeiro, Brazil, 2008. (In Portuguese) [Google Scholar]
- Cerqueira, R. Fatores Ambientais e a Reprodução de Marsupiais e Roedores no Leste do Brasil; Arquivos do Museu Nacional: Rio de Janeiro, Brazil, 2005; Volume 63, pp. 29–39. (In Portuguese) [Google Scholar]
- Bergallo, H.G.; Magnusson, W.E. Effects of Climate and Food Availability on Four Rodent Species in Southeastern Brazil. J. Mammal. 1999, 80, 472–486. [Google Scholar] [CrossRef]
- Loretto, D.; Vieira, M.V. Use of space by the marsupial Marmosops incanus (Didelphimorphia, Didelphidae) in the Atlantic Forest, Brazil. Mamm. Biol. 2008, 73, 255–261. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sample | Capture Date | Weather Conditions | Order | Genus/Species | Sex | Age | Farm | Site of Capture | PRNT * | PCR ** | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Result | Plaque Reduction Percentage | Neutralizing Units per mL | C11R Gene | A56R Gene | E9L Gene | |||||||||
| 46 | 11 May 2021 | Dry | Rodentia | Trinomys cf setosus | Female | Adult | B | Forest | + | 63.51% | 100 NU/mL | - | - | - |
| 84 | 1 July 2021 | Dry | Didelphimorphia | Marmosops incanus | Male | Adult | C | Forest | + | 70.54% | 100 NU/mL | - | - | - |
| 111 | 7 July 2021 | Dry | Rodentia | Nectomys squamipes | Male | Adult | B | Peridomicile | + | 84.48% | 100 NU/mL | - | - | - |
| 142 | 3 August 2021 | Dry | Didelphimorphia | Marmosops incanus | Male | Adult | C | Forest | + | 60.00% | 100 NU/mL | - | - | - |
| 192 | 14 August 2021 | Dry | Rodentia | Oligoryzomys sp. | Unidentifed | Juvenile | A | Pasture | + | 55.45% | 100 NU/mL | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domingos, I.J.S.; Rocha, K.L.S.; Graciano, J.M.; Almeida, L.R.; Doty, J.B.; Paglia, A.P.; Oliveira, D.B.; Nakazawa, Y.J.; Trindade, G.d.S. Orthopoxvirus Circulation in an Endemic Area in Brazil: Investigation of Infections in Small Mammals during an Absence of Outbreaks. Viruses 2023, 15, 842. https://doi.org/10.3390/v15040842
Domingos IJS, Rocha KLS, Graciano JM, Almeida LR, Doty JB, Paglia AP, Oliveira DB, Nakazawa YJ, Trindade GdS. Orthopoxvirus Circulation in an Endemic Area in Brazil: Investigation of Infections in Small Mammals during an Absence of Outbreaks. Viruses. 2023; 15(4):842. https://doi.org/10.3390/v15040842
Chicago/Turabian StyleDomingos, Iago J. S., Kamila L. S. Rocha, Jessica M. Graciano, Lara R. Almeida, Jeffrey B. Doty, Adriano P. Paglia, Danilo B. Oliveira, Yoshinori J. Nakazawa, and Giliane de S. Trindade. 2023. "Orthopoxvirus Circulation in an Endemic Area in Brazil: Investigation of Infections in Small Mammals during an Absence of Outbreaks" Viruses 15, no. 4: 842. https://doi.org/10.3390/v15040842
APA StyleDomingos, I. J. S., Rocha, K. L. S., Graciano, J. M., Almeida, L. R., Doty, J. B., Paglia, A. P., Oliveira, D. B., Nakazawa, Y. J., & Trindade, G. d. S. (2023). Orthopoxvirus Circulation in an Endemic Area in Brazil: Investigation of Infections in Small Mammals during an Absence of Outbreaks. Viruses, 15(4), 842. https://doi.org/10.3390/v15040842