
Phylogeny of Shrew- and Mole-Borne Hantaviruses in Poland and Ukraine
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
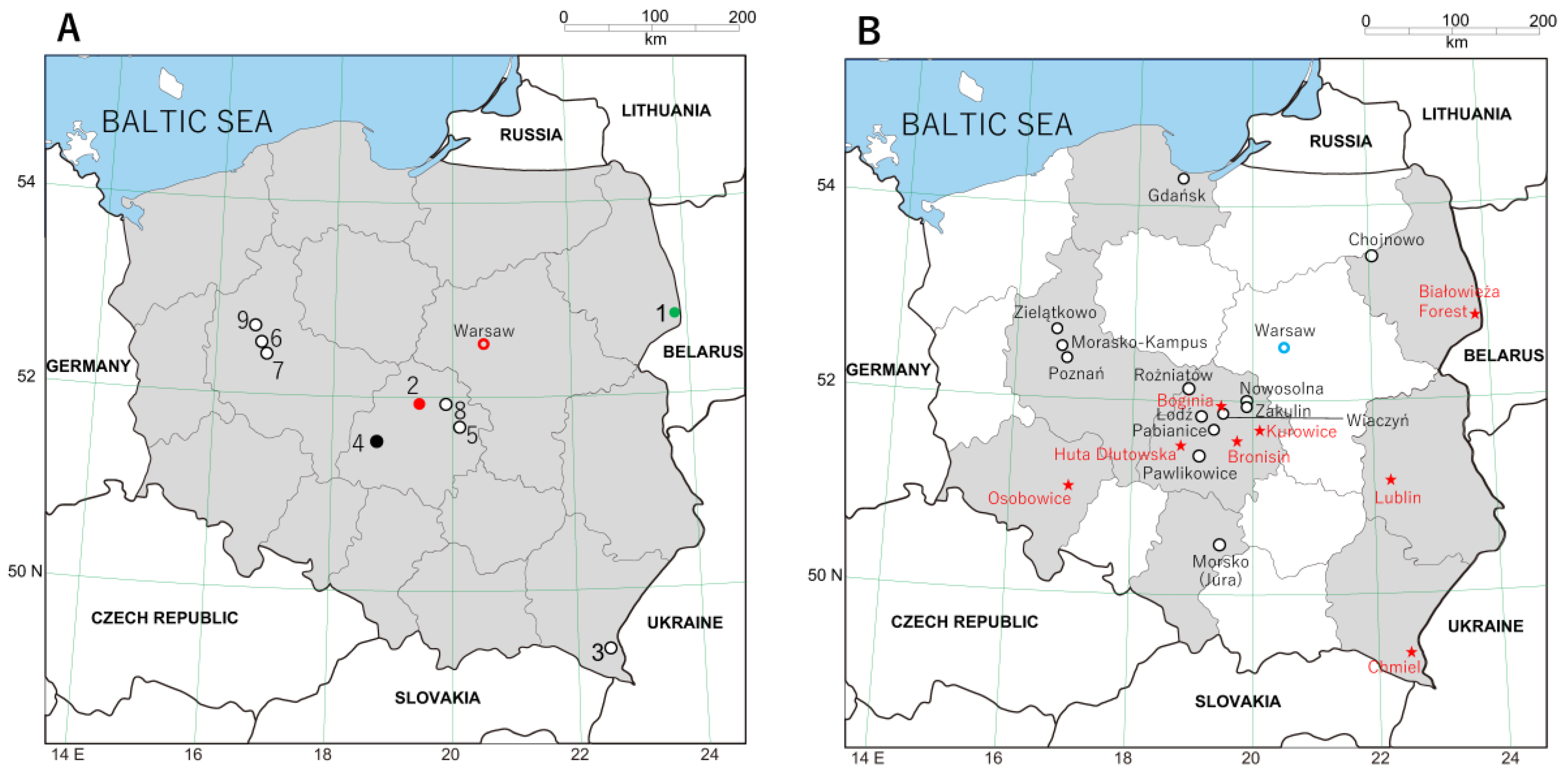
2.1. Trap Sites and Specimen Processing
2.2. Ethics Statement
2.3. RNA Extraction, cDNA Synthesis and RT-PCR Amplification
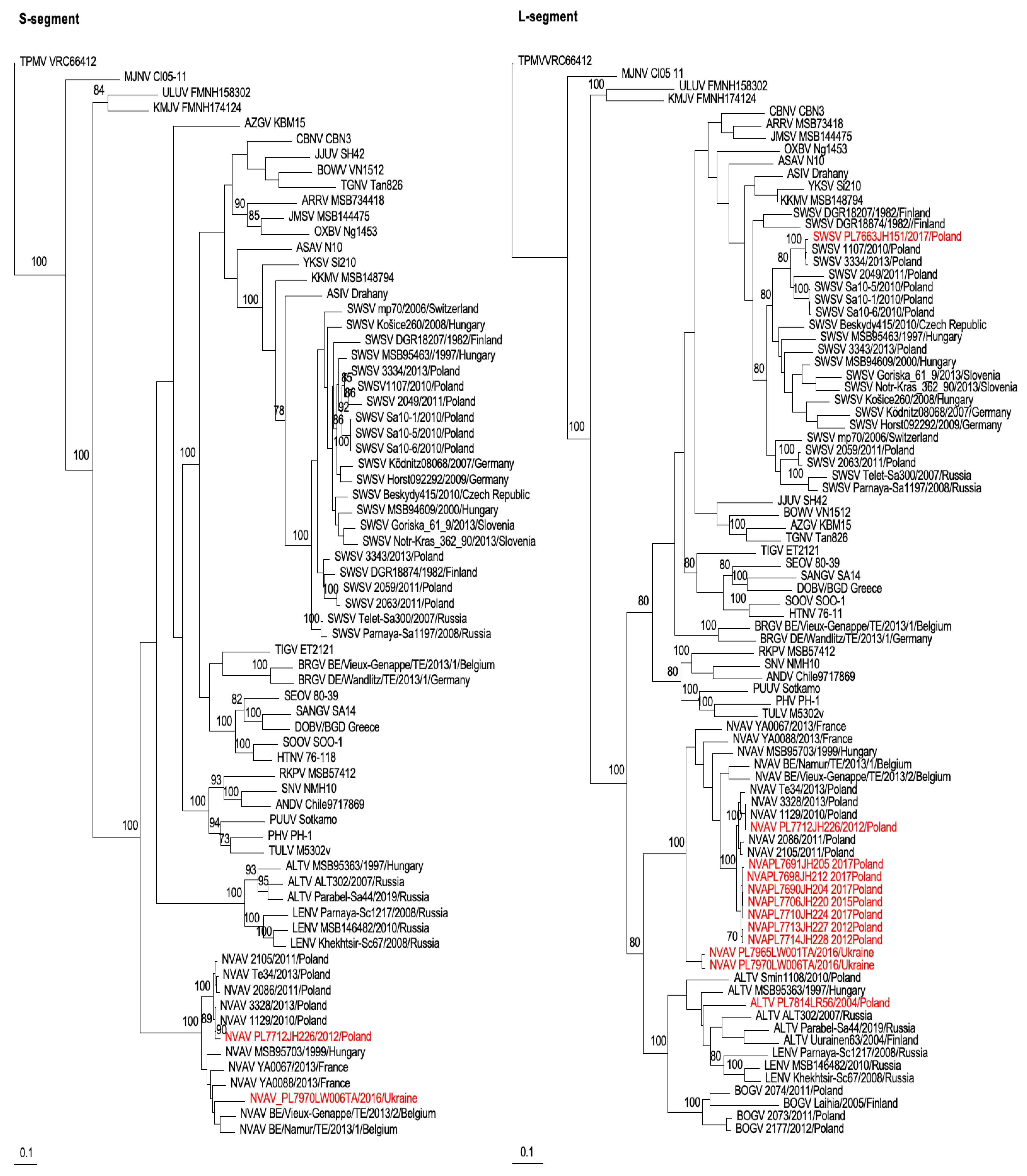
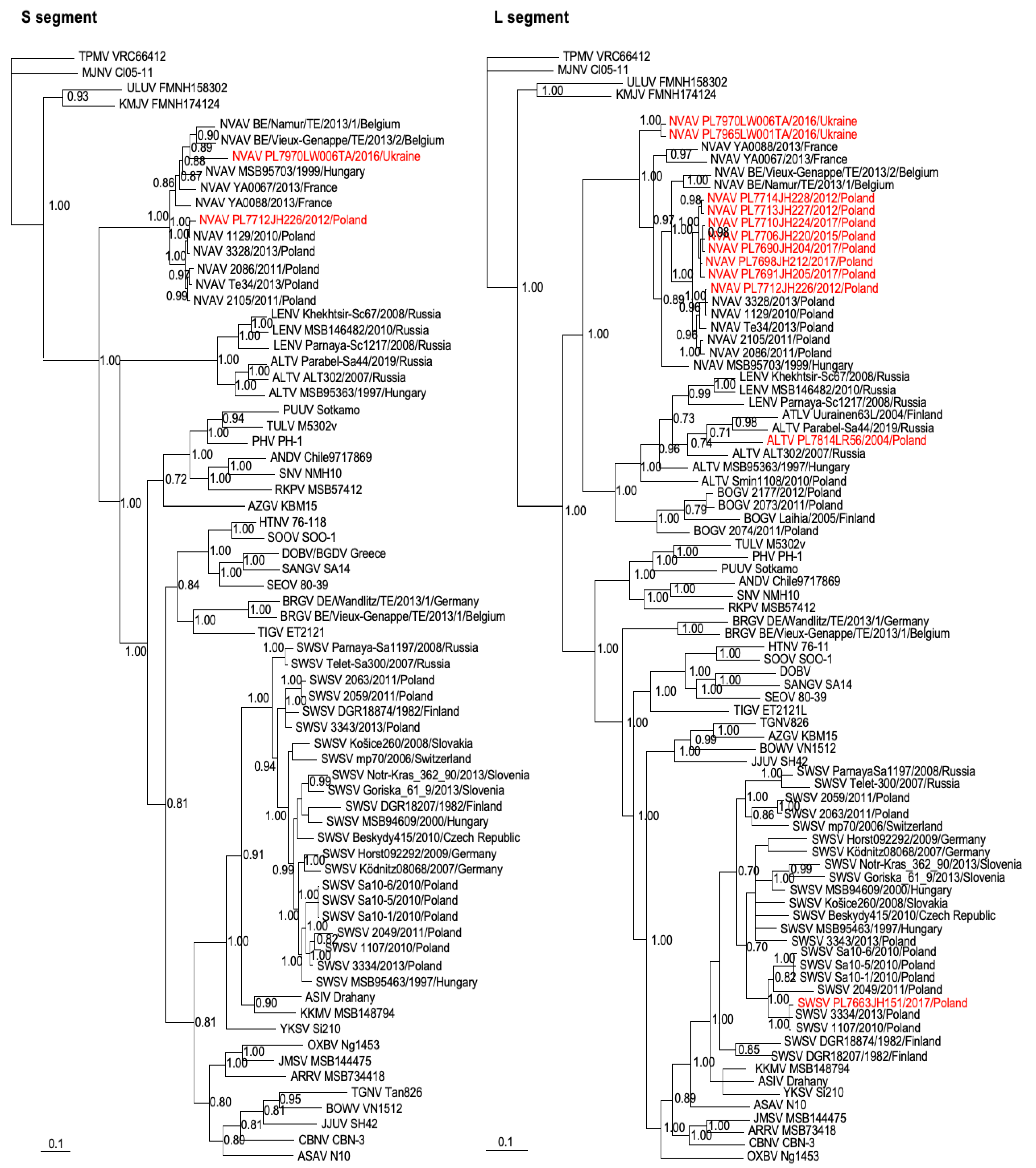
2.4. Genetic and Phylogenetic Analyses
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maes, P.; Adkins, S.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, É.; Blair, C.D.; Briese, T.; et al. Taxonomy of the order Bunyavirales: Second update 2018. Arch. Virol. 2019, 164, 927–941. [Google Scholar] [CrossRef] [Green Version]
- Laenen, L.; Vergote, V.; Calisher, C.H.; Klempa, B.; Klingström, J.; Kuhn, J.H.; Maes, P. Hantaviridae: Current Classification and Future Perspectives. Viruses 2019, 11, 788. [Google Scholar] [CrossRef] [Green Version]
- Plyusnin, A.; Vapalahti, O.; Vaheri, A. Hantaviruses: Genome structure, expression and evolution. J. Gen. Virol. 1996, 77, 2677–2687. [Google Scholar] [CrossRef]
- Khaiboullina, S.F.; Morzunov, S.P.; Jeor, S.C.S. Hantaviruses: Molecular Biology, Evolution and Pathogenesis. Curr. Mol. Med. 2005, 5, 773–790. [Google Scholar] [CrossRef]
- Yanagihara, R.; Gu, S.H.; Arai, S.; Kang, H.J.; Song, J.-W. Hantaviruses: Rediscovery and new beginnings. Virus Res. 2014, 187, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, S.; Yanagihara, R. Genetic Diversity and Geographic Distribution of Bat-borne Hantaviruses. Curr. Issues Mol. Biol. 2020, 39, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Gajdusek, D.C. Hemorrhagic Fevers in Asia: A Problem in Medical Ecology. Geogr. Rev. 1956, 46, 20–42. [Google Scholar] [CrossRef]
- Lähdevirta, J. Nephropathia epidemica in Finland. A clinical histological and epidemiological study. Ann. Clin. Res. 1971, 3, 1–54. [Google Scholar]
- Yanagihara, R.; Gajdusek, D.C. Hemorrhagic fever with renal syndrome: A historical perspective and review of recent advances. In CRC Handbook of Viral and Rickettsial Hemorrhagic Fevers; Gear, J.H.S., Ed.; CRC Press, Inc.: Boca Raton, FL, USA, 1988; pp. 151–188. [Google Scholar]
- Duchin, J.S.; Koster, F.T.; Peters, C.; Simpson, G.L.; Tempest, B.; Zaki, S.R.; Ksiazek, T.G.; Rollin, P.E.; Nichol, S.; Umland, E.T.; et al. Hantavirus Pulmonary Syndrome: A Clinical Description of 17 Patients with a Newly Recognized Disease. N. Engl. J. Med. 1994, 330, 949–955. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yashina, L.N.; Panov, V.V.; Abramov, S.A.; Smetannikova, N.A.; Luchnikova, E.M.; Dupal, T.A.; Krivopalov, A.V.; Arai, S.; Yanagihara, R. Academ virus, a novel hantavirus in the Siberian mole (Talpa altaica) from Russia. Viruses 2022, 14, 309. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Bennett, S.N.; Hope, A.G.; Cook, J.A.; Yanagihara, R. Shared Ancestry between a Newfound Mole-Borne Hantavirus and Hantaviruses Harbored by Cricetid Rodents. J. Virol. 2011, 85, 7496–7503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, S.; Taniguchi, S.; Aoki, K.; Yoshikawa, Y.; Kyuwa, S.; Tanaka-Taya, K.; Masangkay, J.S.; Omatsu, T.; Puentespina, R., Jr.; Watanabe, S.; et al. Molecular phylogeny of a newfound hantavirus harbored by the Geoffroy’s rousette (Rousettus amplexicaudatus), a frugivorous bat species in the Philippines. Infect. Genet. Evol. 2016, 45, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Aoki, K.; Nguyễn, S.T.; Vương, T.T.; Kikuchi, F.; Kinoshita, G.; Fukui, D.; Hoàng, T.T.; Gu, S.H.; Yoshikawa, Y.; et al. Đakrông virus, a novel mobatvirus (Hantaviridae) harbored by the Stoliczka’s Asian trident bat (Aselliscus stoliczkanus) in Vietnam. Sci. Rep. 2019, 9, 10239. [Google Scholar] [CrossRef] [Green Version]
- Straková, P.; Dufkova, L.; Širmarová, J.; Salát, J.; Bartonička, T.; Klempa, B.; Pfaff, F.; Höper, D.; Hoffmann, B.; Ulrich, R.G.; et al. Novel hantavirus identified in European bat species Nyctalus noctula. Infect. Genet. Evol. 2017, 48, 127–130. [Google Scholar] [CrossRef]
- Dafalla, M.; Orłowska, A.; Keleş, S.J.; Straková, P.; Schlottau, K.; Jeske, K.; Hoffmann, B.; Wibbelt, G.; Smreczak, M.; Müller, T.; et al. Hantavirus Brno loanvirus is highly specific to the common noctule bat (Nyctalus noctula) and widespread in Central Europe. Virus Genes 2022, 59, 323–332. [Google Scholar] [CrossRef]
- Gavrilovskaya, I.N.; Apekina, N.S.; Myasnikov, Y.A.; Bernshtein, A.D.; Ryltseva, E.V.; Gorbachkova, E.A.; Chumakov, M.P. Features of circulation of hemorrhagic fever with renal syndrome (HFRS) virus among small mammals in the European U.S.S.R. Arch. Virol. 1983, 75, 313–316. [Google Scholar] [CrossRef]
- Tkachenko, E.A.; Ivanov, A.P.; Donets, M.A.; Miasnikov, Y.A.; Ryltseva, E.V.; Gaponova, L.K.; Bashkirtsev, V.N.; Okulova, N.M.; Drozdov, S.G.; Slonova, R.A.; et al. Potential reservoir and vectors of haemorrhagic fever with renal syndrome (HFRS) in the U. S. S. R. Ann. Soc. Belg. Med. Trop. 1983, 63, 267–269. [Google Scholar]
- Gligić, A.; Stojanović, R.; Obradović, M.; Hlaca, D.; Dimković, N.; Diglisić, G.; Lukac, V.; Ler, Z.; Bogdanović, R.; Antonijević, B.; et al. Hemorrhagic fever with renal syndrome in Yugoslavia: Epidemiologic and epizootiologic features of a nationwide outbreak in 1989. Eur. J. Epidemiol. 1992, 8, 816–825. [Google Scholar] [CrossRef]
- Bennett, S.N.; Gu, S.H.; Kang, H.J.; Arai, S.; Yanagihara, R. Reconstructing the evolutionary origins and phylogeography of hantaviruses. Trends Microbiol. 2014, 22, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.H.; Markowski, J.; Kang, H.J.; Hejduk, J.; Sikorska, B.; Liberski, P.P.; Yanagihara, R. Boginia virus, a newfound hantavirus harbored by the Eurasian water shrew (Neomys fodiens) in Poland. Virol. J. 2013, 10, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, S.H.; Hejduk, J.; Markowski, J.; Kang, H.J.; Markowski, M.; Połatyńska, M.; Sikorska, B.; Liberski, P.P.; Yanagihara, R. Co-circulation of soricid- and talpid-borne hantaviruses in Poland. Infect. Genet. Evol. 2014, 28, 296–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.J.; Gu, S.H.; Yashina, L.N.; Cook, J.A.; Yanagihara, R. Highly divergent molecular variants of soricid-borne Altai virus (Hantaviridae) in Eurasia suggest ancient host-switching events. Viruses 2019, 11, 857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H.; No, J.S.; Kim, W.-K.; Gajda, E.; Perec-Matysiak, A.; Kim, J.-A.; Hildebrand, J.; Yanagihara, R.; Song, J.-W. Molecular Epidemiology and Genetic Diversity of Orthohantaviruses in Small Mammals in Western Poland. Am. J. Trop. Med. Hyg. 2020, 103, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Kikuchi, F.; Hejduk, J.; Markowski, J.; Tanaka-Taya, K.; Sikorska, B.; Morikawa, S.; Suzuki, M.; Liberski, P.P.; Yanagihara, R. Failure to detect hantavirus in vesper bats in Poland. Acta Sci. Microbiol. 2020, 3, 1–5. [Google Scholar]
- Demchyshyna, I.V.; Glass, G.E.; Hluzd, O.A.; Kutseva, V.V.; Taylor, M.K.; Williams, E.P.; Kurpita, V.; Jonsson, C.B. Cocircultion of two orthohantavirus species in small mammals in the northwestern Ukraine. J. Wildl. Dis. 2020, 56, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.P.; Taylor, M.K.; Demchyshyna, I.; Nebogatkin, I.; Nesterova, O.; Khuda, I.; Chernenko, L.; Hluzd, O.A.; Kutseva, V.V.; Glass, G.E.; et al. Prevalence of Hantaviruses Harbored by Murid Rodents in Northwestern Ukraine and Discovery of a Novel Puumala Virus Strain. Viruses 2021, 13, 1640. [Google Scholar] [CrossRef]
- Kang, H.J.; Arai, S.; Hope, A.G.; Song, J.-W.; Cook, J.A.; Yanagihara, R. Genetic diversity and phylogeography of Seewis virus in the Eurasian common shrew in Finland and Hungary. Virol. J. 2009, 6, 208. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.J.; Bennett, S.N.; Sumibcay, L.; Arai, S.; Hope, A.G.; Mocz, G.; Song, J.-W.; Cook, J.A.; Yanagihara, R. Evolutionary Insights from a Genetically Divergent Hantavirus Harbored by the European Common Mole (Talpa europaea). PLoS ONE 2009, 4, e6149. [Google Scholar] [CrossRef]
- Gu, S.H.; Nicolas, V.; Lalis, A.; Sathirapongsasuti, N.; Yanagihara, R. Complete genome sequence analysis and molecular phylogeny of a newfound hantavirus harbored by the Doucet’s musk shrew (Crocidura douceti) in Guinea. Infect. Genet. Evol. 2013, 20, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Faliński, J.B. Vegetation dynamics in temperate lowland primeval forest. Ecological studies in Białowieża Forest. Geobotany 1986, 8, 1–357. [Google Scholar]
- Kujawa, A.; Orczewska, A.; Falkowski, M.; Blicharska, M.; Bohdan, A.; Buchholz, L.; Chylarecki, P.; Gutowski, J.M.; Latałowa, M.; Mysłajek, R.W.; et al. The Białowieża Forest–a UNESCO Natural Heritage Site–protection priorities. For. Res. Pap. 2016, 77, 302–332. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.H.; Dormion, J.; Hugot, J.-P.; Yanagihara, R. High prevalence of Nova hantavirus infection in the European mole (Talpa europaea) in France. Epidemiol. Infect. 2014, 142, 1167–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laenen, L.; Vergote, V.; Kafetzopoulou, L.E.; Bokalinga, T.W.; Vassou, D.; Cook, J.A.; Hugot, J.P.; Deboutte, W.; Kang, H.J.; Witkowski, P.T.; et al. A novel hantavirus of the European mole, Bruges virus, is involved in frequent Nova virus co-infections. Genome Biol. Evol. 2018, 10, 45–55. [Google Scholar] [CrossRef]
- Radosa, L.; Schlegel, M.; Gebauer, P.; Ansorge, H.; Heroldová, M.; Jánová, E.; Stanko, M.; Mošanský, L.; Fričová, J.; Pejčoch, M.; et al. Detection of shrew-borne hantavirus in Eurasian pygmy shrew (Sorex minutus) in Central Europe. Infect. Genet. Evol. 2013, 19, 403–410. [Google Scholar] [CrossRef]
- Lee, H.W.; Lee, P.W.; Johnson, K.M. Isolation of the Etiologic Agent of Korean Hemorrhagic Fever. J. Infect. Dis. 1978, 137, 298–308. [Google Scholar] [CrossRef]
- Lee, H.W.; French, G.R.; Tsuchiya, K.; Baek, L.J.; Lee, P.W.; Foulke, R.S. Observations on Natural and Laboratory Infection of Rodents with the Etiologic Agent of Korean Hemorrhagic Fever. Am. J. Trop. Med. Hyg. 1981, 30, 477–482. [Google Scholar] [CrossRef]
- Yanagihara, R.; Amyx, H.L.; Gajdusek, D.C. Experimental infection with Puumala virus, the etiologic agent of nephropathia epidemica, in bank voles (Clethrionomys glareolus). J. Virol. 1985, 55, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.J.; Bennett, S.N.; Dizney, L.; Sumibcay, L.; Arai, S.; Ruedas, L.A.; Song, J.-W.; Yanagihara, R. Host switch during evolution of a genetically distinct hantavirus in the American shrew mole (Neurotrichus gibbsii). Virology 2009, 388, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, S.; Ohdachi, S.D.; Asakawa, M.; Kang, H.J.; Mocz, G.; Arikawa, J.; Okabe, N.; Yanagihara, R. Molecular phylogeny of a newfound hantavirus in the Japanese shrew mole (Urotrichus talpoides). Proc. Natl. Acad. Sci. USA 2008, 105, 16296–16301. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.J.; Gu, S.H.; Cook, J.A.; Yanagihara, R. Dahonggou Creek virus, a divergent lineage of hantavirus harbored by the long-tailed mole (Scaptonyx fusicaudus). Trop. Med. Health 2016, 44, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójcik-Fatla, A.; Zając, V.; Knap, J.P.; Sroka, J.; Cisak, E.; Sawczyn, A.; Dutkiewicz, J. A small-scale survey of hantavirus in mammals from eastern Poland. Ann. Agric. Environ. Med. 2013, 20, 283–286. [Google Scholar]
- Ali, H.S.; Drewes, S.; Sadowska, E.T.; Mikowska, M.; Groschup, M.H.; Heckel, G.; Koteja, P.; Ulrich, R.G. First Molecular Evidence for Puumala Hantavirus in Poland. Viruses 2014, 6, 340–353. [Google Scholar] [CrossRef] [Green Version]
- Sadkowska-Todys, M.; Dudek-Godeau, D.; Kamińska, S.; Baumann-Popczyk, A.; Czerwiński, M.; Kucharczyk, B.; Zieliński, A. Occurrence and maintenance of hantavirus infections among rodent populations in their natural habitat--results of a field study from Podkarpackie province, Poland 2010–2012. Prz. Epidemiol. 2015, 69, 283–288. [Google Scholar]
- Rosenfeld, U.M.; Drewes, S.; Ali, H.S.; Sadowska, E.T.; Mikowska, M.; Heckel, G.; Koteja, P.; Ulrich, R.G. A highly divergent Puumala virus lineage in southern Poland. Arch. Virol. 2017, 162, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Kolodziej, M.; Melgies, A.; Joniec-Wiechetek, J.; Michalski, A.; Nowakowska, A.; Pitucha, G.; Niemcewicz, M. First molecular characterization of Dobrava-Belgrade virus found in Apodemus flavicollis in Poland. Ann. Agric. Environ. Med. 2018, 25, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Knap, J.P.; Trybusz, A. Haemorrhagic fever with renal syndrome (HFRS)—Hantavirus infection disease appearing in Poland. Pol. Merkur. Lek. 2006, 21, 411–417. (In Polish) [Google Scholar]
- Nowakowska, A.; Heyman, P.; Knap, J.P.; Burzyński, W.; Witas, M. The first established focus of hantavirus infection in Poland, 2007. Ann. Agric. Environ. Med. 2009, 16, 79–85. [Google Scholar]
- Gut, A.K.; Gut, R.; Pencuła, M.; Jarosz, M. New cases of suspected HFRS (Hantavirus infection) in south-eastern Poland. Ann. Agric. Environ. Med. 2013, 20, 544–548. [Google Scholar]
- Michalski, A.; Niemcewicz, M.; Bielawska-Drózd, A.; Nowakowska, A.; Gaweł, J.; Pitucha, G.; Joniec, J.; Zielonka, K.; Marciniak-Niemcewicz, A.; Kocik, J. Surveillance of Hantaviruses in Poland: A Study of Animal Reservoirs and Human Hantavirus Disease in Subcarpathia. Vector-Borne Zoonotic Dis. 2014, 14, 514–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GeurtsvanKessel, C.H.; Goeijenbier, M.; Verner-Carlsson, J.; Litjens, E.; Bos, W.-J.; Pas, S.D.; Melo, M.M.; Koopmans, M.; Lundkvist, A.; Reusken, C.B.E.M. Two clinical cases of renal syndrome caused by Dobrava/Saaremaa hantaviruses imported to the Netherlands from Poland and Belarus, 2012–2014. Infect. Ecol. Epidemiol. 2016, 6, 30548. [Google Scholar] [CrossRef]
- Gut, A.K.; Gut, R.; Rymarz, A.; Woźniak-Kosek, A. Hemorrhagic fever with renal syndrome (HFRS) in Poland. Prz. Epidemiol. 2018, 72, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Stojak, J. Hantavirus infections in humans in Poland—Current state of knowledge and perspectives for research. Eur. J. Public Health 2020, 30, 982–985. [Google Scholar] [CrossRef] [PubMed]
- Sadkowska-Todys, M.; Gut, W.; Baumann, A.; Siennicka, J.; Litwińska, B.; Zieliński, A. Occurrence of human hantavirus infections in Poland. Prz. Epidemiol. 2007, 61, 497–503. (In Polish) [Google Scholar]
- Grygorczuk, S.; Pancewicz, S.; Zajkowska, J.; Kondrusik, M.; Swierzbińska, R.; Moniuszko, A.; Pawlak-Zalewska, W. Detection of anti-hantavirus antibodies in forest workers in the north-east of Poland. Prz. Epidemiol. 2008, 62, 531–537. (In Polish) [Google Scholar]
- Tokarska-Rodak, M.; Plewik, D.; Michalski, A.J.; Kołodziej, M.; Mełgieś, A.; Pańczuk, A.; Konon, H.; Niemcewicz, M. Serological surveillance of vector-borne and zoonotic diseases among hunters in eastern Poland. J. Vector Borne Dis. 2016, 53, 355–361. [Google Scholar]
- Kuhn, J.H.; Schmaljohn, C.S. A Brief History of Bunyaviral Family Hantaviridae. Diseases 2023, 11, 38. [Google Scholar] [CrossRef]
- Carey, D.E.; Reuben, R.; Panicker, K.N.; Shope, R.E.; Myers, R.M. Thottapalayam virus: A presumptive arbovirus isolated from a shrew in India. Indian J. Med. Res. 1971, 59, 1758–1760. [Google Scholar]
- Song, J.-W.; Kang, H.J.; Gu, S.H.; Moon, S.S.; Bennett, S.N.; Song, K.-J.; Baek, L.J.; Kim, H.C.; O’Guinn, M.L.; Chong, S.T.; et al. Characterization of Imjin virus, a newly isolated hantavirus from the Ussuri white-toothed shrew (Crocidura lasiura). J. Virol. 2009, 83, 6184–6191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, S.H.; Kumar, M.; Sikorska, B.; Hejduk, J.; Markowski, J.; Markowski, M.; Liberski, P.P.; Yanagihara, R. Isolation and partial characterization of a highly divergent lineage of hantavirus from the European mole (Talpa europaea). Sci. Rep. 2016, 6, 21119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, J.H.; Bradfute, S.B.; Calisher, C.H.; Klempa, B.; Klingström, J.; Laenen, L.; Palacios, G.; Schmaljohn, C.S.; Tischler, N.D.; Maes, P. Pending Reorganization of Hantaviridae to Include Only Completely Sequenced Viruses: A Call to Action. Viruses 2023, 15, 660. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Country | Capture Site | Year(s) | Tested | Positive | Hantavirus |
|---|---|---|---|---|---|---|
| Sorex araneus | Poland | Białowieża Forest | 1990–2005 | 29 | 0 | |
| Boginia | 2017 | 14 | 1 | SWSV | ||
| Chmiel | 2017 | 18 | 0 | |||
| Kurowice | 2017 | 25 | 0 | |||
| Poznań (Morasko-Kampus) | 2017 | 7 | 0 | |||
| Poznań (city center) | 2013–2016 | 24 | 0 | |||
| Sorex minutus | Poland | Białowieża Forest | 1990–2004 | 26 | 1 | ALTV |
| Boginia | 2017 | 18 | 0 | |||
| Chmiel | 2017 | 9 | 0 | |||
| Kurowice | 2017 | 13 | 0 | |||
| Neomys anomalus | Poland | Białowieża Forest | 1992–2009 | 58 | 0 | |
| Chmiel | 2017 | 7 | 0 | |||
| Neomys fodiens | Poland | Białowieża Forest | 1999–2009 | 29 | 0 | |
| Boginia | 2017 | 2 | 0 | |||
| Chmiel | 2017 | 1 | 0 | |||
| Kurowice | 2017 | 13 | 0 | |||
| Poznań (Morasko-Kampus) | 2013–2016 | 14 | 0 | |||
| Zielątkowo | 2017 | 13 | 0 | |||
| Talpa europaea | Poland | Huta Dłutowska | 2012–2017 | 24 | 8 | NVAV |
| Kurowice | 2017 | 1 | 0 | |||
| Zakulin | 2017 | 1 | 0 | |||
| Ukraine | Lviv | 2016 | 10 | 2 | NVAV |
| Capture Site | Province | Sorex araneus | Sorex minutus | Neomys anomalus | Neomys fodiens | Talpa europaea |
|---|---|---|---|---|---|---|
| Białowieża Forest | Podlasie | 0/29 | 1/26 | 0/58 | 0/29 | |
| Boginia | Łódź | 3/33 | 0/24 | 2/4 | 0/4 | |
| Bronisin | Łódź | 1/3 | ||||
| Chmiel | Subcarpathia | 6/36 | 1/30 | 1/19 | 0/1 | |
| Chojnowo | Podlasie | 0/1 | ||||
| Gdańsk | Pomerania | 0/2 | ||||
| Huta Dłutowska | Łódź | 4/18 | 0/9 | 2/2 | 46/82 | |
| Morsko (Jura) | Silesia | 0/1 | ||||
| Kurowice | Łódź | 3/38 | 3/40 | 3/29 | 0/1 | |
| Łódź | Łódź | 0/1 | ||||
| Lublin | Lublin | 1/8 | ||||
| Nowosolna | Łódź | 0/1 | 0/5 | |||
| Osobowice | Lower Silesia | 3/8 | ||||
| Pabianice | Łódź | 0/1 | 0/1 | |||
| Pawlikowice | Łódź | 0/1 | ||||
| Poznań (city center) | Greater Poland | 0/24 | 0/14 | |||
| Poznań (Morasko-Kampus) | Greater Poland | 0/7 | ||||
| Rożniatów | Łódź | 0/1 | ||||
| Wiączyń | Łódź | 0/1 | ||||
| Zakulin | Łódź | 0/1 | ||||
| Zielątkowo | Greater Poland | 0/13 | ||||
| Hantavirus | SWSV | SWSV ATLV | SWSV | BOGV | NVAV |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kikuchi, F.; Arai, S.; Hejduk, J.; Hayashi, A.; Markowski, J.; Markowski, M.; Rychlik, L.; Khodzinskyi, V.; Kamiya, H.; Mizutani, T.; et al. Phylogeny of Shrew- and Mole-Borne Hantaviruses in Poland and Ukraine. Viruses 2023, 15, 881. https://doi.org/10.3390/v15040881
Kikuchi F, Arai S, Hejduk J, Hayashi A, Markowski J, Markowski M, Rychlik L, Khodzinskyi V, Kamiya H, Mizutani T, et al. Phylogeny of Shrew- and Mole-Borne Hantaviruses in Poland and Ukraine. Viruses. 2023; 15(4):881. https://doi.org/10.3390/v15040881
Chicago/Turabian StyleKikuchi, Fuka, Satoru Arai, Janusz Hejduk, Ai Hayashi, Janusz Markowski, Marcin Markowski, Leszek Rychlik, Vasyl Khodzinskyi, Hajime Kamiya, Tetsuya Mizutani, and et al. 2023. "Phylogeny of Shrew- and Mole-Borne Hantaviruses in Poland and Ukraine" Viruses 15, no. 4: 881. https://doi.org/10.3390/v15040881
APA StyleKikuchi, F., Arai, S., Hejduk, J., Hayashi, A., Markowski, J., Markowski, M., Rychlik, L., Khodzinskyi, V., Kamiya, H., Mizutani, T., Suzuki, M., Sikorska, B., Liberski, P. P., & Yanagihara, R. (2023). Phylogeny of Shrew- and Mole-Borne Hantaviruses in Poland and Ukraine. Viruses, 15(4), 881. https://doi.org/10.3390/v15040881