
Transmitted HIV Drug Resistance in Bulgaria Occurs in Clusters of Individuals from Different Transmission Groups and Various Subtypes (2012–2020)

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Specimen Preparation
2.2. Sequence Analyses
2.3. Statistical Analysis
3. Results
3.1. Study Population
3.2. High HIV-1 Diversity in Drug-Naive Patients in Bulgaria
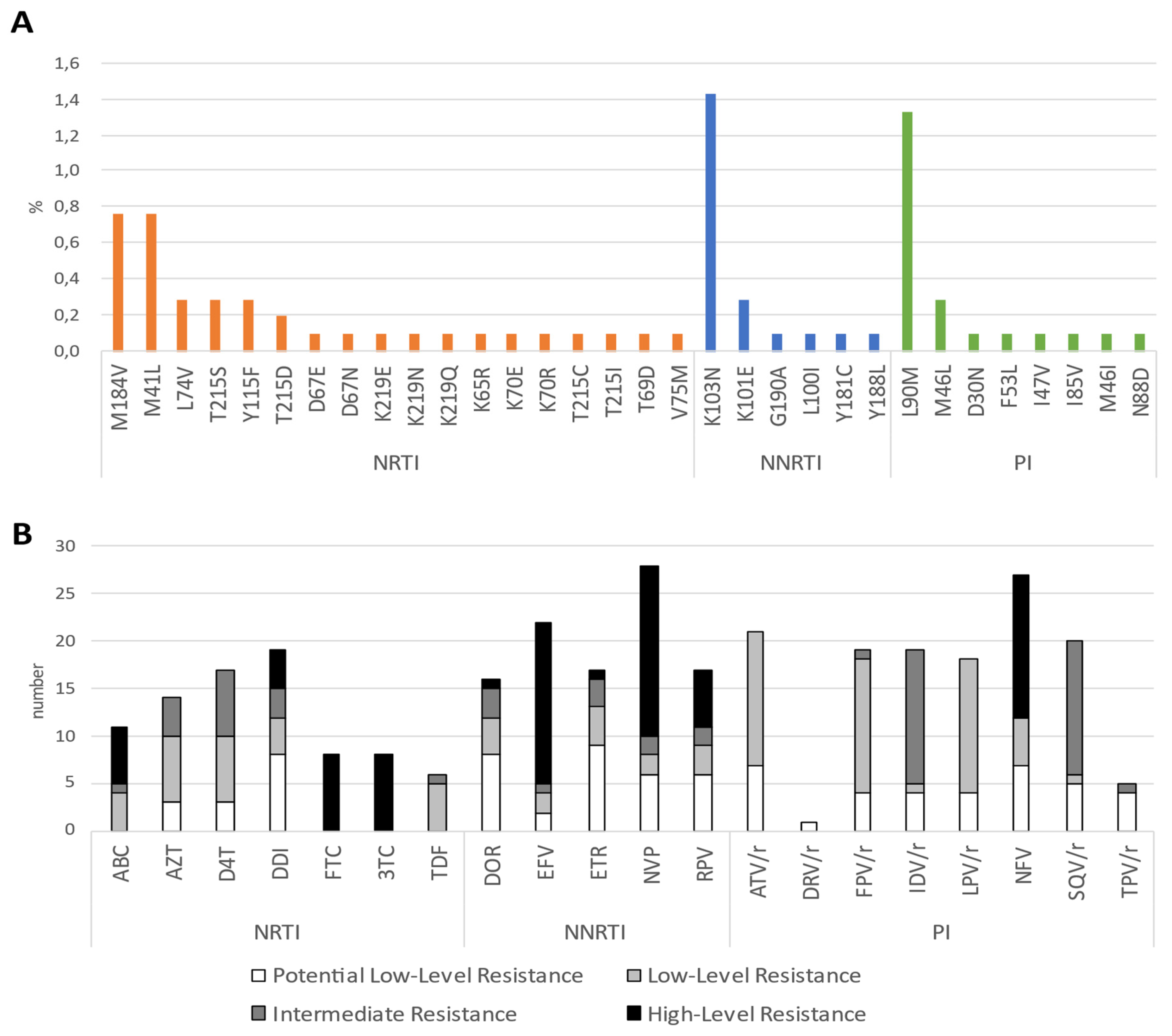
3.3. Evidence for Transmitted Drug Resistance in Drug-Naive Patients
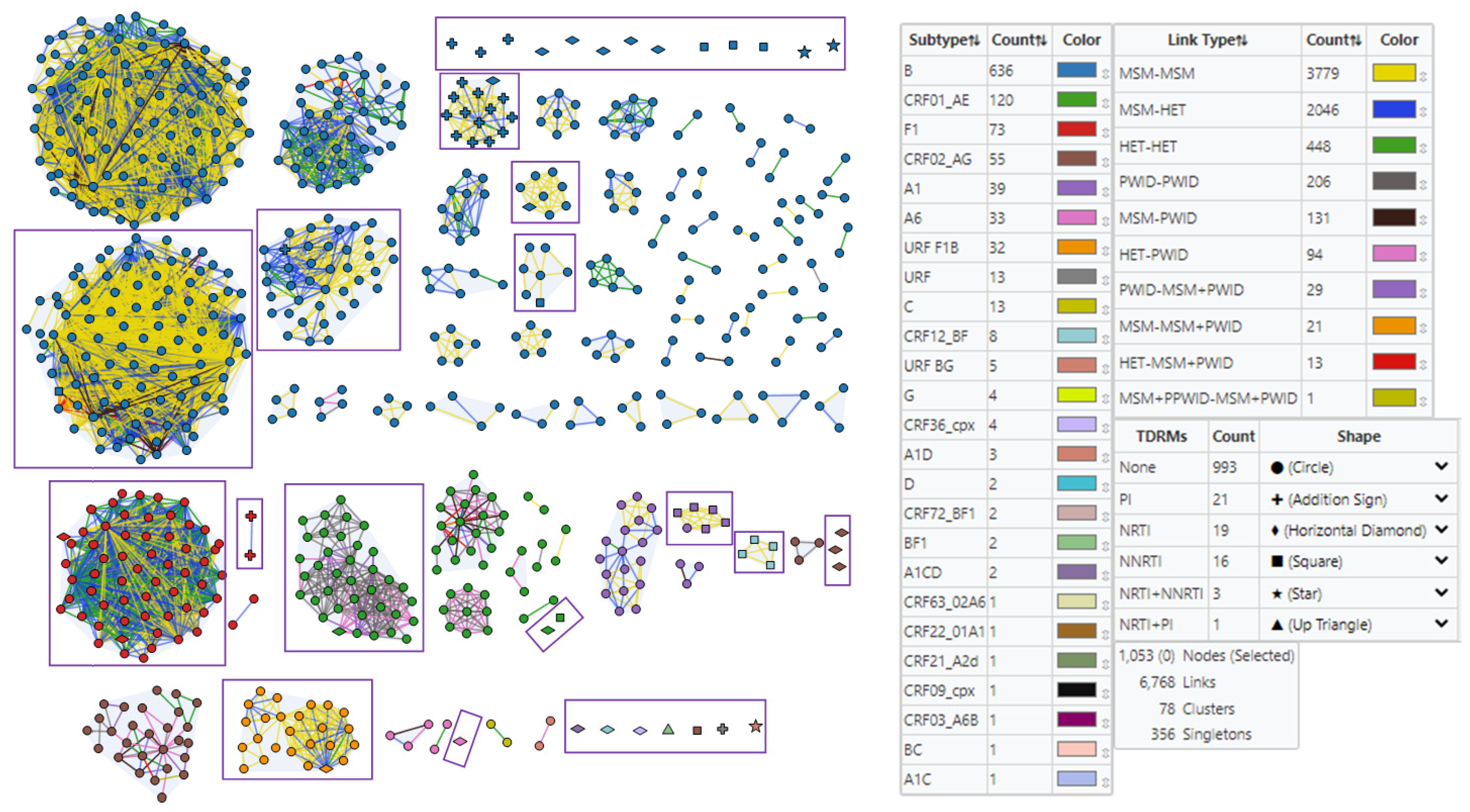
3.4. Detection of HIV-1 Transmission Clusters with SDRMs
3.4.1. Subtype and Prevalent Infection Composition of Clusters
3.4.2. Clustering of HIV-1 Sequences with SDRMs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, M.S.; McCauley, M.; Gamble, T.R. HIV treatment as prevention and HPTN 052. Curr. Opin. Hiv. Aids 2012, 7, 99–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- 90-90-90. An Ambitious Treatment Target to Help End the AIDS Epidemic. UNAIDS/JC2684. Joint United Nations Programme on HIV/AIDS (UNAIDS). 2014. Available online: https://www.unaids.org/sites/default/files/media_asset/90-90-90_en.pdf (accessed on 28 November 2022).
- HIV Drug Resistance Strategy, 2021 Update. Geneva: World Health Organization; 2021. License: CC BY-NC-SA 3.0 IGO. Available online: https://apps.who.int/iris/bitstream/handle/10665/343175/9789240030565-eng.pdf?sequence=1&isAllowed=y (accessed on 28 November 2022).
- McClung, R.P.; Atkins, A.D.; Kilkenny, M.; Bernstein, K.T.; Willenburg, K.S.; Weimer, M.; Robilotto, S.; Panneer, N.; Thomasson, E.; Adkins, E.; et al. Response to a large HIV outbreak, Cabell County, West Virginia, 2018–2019. Am. J. Prev. Med. 2021, 61 (Suppl. 1), 143–150. [Google Scholar] [CrossRef] [PubMed]
- Saag, M.S.; Gandhi, R.T.; Hoy, J.F.; Landovitz, R.J.; Thompson, M.A.; Sax, P.E.; Smith, D.M.; Benson, C.A.; Buchbinder, S.P.; Del Rio, C.; et al. Antiretroviral drugs for treatment and prevention of HIV infection in adults: 2020 recommendations of the International Antiviral Society-USA Panel. JAMA 2020, 324, 1651–1669. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.E.; Camacho, R.J.; Otelea, D.; Kuritzkes, D.R.; Fleury, H.; Kiuchi, M.; Heneine, W.; Kantor, R.; Jordan, M.R.; Schapiro, J.M.; et al. Drug resistance mutations for surveillance of transmitted HIV-1 drug-resistance: 2009 update. PLoS ONE 2009, 4, e4724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafer, R.W.; Rhee, S.Y.; Bennett, D.E. Consensus drug resistance mutations for epidemiological surveillance: Basic principles and potential controversies. Antivir. Ther. 2008, 13 (Suppl. 2), 59–68. [Google Scholar] [CrossRef] [PubMed]
- Gifford, R.J.; Rhee, S.Y.; Eriksson, N.; Liu, T.F.; Kiuchi, M.; Das, A.K.; Shafer, R.W. Sequence editing by apolipoprotein B RNA-editing catalytic component-B and epidemiological surveillance of transmitted HIV-1 Drug Resistance. AIDS 2008, 22, 717–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofstra, L.M.; Sauvageot, N.; Albert, J.; Alexiev, I.; Garcia, F.; Struck, D.; Georgiou, O. Transmission of HIV drug resistance and the predicted effect on current first-line regimens in Europe. Clin. Infect. Dis. 2016, 62, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.Y.; Kassaye, S.G.; Barrow, G.; Sundaramurthi, J.C.; Jordan, M.R.; Shafer, R.W. HIV-1 transmitted drug resistance surveillance: Shifting trends in study design and prevalence estimates. J. Int. AIDS Soc. 2020, 23, e25611. [Google Scholar] [CrossRef] [PubMed]
- Oroz, M.; Begovac, J.; Planinić, A.; Rokić, F.; Lunar, M.M.; Zorec, T.M.; Lepej, S.Ž. Analysis of HIV-1 diversity, primary drug resistance and transmission networks in Croatia. Sci. Rep. 2019, 9, 17307. [Google Scholar] [CrossRef] [Green Version]
- Kantzanou, M.; Karalexi, M.A.; Papachristou, H.; Vasilakis, A.; Rokka, C.; Katsoulidou, A. Transmitted drug resistance among HIV-1 drug-naïve patients in Greece. Int. J. Infect. Dis. 2021, 105, 42–48. [Google Scholar] [CrossRef]
- Temereanca, A.; Ene, L.; Mehta, S.; Manolescu, L.; Duiculescu, D.; Ruta, S. Transmitted HIV drug resistance in treatment-naive Romanian patients. J. Med. Virol. 2013, 85, 1139–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanojevic, M.; Siljic, M.; Salemovic, D.; Pesic-Pavlovic, I.; Zerjav, S.; Nikolic, V.; Ranin, J.; Jevtovic, D. Ten years survey of primary HIV-1 resistance in Serbia: The occurrence of multiclass resistance. AIDS Res. Hum. Retrovir. 2014, 30, 634–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunar, M.M.; Lepej, S.Ž.; Tomažič, J.; Vovko, T.D.; Pečavar, B.; Turel, G.; Maver, M.; Poljak, M. HIV-1 transmitted drug resistance in Slovenia and its impact on predicted treatment effectiveness: 2011–2016 update. PLoS ONE 2018, 13, e0196670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexiev, I.; Beshkov, D.; Shankar, A.; Hanson, D.L.; Paraskevis, D.; Georgieva, V.; Karamacheva, L.; Taskov, H.; Varleva, T.; Elenkov, I.; et al. Detailed molecular epidemiologic characterization of HIV-1 infection in Bulgaria reveals broad diversity and evolving phylodynamics. PLoS ONE 2013, 8, e59666. [Google Scholar] [CrossRef] [Green Version]
- Alexiev, I.; Shankar, A.; Wensing, A.M.J.; Beshkov, D.; Elenkov, I.; Stoycheva, M.; Switzer, W.M. Low HIV-1 transmitted drug resistance in Bulgaria against a background of high clade diversity. J. Antimicrob. Chemother. 2015, 70, 1874–1880. [Google Scholar] [CrossRef] [Green Version]
- WHO. Prevalence of HIV Among Adults Aged 15 to 49 (%). Available online: https://www.who.int/data/gho/data/indicators/indicator-details/GHO/prevalence-of-hiv-among-adults-aged-15-to-49-(-) (accessed on 27 March 2023).
- Alexiev, I.; Mavian, C.; Paisie, T.; Ciccozzi, M.; Dimitrova, R.; Gancheva, A.; Kostadinova, A.; Seguin-Devaux, C.; Salemi, M. Analysis of the origin and dissemination of HIV-1 subtype C in Bulgaria. Viruses 2022, 14, 263. [Google Scholar] [CrossRef]
- Alexiev, I.; Shankar, A.; Dimitrova, R.; Gancheva, A.; Kostadinova, A.; Teoharov, P.; Golkocheva, E.; Nikolova, M.; Muhtarova, M.; Elenkov, I.; et al. Origin and spread of HIV-1 in persons who inject drugs in Bulgaria. Infect. Genet. Evol. 2016, 46, 269–278. [Google Scholar] [CrossRef]
- Alexiev, I.; Lo Presti, A.; Dimitrova, R.; Foley, B.; Gancheva, A.; Kostadinova, A.; Nikolova, L.; Angeletti, S.; Cella, E.; Elenkov, I.; et al. Origin and spread of HIV-1 subtype B among heterosexual individuals in Bulgaria. AIDS Res. Hum. Retrovir. 2018, 34, 244–253. [Google Scholar] [CrossRef]
- Alexiev, I.; Campbell, E.; Knyazev, S.; Pan, Y.; Grigorova, L.; Dimitrova, R.; Partsuneva, A.; Gancheva, A.; Kostadinova, A.; Seguin-Devaux, C.; et al. Molecular epidemiology of the HIV-1 Subtype B sub-epidemic in Bulgaria. Viruses 2020, 12, 441. [Google Scholar] [CrossRef] [Green Version]
- Alexiev, I.; Campbell, E.M.; Knyazev, S.; Pan, Y.; Grigorova, L.; Dimitrova, R.; Partsuneva, A.; Gancheva, A.; Kostadinova, A.; Seguin-Devaux, C. Molecular eEpidemiological analysis of the origin and transmission dynamics of the HIV-1 CRF01_AE sub-epidemic in Bulgaria. Viruses 2021, 13, 116. [Google Scholar] [CrossRef]
- Struck, D.; Lawyer, G.; Ternes, A.M.; Schmit, J.C.; Bercoff, D.P. COMET: Adaptive context-based modeling for ultrafast HIV-1 subtype identification. Nucleic Acids Res. 2014, 42, e144. [Google Scholar] [CrossRef] [PubMed]
- Peña, A.C.P.; Faria, N.R.; Imbrechts, S.; Libin, P.; Abecasis, A.B.; Deforche, K.; Gomez, A.; Camacho, R.J.; de Oliveira, T.; Vandamme, A.M. Automated subtyping of HIV-1 genetic sequences for clinical and surveillance purposes: Performance evaluation of the new REGA version 3 and seven other tools. Infect. Genet. Evol. 2013, 19, 337–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, A.K.; Zhang, M.; Bulla, I.; Leitner, T.; Korber, B.; Morgenstern, B.; Stanke, M. jpHMM: Improving the reliability of recombination prediction in HIV-1. Nucleic Acids Res. 2009, 37, 647–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuraku, S.; Zmasek, C.M.; Nishimura, O.; Katoh, K. aLeaves facilitates on-demand exploration of metazoan gene family trees on MAFFT sequence alignment server with enhanced interactivity. Nucleic Acids Res. 2013, 41, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Paramvir, S.D.; Adam, P.A. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Los Alamos National Laboratory Database. Available online: https://www.hiv.lanl.gov/content/index (accessed on 21 March 2022).
- FigTree Version–v1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 21 March 2022).
- HIV Drug Resistance Database. Available online: https://hivdb.stanford.edu/cpr/form/PRRT/ (accessed on 21 March 2022).
- Campbell, E.M.; Boyles, A.; Shankar, A.; Kim, J.; Knyazev, S.; Cintron, R.; Switzer, W.M. MicrobeTrace: Retooling molecular epidemiology for rapid public health response. PLoS Comput. Biol. 2021, 17, e1009300. [Google Scholar] [CrossRef]
- Miranda, M.N.S.; Pingarilho, M.; Pimentel, V.; Martins, M.D.R.O.; Kaiser, R.; Seguin-Devaux, C.; Paredes, R.; Zazzi, M.; Incardona, F.; Abecasis, A.B. Trends of Transmitted and Acquired Drug Resistance in Europe from 1981 to 2019: A Comparison Between the Populations of Late Presenters and Non-late Presenters. Front. Microbiol. 2022, 13, 846943. [Google Scholar] [CrossRef]
- EACS Guidelines Version 9.0 October 2017. Available online: https://www.eacsociety.org/media/guidelines_9.0-english.pdf (accessed on 27 March 2023).
- Tang, M.W.; Rhee, S.Y.; Bertagnolio, S.; Ford, N.; Holmes, S.; Sigaloff, K.C.; Hamers, R.L.; Wit, T.F.R.; Fleury, H.J.; Kanki, P.J.; et al. Nucleoside reverse transcriptase inhibitor resistance mutations associated with first-line stavudine-containing antiretroviral therapy: Programmatic implications for countries phasing out stavudine. J. Infect. Dis. 2013, 207 (Suppl. 2), S70–S77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knox, D.C.; Anderson, P.L.; Harrigan, P.R.; Tan, D.H. Multidrug-resistant HIV-1 infection despite preexposure prophylaxis. N. Engl. J. Med. 2017, 376, 501–502. [Google Scholar] [CrossRef] [PubMed]
- Guan-Jhou, C.; Yu-Lin, L.; Chen-Hsiang, L.; Hsin-Yun, S.; Chien-Yu, C.; Hung-Chin, T.; Sung, H.H.; Yi-Chieh, L.; Min-Han, H.; Sui-Yuan, C.; et al. Impact of archived M184V/I mutation on the effectiveness of switch to co-formulated elvitegravir, cobicistat, emtricitabine and tenofovir alafenamide among virally suppressed people living with HIV. J. Antimicrob. Chemother. 2020, 75, 2986–2993. [Google Scholar] [CrossRef]
- Podlekareva, D.; Grint, D.; Karpov, I.; Rakmanova, A.; Mansinho, K.; Chentsova, N.; Zeltina, I.; Losso, M.; Parczewski, M.; Lundgren, J.; et al. Time trends in d4T utilization in Europe. HIV Med. 2015, 16, 533–543. [Google Scholar] [CrossRef]
- Naugler, W.E.; Yong, F.H.; Carey, V.J.; Dragavon, J.A.; Coombs, R.W.; Frenkel, L.M. T69D/N pol mutation, human immunodeficiency virus type 1 RNA levels, and syncytium-inducing phenotype are associated with CD4 cell depletion during didanosine therapy. J. Infect. Dis. 2002, 185, 448–455. [Google Scholar] [CrossRef]
- Bacheler, L.T.; Anton, E.D.; Kudish, P.; Baker, D.; Bunville, J.; Krakowski, K.; Bolling, L.; Aujay, M.; Wang, X.V.; Ellis, D.; et al. Human immunodeficiency virus type 1 mutations selected in patients failing efavirenz combination therapy. Antimicrob. Agents Chemother. 2000, 44, 2475–2484. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.Y.; Tzou, P.L.; Shafer, R.W. Temporal trends in HIV-1 mutations used for the surveillance of transmitted drug resistance. Viruses 2021, 13, 879. [Google Scholar] [CrossRef]
- Bertagnolio, S.; Hermans, L.; Jordan, M.R.; Avila-Rios, S.; Iwuji, C.; Derache, A.; Delaporte, E.; Wensing, A.; Aves, T.; Borhan, A.S.M.; et al. Clinical impact of pretreatment human immunodeficiency virus drug resistance in people initiating nonnucleoside reverse transcriptase inhibitor–containing antiretroviral therapy: A systematic review and meta-analysis. J. Infect. Dis. 2020, 224, 377–388. [Google Scholar] [CrossRef]
- Wertheim, J.O.; Oster, A.M.; Johnson, J.A.; Switzer, W.M.; Saduvala, N.; Hernandez, A.L.; Hall, H.I.; Heneine, W. Transmission fitness of drug-resistant HIV revealed in a surveillance system transmission network. Virus Evol. 2017, 3, vex008. [Google Scholar] [CrossRef] [Green Version]
- EACS Guidelines, Version 11.0—October 2021. Available online: https://www.eacsociety.org/guidelines/eacs-guidelines/ (accessed on 28 November 2022).
- Turner, D.; Amit, S.; Chalom, S.; Penn, O.; Pupko, T.; Katchman, E.; Matus, N.; Tellio, H.; Katzir, M.; Avidor, B. Emergence of an HIV-1 cluster harbouring the major protease L90M mutation among treatment-naive patients in Tel Aviv, Israel. HIV Med. 2012, 13, 202–206. [Google Scholar] [CrossRef]
- Brenner, B.G.; Roger, M.; Moisi, D.D.; Oliveira, M.; Hardy, I.; Turgel, R.; Charest, H.; Routy, J.P.; Wainberg, M.A.; Montreal PHI Cohort and HIV Prevention Study Groups. Transmission networks of drug resistance acquired in primary/early stage HIV infection. AIDS 2008, 22, 2509–2515. [Google Scholar] [CrossRef] [PubMed]
- Sevin, A.D.; DeGruttola, V.; Nijhuis, M.; Schapiro, J.M.; Foulkes, A.S.; Para, M.F.; Boucher, C.A. Methods for investigation of the relationship between drug-susceptibility phenotype and human immunodeficiency virus type 1 genotype with applications to AIDS clinical trials group 333. J. Infect. Dis. 2000, 182, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Stanojevic, M.; Alexiev, I.; Beshkov, D.; Gökengin, D.; Mezei, M.; Minarovits, J.; Otelea, D.; Paraschiv, S.; Poljak, M.; Zidovec-Lepej, S.; et al. HIV-1 molecular epidemiology in the Balkans: A melting pot for high genetic diversity. AIDS Rev. 2012, 14, 28–36. [Google Scholar] [PubMed]
- Salemi, M.; De Oliveira, T.; Ciccozzi, M.; Rezza, G.; Goodenow, M.M. High-resolution molecular epidemiology and evolutionary history of HIV-1 subtypes in Albania. PLoS ONE 2008, 3, e1390. [Google Scholar] [CrossRef] [Green Version]
- Niculescu, I.; Paraschiv, S.; Paraskevis, D.; Abagiu, A.; Batan, I.; Banica, L.; Otelea, D. Recent HIV-1 outbreak among intravenous drug users in Romania: Evidence for cocirculation of CRF14_BG and subtype F1 strains. AIDS Res. Hum. Retrovir. 2015, 31, 488–495. [Google Scholar] [CrossRef] [Green Version]
- Abidi, S.H.; Aibekova, L.; Davlidova, S.; Amangeldiyeva, A.; Foley, B.; Ali, S. Origin and evolution of HIV-1 subtype A6. PLoS ONE 2021, 16, e0260604. [Google Scholar] [CrossRef]
- Jovanovic, L.; Siljic, M.; Cirkovic, V.; Salemovic, D.; Jevtovic, D.; Alexiev, I.; Zidovec-Lepej, S.; Oroz, M.; Begovac, J.; Paraskevis, D.; et al. HIV-1 Subtype B spread through cross-border clusters in the Balkans: A molecular analysis in view of incidence trends. AIDS 2022, 10, 1097. [Google Scholar] [CrossRef]
- Alexiev, I.; Dimitrova, R.; Gancheva, A.; Kostadinova, A.; Grigorova, L.; Partsuneva, A.; Nikolova, M. Molecular epidemiological analysis of the transmission clusters of the HIV-1 circulating recombinant forms CRF01_AE and CRF02_AG in Bulgaria. Probl. Infect. Parasit. Dis. 2020, 48, 25–32. [Google Scholar] [CrossRef]
- Kouyos, R.D.; von Wyl, V.; Yerly, S.; Böni, J.; Taffé, P.; Shah, C.; Bürgisser, P.; Klimkait, T.; Weber, R.; Hirschel, B.; et al. Molecular epidemiology reveals long-term changes in HIV type 1 subtype B transmission in Switzerland. J. Infect. Dis. 2010, 201, 1488–1497. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristic | HIV-1 Infections with SDRMs (n = 60) | HIV-1 Infections without SDRMs (n = 993) | p 1 |
|---|---|---|---|
| Gender | 0.446 | ||
| Male | 54 (90.0%) | 851 (85.7%) | |
| Female | 6 (10.0%) | 142 (14.3%) | |
| Likely route of HIV infection 2 | 0.242 | ||
| HET | 22 (36.6%) | 379 (38.2%) | |
| MMSC | 36 (60.0%) | 498 (50.2%) | |
| PWID | 2 (3.3%) | 99 (10.0%) | |
| MMSC + PWID | 0 (0.0%) | 17 (1.7%) | |
| Likely country of infection | 0.596 | ||
| Bulgaria | 48 (80.0%) | 824 (82.9%) | |
| Abroad | 12 (20.0%) | 169 (17.0%) | |
| Migrants 3 | 0.239 | ||
| Yes | 15 (25.0%) | 187(18.8%) | |
| No | 45 (75.0%) | 806 (81.2%) | |
| Individuals incarcerated | 0.717 | ||
| Yes | 1 (1.7%) | 34 (3.4%) | |
| No | 59 (98.3%) | 959 (96.6%) | |
| Blood donors | 0.043 | ||
| Yes | 0 (0.0%) | 56 (5.6%) | |
| No | 60 (100.0%) | 937 (94.4%) | |
| Sex workers | 0.537 | ||
| Yes | 2 (3.3%) | 29 (2.9%) | |
| No | 58 (96.7%) | 964 (97.1%) | |
| Pregnant Women | 0.643 | ||
| Yes | 2 (3.3%) | 22 (2.4%) | |
| No | 58 (96.7%) | 971 (97.8%) | |
| STI 4 | 0.926 | ||
| Yes | 15 (25.0%) | 243 (24.5%) | |
| No | 45 (75.0%) | 750 (75.5%) | |
| Subtype 5 | <0.0001 | ||
| B | 32 (53.3%) | 604 (60.8%) | |
| F1 | 4 (6.7%) | 69 (6.9%) | |
| CRF02_AG | 4 (6.7%) | 51 (5.1%) | |
| A1 | 5 (8.3%) | 34 (3.4%) | |
| CRF12_BF | 5 (8.3%) | 3 (0.3%) | |
| Other | 10 (16.7%) | 232 (23.4%) | |
| HIV-1 Subtype | HIV-1 Subtype (%) | Women (%) | Men (%) | Bulgarians Infected in Bulgaria (%) | Bulgarians Infected Abroad (%) | Foreigners Diagnosed in Bulgaria (%) | HET 1 (%) | MMSC 2 (%) | PWID 3 (%) | MMSC + PWID 4 (%) | Any SDRMs (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | 39 (3.7) | 3 (2) | 36 (4.0) | 32 (3.8) | 5 (3.3) | 2 (4) | 9 (2.2) | 28 (5.2) | 2 (2) | 5 (8.3) | |
| A1C | 1 (0.1) | 1 (0.7) | 1 (0.1) | 1 (0.2) | |||||||
| A1CD | 2 (0.2) | 1 (0.7) | 1 (0.1) | 1 (0.1) | 1 (0.7) | 2 (0.5) | 1 (1.7) | ||||
| A1D | 3 (0.3) | 2 (1.4) | 1 (0.1) | 1 (0.1) | 2 (4) | 2 (0.5) | 1 (0.2) | 1 (1.7) | |||
| A6 | 33 (3.1) | 12 (8.1) | 21 (2.3) | 19 (2.2) | 7 (4.6) | 7 (14) | 24 (6) | 8 (1.5) | 1 (1) | 1 (1.7) | |
| B | 636 (60.4) | 49 (33.1) | 587 (64.9) | 533 (62.6) | 88 (57.9) | 15 (30) | 213 (53.1) | 404 (75.7) | 15 (14.9) | 4 (23.5) | 32 (53.3) |
| BC | 1 (0.1) | 1 (0.1) | 1 (0.7) | 1 (0.2) | |||||||
| BF1 | 2 (0.2) | 2 (0.2) | 1 (0.1) | 1 (0.7) | 2 (0.5) | 1 (1.7) | |||||
| C | 13 (1.2) | 9 (6.1) | 4 (0.4) | 8 (0.9) | 3 (2) | 2 (4) | 12 (3) | 1 (5.9) | |||
| CRF01_AE | 120 (11.4) | 32 (21.6) | 88 (9.7) | 101 (11.9) | 17 (11.2) | 2 (4) | 40 (10) | 10 (1.9) | 64 (63.4) | 6 (35.3) | 3 (5) |
| CRF02_AG | 55 (5.2) | 17 (11.5) | 38 (4.2) | 43 (5.1) | 6 (3.9) | 6 (12) | 30 (7.5) | 6 (1.1) | 13 (12.9) | 6 (35.3) | 4 (6.7) |
| CRF03_A6B | 1 (0.1) | 1 (0.1) | 1 (0.1) | 1 (1) | |||||||
| CRF09_cpx | 1 (0.1) | 1 (0.1) | 1 (2) | 1 (0.2) | |||||||
| CRF12_BF | 8 (0.8) | 1 (0.7) | 7 (0.8) | 6 (0.7) | 1 (0.7) | 1 (2) | 2 (0.5) | 6 (1.1) | 5 (8.3) | ||
| CRF21_A2D | 1 (0.1) | 1 (0.1) | 1 (0.1) | 1 (1) | |||||||
| CRF22_01A1 | 1 (0.1) | 1 (0.7) | 1 (2) | 1 (0.2) | |||||||
| CRF36_cpx | 4 (0.4) | 1 (0.7) | 3 (0.3) | 3 (0.4) | 1 (2) | 3 (0.7) | 1 (1) | 1 (1.7) | |||
| CRF63_02A6 | 1 (0.1) | 1 (0.1) | 1 (0.1) | 1 (0.2) | |||||||
| CRF72_BF1 | 2 (0.2) | 2 (0.2) | 2 (0.2) | 2 (0.5) | |||||||
| D | 2 (0.2) | 1 (0.7) | 1 (0.1) | 2 (4) | 2 (0.5) | ||||||
| F1 | 73 (6.9) | 12 (8.1) | 61 (6.7) | 60 (7.1) | 10 (6.6) | 3 (6) | 37 (9.2) | 35 (6.6) | 1 (1) | 4 (6.7) | |
| G | 4 (0.4) | 2 (1.4) | 2 (0.2) | 2 (1.3) | 2 (4) | 3 (0.7) | 1 (0.2) | ||||
| URFs 5 | 50 (4.7) | 4 (2.7) | 46 (5.1) | 37 (4.3) | 10 (6.6) | 3 (6) | 14 (3.5) | 34 (6.4) | 2 (2) | 2 (3.3) | |
| Total | 1053 | 148 (14.1) | 905 (85.9) | 851 (80.8) | 152 (14.4) | 50 (4.7) | 401 (38.1) | 534 (50.7) | 101 (9.6) | 17 (1.6) | 60 (5.7) |
| Characteristic 1 | Sequences with SDRMs in Clusters (n = 34) | Sequences without SDRMs in Clusters (n = 663) | p Value 2 | Non-Clustering Sequences with SDRMs (n = 26) | Non-Clustering Sequences without SDRMS (n = 330) | p Value 3 |
|---|---|---|---|---|---|---|
| Gender | 0.104 | 1.0 | ||||
| Male | 34 (100%) | 604 (91.1%) | 20 (76.9%) | 247 (75.9%) | ||
| Female | 0 (0%) | 59 (8.9%) | 6 (23.1%) | 83 (25.1%) | ||
| Likely route of HIV infection | 0.442 | 0.846 | ||||
| HET | 6 (17.6%) | 190 (28.7%) | 17 (65.4%) | 189 (57.3%) | ||
| MMSC | 28 (82.4%) | 384 (57.9%) | 8 (30.8%) | 114 (34.5%) | ||
| PWID | 0 (0%) | 80 (12.1%) | 2 (7.7%) | 19 (5.8%) | ||
| MMSC + PWID | 0 (0%) | 9 (1.4%) | 0 (0%) | 8 (2.4%) | ||
| Subtype | <0.0001 | 0.250 | ||||
| B | 19 (55.9%) | 444 (83.7%) | 13 (50.0%) | 160 (48.5%) | ||
| F1 | 4 (11.8%) | 59 (8.9%) | 0 (0%) | 10 (3.0%) | ||
| CRF02_AG | 0 (0%) | 30 (4.5%) | 4 (15.4%) | 21 (6.4%) | ||
| A1 | 5 (14.7%) | 22 (3.3%) | 0 (0%) | 12 (3.6%) | ||
| CRF12_BF | 4 (11.8%) | 0 (0%) | 1 (3.8%) | 3 (0.9%) | ||
| Other | 2 (5.9%) | 108 (16.3%) | 8 (30.8%) | 124 (37.6%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexiev, I.; Shankar, A.; Pan, Y.; Grigorova, L.; Partsuneva, A.; Dimitrova, R.; Gancheva, A.; Kostadinova, A.; Elenkov, I.; Yancheva, N.; et al. Transmitted HIV Drug Resistance in Bulgaria Occurs in Clusters of Individuals from Different Transmission Groups and Various Subtypes (2012–2020). Viruses 2023, 15, 941. https://doi.org/10.3390/v15040941
Alexiev I, Shankar A, Pan Y, Grigorova L, Partsuneva A, Dimitrova R, Gancheva A, Kostadinova A, Elenkov I, Yancheva N, et al. Transmitted HIV Drug Resistance in Bulgaria Occurs in Clusters of Individuals from Different Transmission Groups and Various Subtypes (2012–2020). Viruses. 2023; 15(4):941. https://doi.org/10.3390/v15040941
Chicago/Turabian StyleAlexiev, Ivailo, Anupama Shankar, Yi Pan, Lyubomira Grigorova, Alexandra Partsuneva, Reneta Dimitrova, Anna Gancheva, Asya Kostadinova, Ivaylo Elenkov, Nina Yancheva, and et al. 2023. "Transmitted HIV Drug Resistance in Bulgaria Occurs in Clusters of Individuals from Different Transmission Groups and Various Subtypes (2012–2020)" Viruses 15, no. 4: 941. https://doi.org/10.3390/v15040941
APA StyleAlexiev, I., Shankar, A., Pan, Y., Grigorova, L., Partsuneva, A., Dimitrova, R., Gancheva, A., Kostadinova, A., Elenkov, I., Yancheva, N., Grozdeva, R., Strashimirov, D., Stoycheva, M., Baltadzhiev, I., Doichinova, T., Pekova, L., Kosmidis, M., Emilova, R., Nikolova, M., & Switzer, W. M. (2023). Transmitted HIV Drug Resistance in Bulgaria Occurs in Clusters of Individuals from Different Transmission Groups and Various Subtypes (2012–2020). Viruses, 15(4), 941. https://doi.org/10.3390/v15040941