


Development of a Multi-Epitope Universal mRNA Vaccine Candidate for Monkeypox, Smallpox, and Vaccinia Viruses: Design and In Silico Analyses

 , , ,
, , ,
Abstract
:1. Introduction
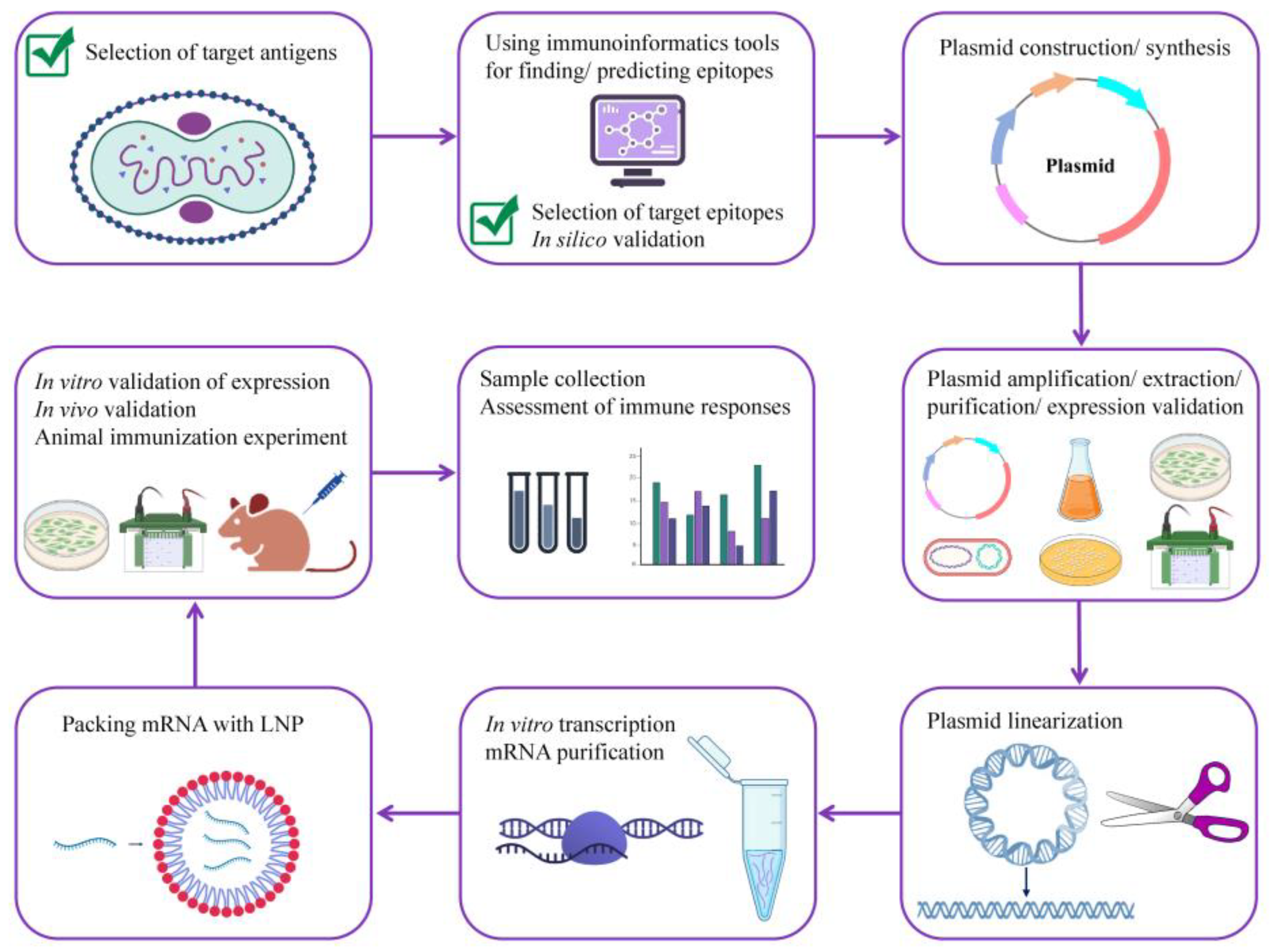
2. Methods
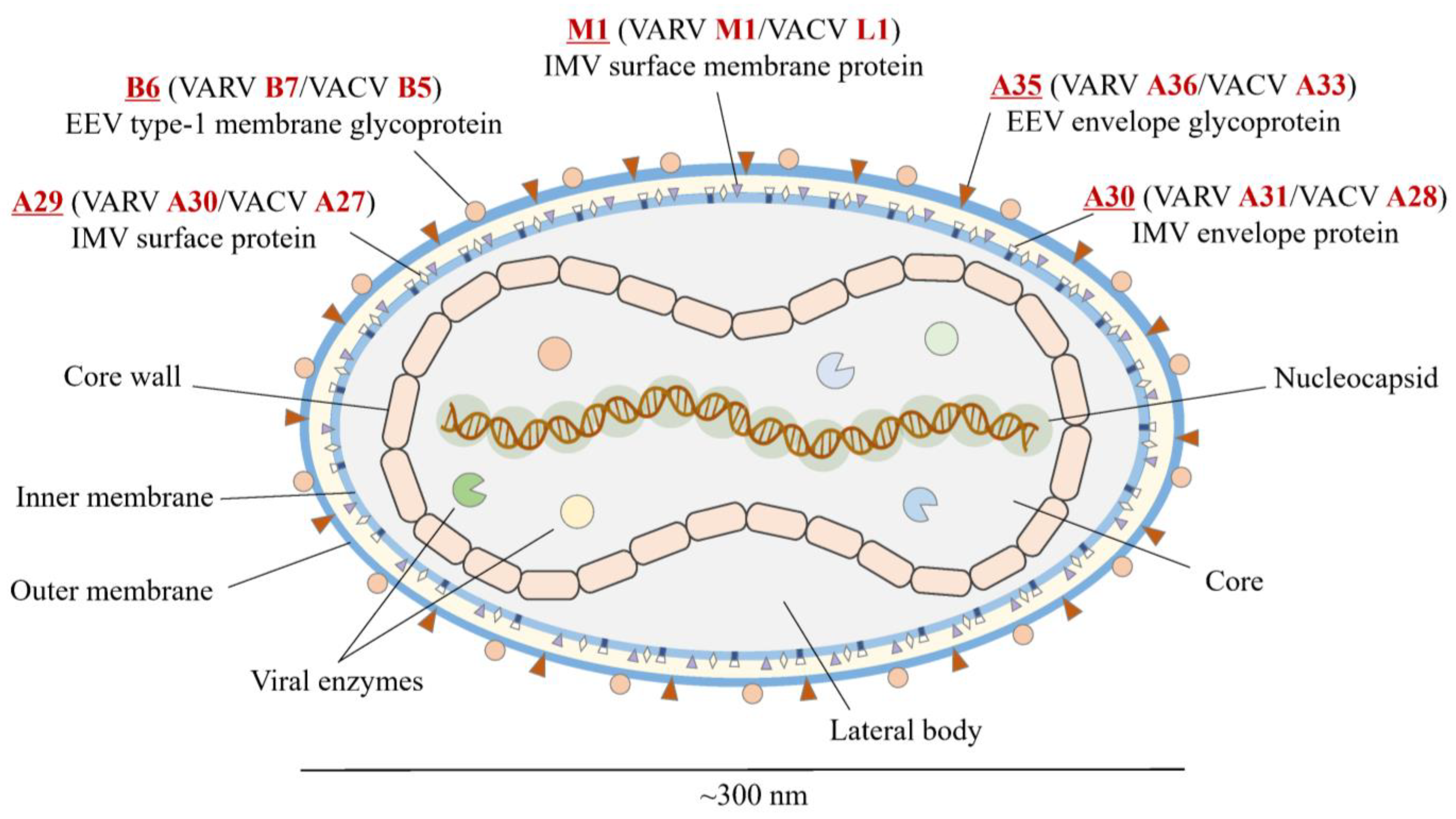
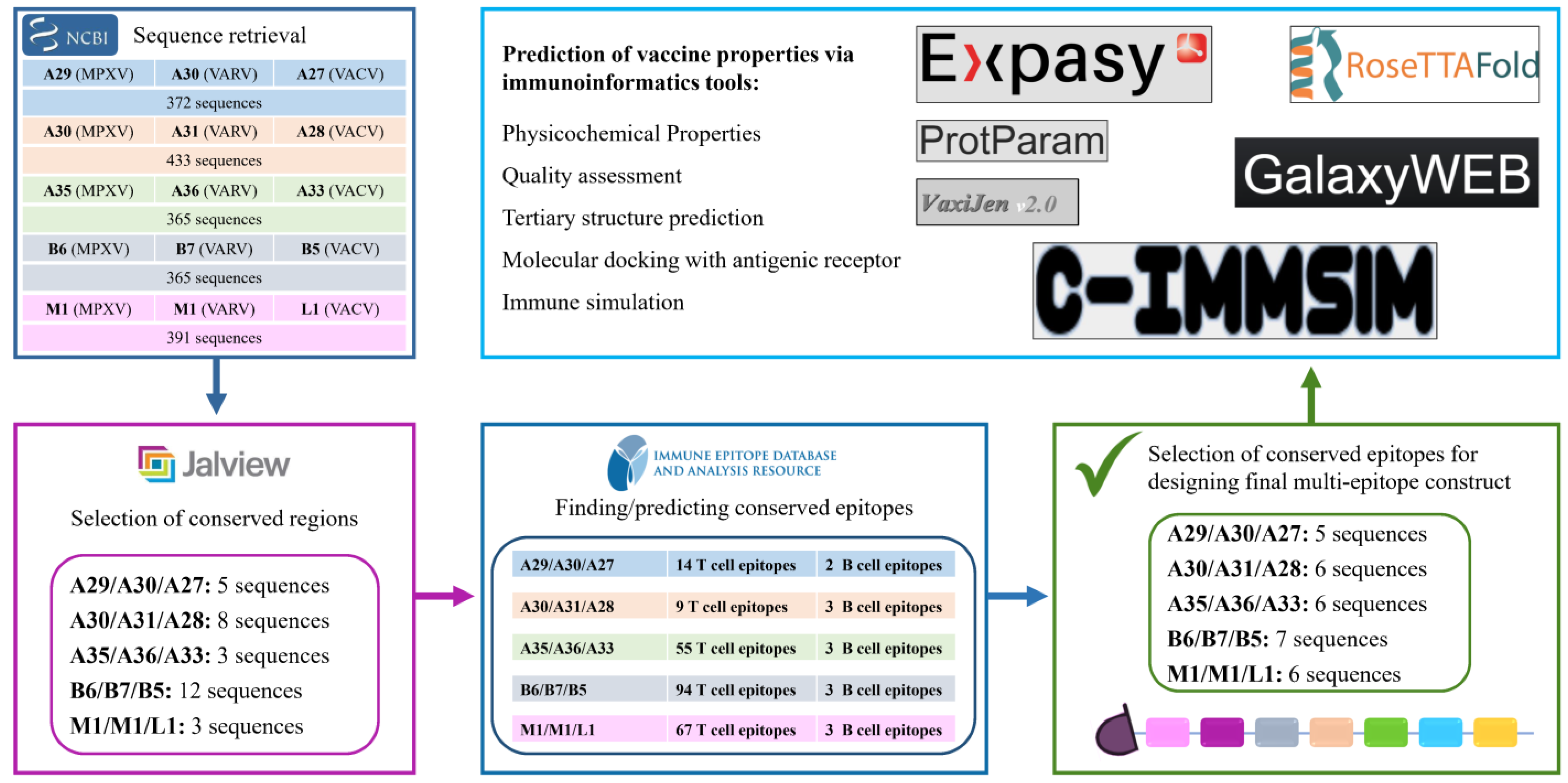
2.1. Antigen Selection
2.2. Selection of Conserved Regions and Epitopes
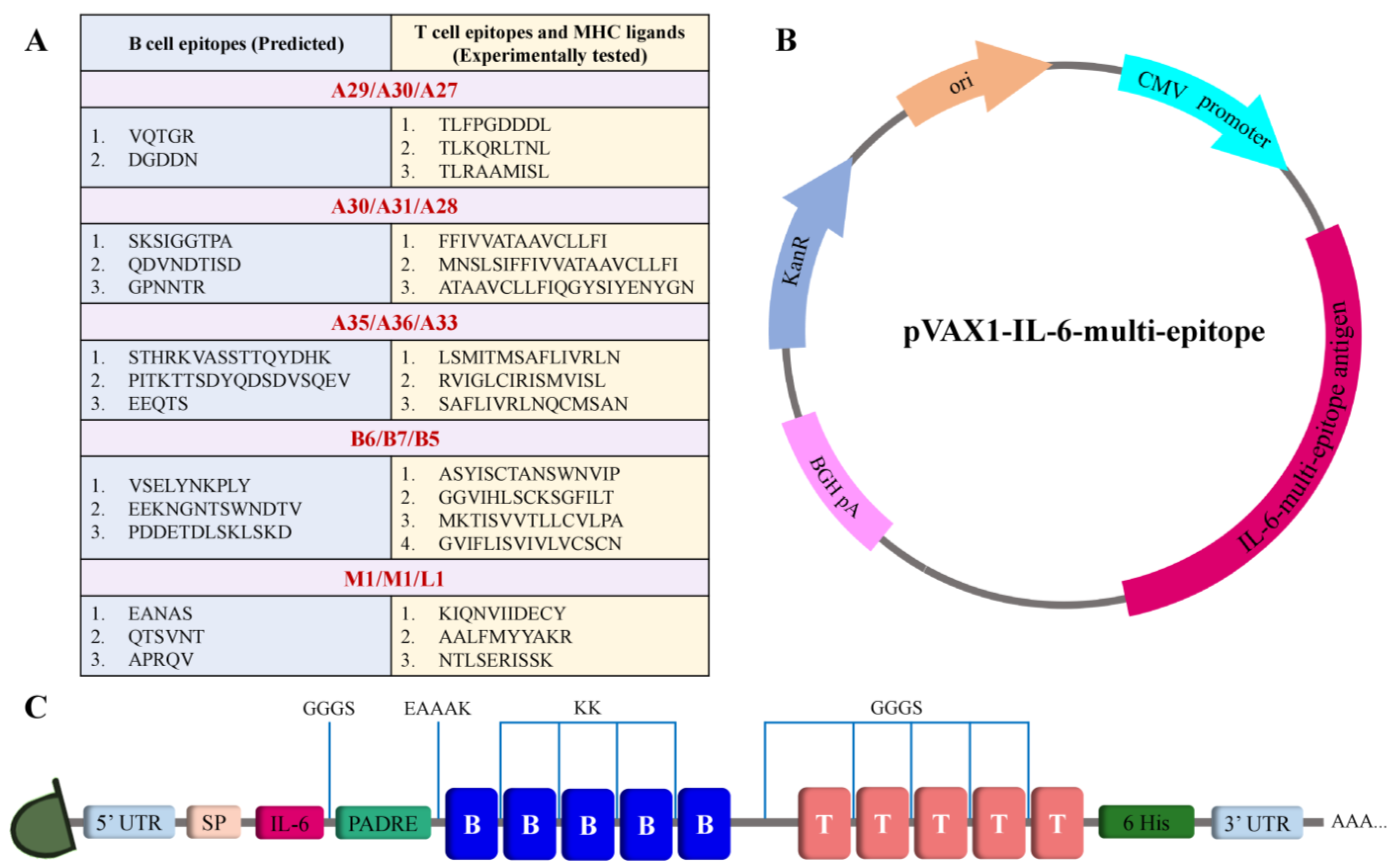
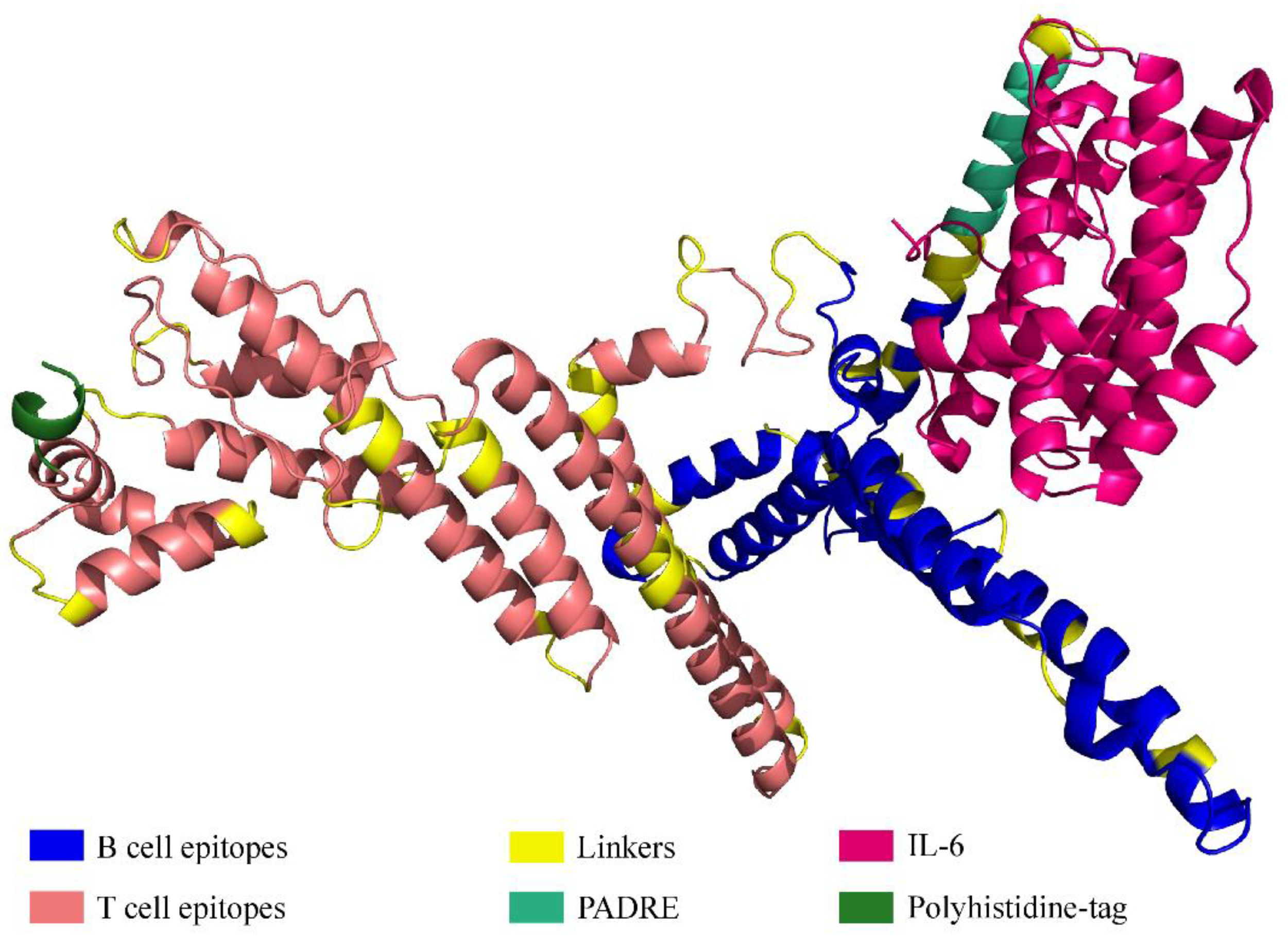
2.3. Vaccine Design
2.4. Prediction of Vaccine Properties
2.5. Tertiary Structure Prediction and Evaluation of Its Quality; Discontinuous B Cell Epitope Prediction
2.6. Immune Simulation
2.7. In Silico Validation of Vaccine Protein Binding to the Host Receptors
3. Results
3.1. Selecting Conserved Epitopes
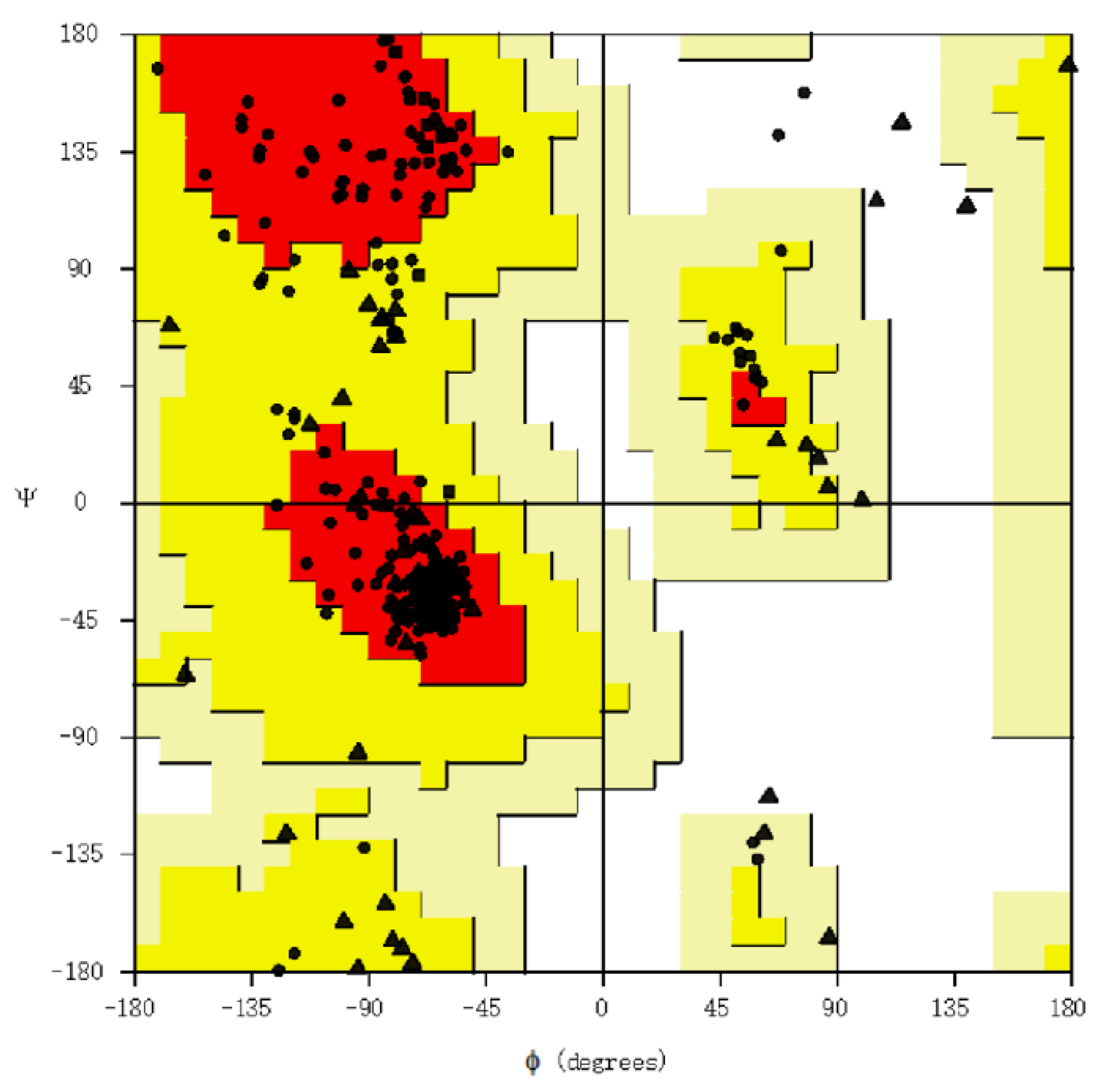
3.2. Assessment of Structure
3.3. Physicochemical Properties, Allergenicity, and Antigenicity Analyses
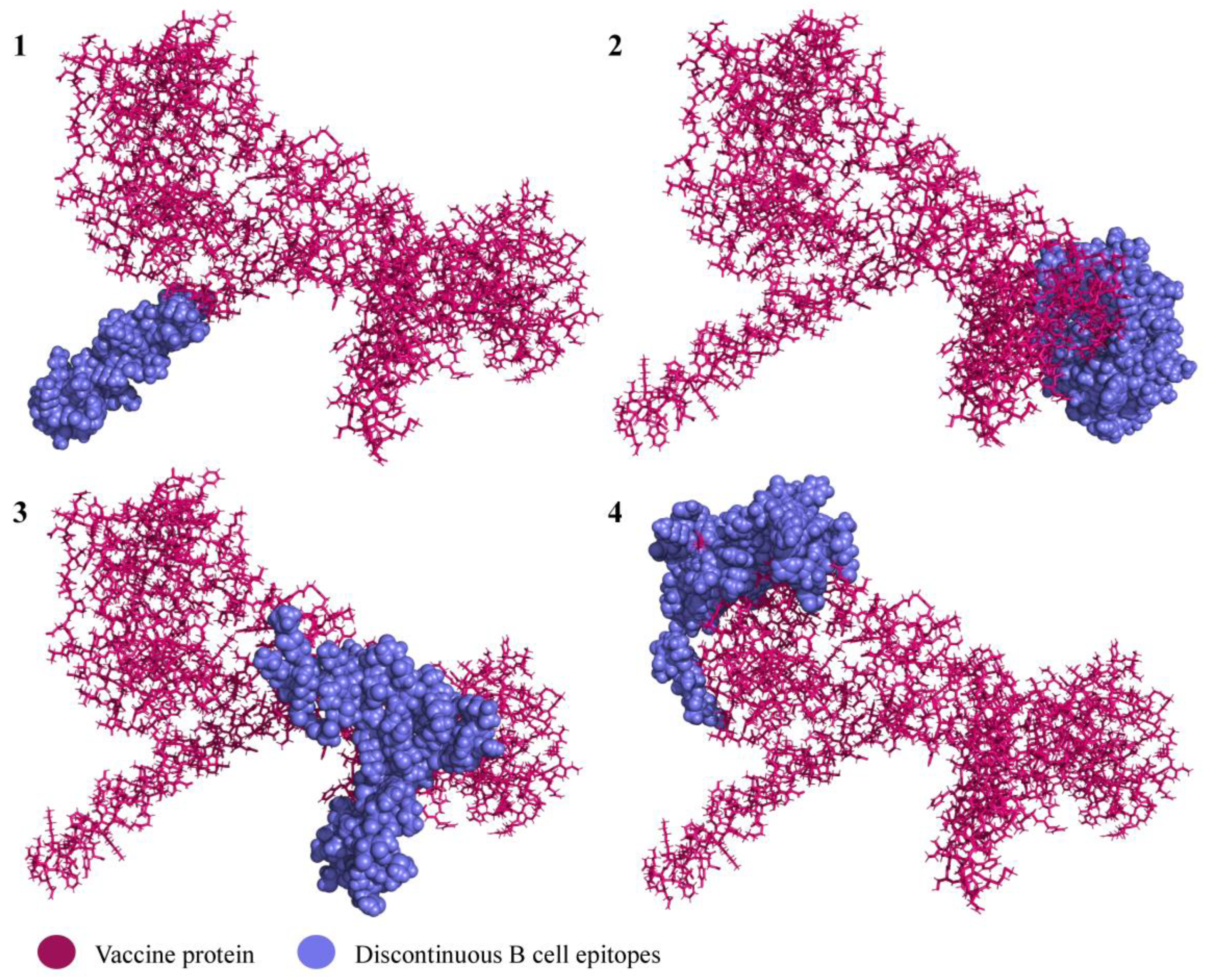
3.4. Conformational B Cell Epitopes
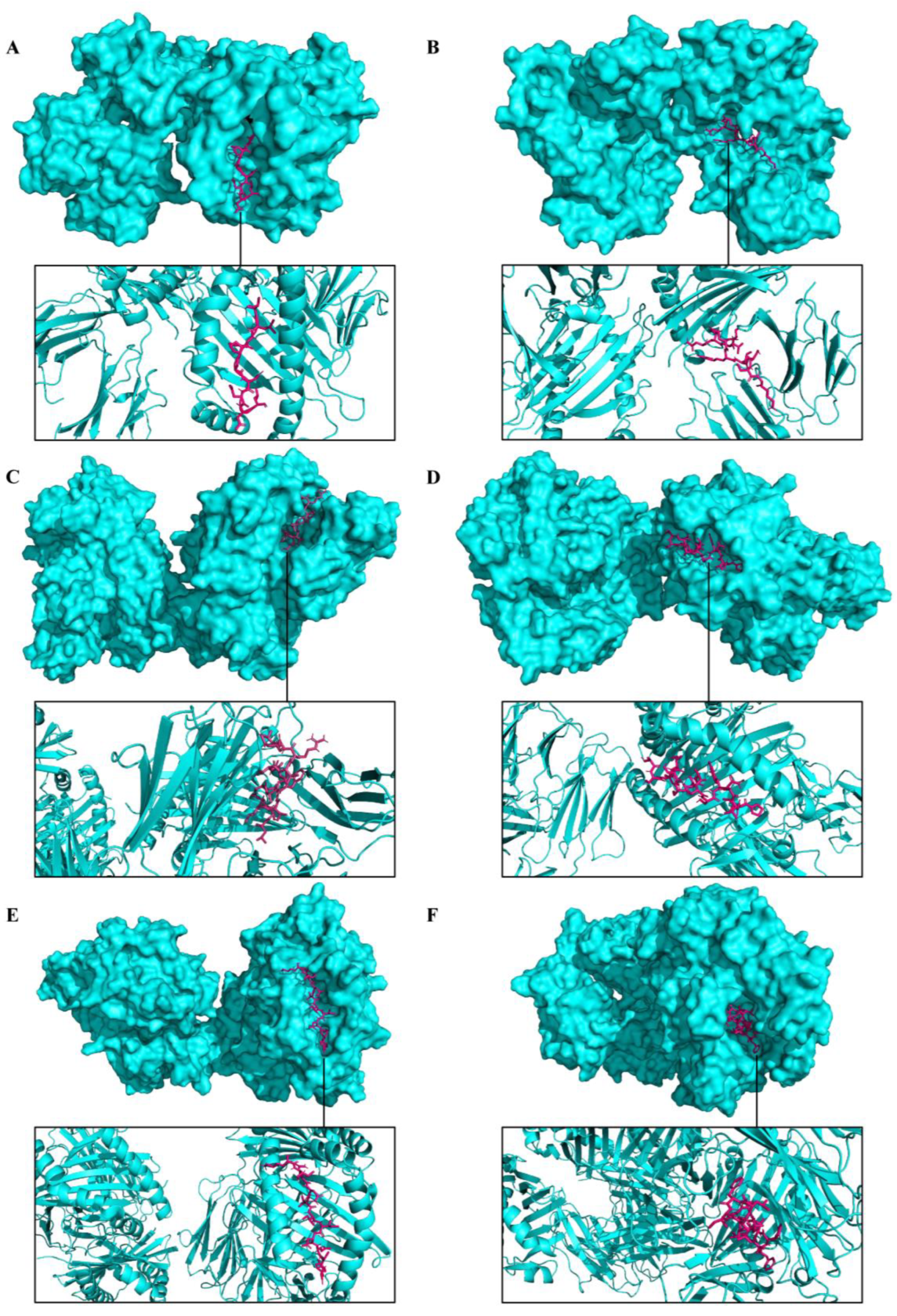
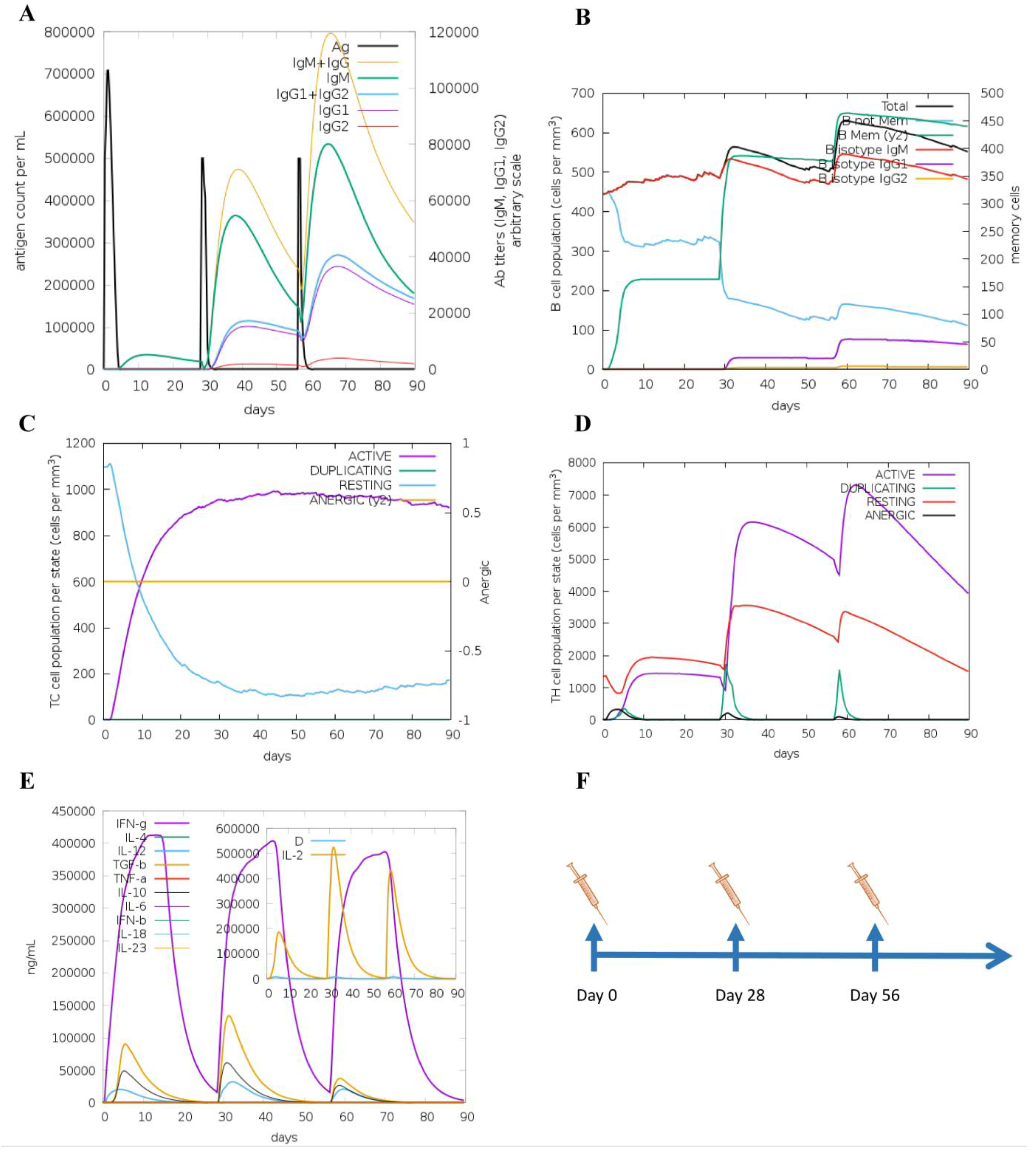
3.5. Molecular Docking and Immune Responses following Immune Simulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Centers for Disease Control and Prevention (CDC). 2022 Monkeypox Outbreak Global Map. Available online: https://www.cdc.gov/poxvirus/monkeypox/response/2022/world-map.html (accessed on 6 March 2023).
- Vandenbogaert, M.; Kwasiborski, A.; Gonofio, E.; Descorps, S.; Selekon, B.; Andy, A.; Meyong, N.; Ouilibona, R.S.; Gessain, A.; Manuguerra, J.C.; et al. Nanopore Sequencing of a Monkeypox Virus Strain Isolated from a Pustular Lesion in the Central African Republic. Sci. Rep. 2022, 12, 10768. [Google Scholar] [CrossRef] [PubMed]
- Shchelkunov, S.N.; Totmenin, A.V.; Safronov, P.F.; Mikheev, M.V.; Gutorov, V.V.; Ryazankina, O.I.; Petrov, N.A.; Babkin, I.V.; Uvarova, E.A.; Sandakhchiev, L.S.; et al. Analysis of the Monkeypox Virus Genome. Virology 2002, 197, 172–194. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Ji, J.; Shi, D.; Lu, X.; Wang, B.; Wu, N.; Wu, J. Unusual Global Outbreak of Monkeypox: What Should We Do? Front. Med. 2022, 16, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Past U.S. Cases and Outbreaks. Available online: https://www.cdc.gov/poxvirus/monkeypox/outbreak/us-outbreaks.html (accessed on 16 September 2022).
- Gong, Q.; Wang, C.; Chuai, X.; Chiu, S. Monkeypox Virus: A Re-Emergent Threat to Humans. Virol. Sin. 2022, 37, 477–482. [Google Scholar] [CrossRef]
- Sklenovská, N.; Ranst, M. Van Emergence of Monkeypox as the Most Important Orthopoxvirus Infection in Humans. Front. Public Health 2018, 6, 241. [Google Scholar] [CrossRef]
- Rao, A.K.; Schulte, J.; Chen, T.-H.; Hughes, C.M.; Davidson, W.; Neff, J.M.; Markarian, M.; Delea, K.C.; Wada, S.; Liddell, A.; et al. Monkeypox in a Traveler Returning from Nigeria—Dallas, Texas, July 2021. MMWR. Morb. Mortal. Wkly. Rep. 2022, 71, 509–516. [Google Scholar] [CrossRef]
- Costello, V.; Sowash, M.; Gaur, A.; Cardis, M.; Pasieka, H.; Wortmann, G.; Ramdeen, S. Imported Monkeypox from International Traveler, Maryland, USA, 2021. Emerg. Infect. Dis. 2022, 28, 1002–1005. [Google Scholar] [CrossRef]
- Haider, N.; Guitian, J.; Simons, D.; Asogun, D.; Ansumana, R.; Honeyborne, I.; Velavan, T.P.; Ntoumi, F.; Valdoleiros, S.R.; Petersen, E.; et al. Increased Outbreaks of Monkeypox Highlight Gaps in Actual Disease Burden in Sub-Saharan Africa and in Animal Reservoirs. Int. J. Infect. Dis. 2022, 122, 107–111. [Google Scholar] [CrossRef]
- Martínez, J.I.; Montalbán, E.G.; Bueno, S.J.; Martínez, F.M.; Juliá, A.N.; Díaz, S.; Marín, N.G.; Deorador, E.C.; Forte, A.N.; García, M.A.; et al. Monkeypox Outbreak Predominantly Affecting Men Who Have Sex with Men, Madrid, Spain, 26 April to 16 June 2022. Eurosurveillance 2022, 27, 2200471. [Google Scholar] [CrossRef]
- Heskin, J.; Belfield, A.; Milne, C.; Brown, N.; Walters, Y.; Scott, C.; Bracchi, M.; Moore, L.S.; Mughal, N.; Rampling, T.; et al. Transmission of Monkeypox Virus through Sexual Contact—A Novel Route of Infection. J. Infect. 2022, 85, 334–363. [Google Scholar] [CrossRef]
- Orviz, E.; Negredo, A.; Ayerdi, O.; Vázquez, A.; Muñoz-Gomez, A.; Monzón, S.; Clavo, P.; Zaballos, A.; Vera, M.; Sánchez, P.; et al. Monkeypox Outbreak in Madrid (Spain): Clinical and Virological Aspects. J. Infect. 2022, 85, 412–417. [Google Scholar] [CrossRef]
- Beer, E.M.; Id, V.B.R. A Systematic Review of the Epidemiology of Human Monkeypox Outbreaks and Implications for Outbreak Strategy. PLoS ONE 2019, 13, e0007791. [Google Scholar] [CrossRef] [PubMed]
- Jezek, Z.; Szczeniowski, M.; Paluku, K.M. Human Monkeypox: Clinical Features of 282 Patients. J. Infect. Dis. 1987, 156, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.G.; Frenkel, L.D.; Homann, S.; Guffey, J. A Case of Severe Monkeypox Virus Disease in an American Child: Emerging Infections and Changing Professional Values. Pediatr. Infect. Dis. J. 2003, 22, 1093–1096. [Google Scholar] [CrossRef]
- Shchelkunov, S.N.; Totmenin, A.V.; Babkin, I.V.; Safronov, P.F.; Ryazankina, O.I.; Petrov, N.A.; Gutorov, V.V.; Uvarova, E.A.; Mikheev, M.V.; Sisler, J.R.; et al. Human Monkeypox and Smallpox Viruses: Genomic Comparison. FEBS Lett. 2001, 509, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Huhn, G.D.; Bauer, A.M.; Yorita, K.; Graham, M.B.; Sejvar, J.; Likos, A.; Damon, I.K.; Reynolds, M.G.; Kuehnert, M.J. Clinical Characteristics of Human Monkeypox, and Risk Factors for Severe Disease. Clin. Infect. Dis. 2005, 41, 1742–1751. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Rcheulishvili, N.; Shen, Z.; Papukashvili, D.; Xie, F.; Wang, Z.; Wang, X.; He, Y.; Wang, P.G. Development of an LNP-Encapsulated MRNA-RBD Vaccine against SARS-CoV-2 and Its Variants. Pharmaceutics 2022, 14, 1101. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines-a New Era in Vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Kowalzik, F.; Schreiner, D.; Jensen, C.; Teschner, D.; Gehring, S.; Zepp, F. MRNA-Based Vaccines. Vaccines 2021, 9, 390. [Google Scholar] [CrossRef]
- Pütz, M.M.; Midgley, C.M.; Law, M.; Smith, G.L. Quantification of Antibody Responses against Multiple Antigens of the Two Infectious Forms of Vaccinia Virus Provides a Benchmark for Smallpox Vaccination. Nat. Med. 2006, 12, 1310–1315. [Google Scholar] [CrossRef]
- Wolffe, E.J.; Vijaya, S.; Moss, B. A Myristylated Membrane Protein Encoded by the Vaccinia Virus L1R Open Reading Frame Is the Target of Potent Neutralizing Monoclonal Antibodies. Virology 1995, 211, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Hirao, L.A.; Draghia-akli, R.; Prigge, J.T.; Yang, M.; Satishchandran, A.; Wu, L.; Hammarlund, E.; Khan, A.S.; Babas, T.; Rhodes, L.; et al. Multivalent Smallpox DNA Vaccine Delivered by Intradermal Electroporation Drives Protective Immunity in Nonhuman Primates Against Lethal Monkeypox Challenge. J. Infect. Dis. 2011, 203, 95–102. [Google Scholar] [CrossRef]
- Gilchuk, I.; Gilchuk, P.; Sapparapu, G.; Lampley, R.; Singh, V.; Kose, N.; Blum, D.L.; Hughes, L.J.; Satheshkumar, P.S.; Townsend, M.B.; et al. Cross-Neutralizing and Protective Human Antibody Specificities to Poxvirus Infections. Cell 2016, 167, 684–694.e9. [Google Scholar] [CrossRef] [PubMed]
- Moss, B. Smallpox Vaccines: Targets of Protective Immunity. Immunol. Rev. 2011, 239, 8–26. [Google Scholar] [CrossRef] [PubMed]
- Hooper, J.W.; Thompson, E.; Wilhelmsen, C.; Zimmerman, M.; Ichou, M.A.; Steffen, S.E.; Schmaljohn, C.S.; Schmaljohn, A.L.; Jahrling, P.B. Smallpox DNA Vaccine Protects Nonhuman Primates against Lethal Monkeypox. J. Virol. 2004, 78, 4433–4443. [Google Scholar] [CrossRef] [PubMed]
- Buchman, G.W.; Cohen, M.E.; Xiao, Y.; Richardson-Harman, N.; Silvera, P.; DeTolla, L.J.; Davis, H.L.; Eisenberg, R.J.; Cohen, G.H.; Isaacs, S.N. A Protein-Based Smallpox Vaccine Protects Non-Human Primates from a Lethal Monkeypox Virus Challenge. Vaccine 2010, 28, 6627–6636. [Google Scholar] [CrossRef] [PubMed]
- Heraud, J.; Edghill-smith, Y.; Ayala, V.; Kalisz, I.; Parrino, J.; Kalyanaraman, V.S.; Manischewitz, J.; King, L.R.; Hryniewicz, A.; Trindade, C.J.; et al. Subunit Recombinant Vaccine Protects against Monkeypox. J. Immunol. 2022, 177, 2552–2564. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2-a Multiple Sequence Alignment Editor and Analysis Workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B.; Immune, T.; et al. The Immune Epitope Database (IEDB): 2018 Update. Nucleic Acids Res. 2019, 47, 339–343. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving Sequence-Based B-Cell Epitope Prediction Using Conformational Epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef]
- Rahmani, A.; Baee, M.; Saleki, K.; Moradi, S.; Nouri, H.R. Applying High Throughput and Comprehensive Immunoinformatics Approaches to Design a Trivalent Subunit Vaccine for Induction of Immune Response against Emerging Human Coronaviruses SARS-CoV, MERS-CoV and SARS-CoV-2. J. Biomol. Struct. Dyn. 2021, 40, 6097–6113. [Google Scholar] [CrossRef] [PubMed]
- Tarrahimofrad, H.; Rahimnahal, S.; Zamani, J.; Jahangirian, E.; Aminzadeh, S. Designing a Multi-epitope Vaccine to Provoke the Robust Immune Response against Influenza A H7N9. Sci. Rep. 2021, 11, 24485. [Google Scholar] [CrossRef] [PubMed]
- Sanches, R.C.O.; Tiwari, S.; Ferreira, L.C.G.; Oliveira, F.M.; Lopes, M.D.; Passos, M.J.F.; Maia, E.H.B.; Taranto, A.G.; Kato, R.; Azevedo, V.A.C.; et al. Immunoinformatics Design of Multi-Epitope Peptide-Based Vaccine Against Schistosoma Mansoni Using Transmembrane Proteins as a Target. Front. Immunol. 2021, 12, 621706. [Google Scholar] [CrossRef] [PubMed]
- Ahammad, I.; Lira, S.S. Designing a Novel MRNA Vaccine against SARS-CoV-2: An Immunoinformatics Approach. Int. J. Biol. Macromol. 2020, 162, 820–837. [Google Scholar] [CrossRef]
- Yi, L.; Lee, Y.; Izzard, L.; Hurt, A.C. A Review of DNA Vaccines Against Influenza. Front. Immunol. 2018, 9, 1568. [Google Scholar] [CrossRef]
- Nezafat, N.; Ghasemi, Y.; Javadi, G.; Khoshnoud, M.J.; Omidinia, E. A Novel Multi-Epitope Peptide Vaccine against Cancer: An in Silico Approach. J. Theor. Biol. 2014, 349, 121–134. [Google Scholar] [CrossRef]
- Boyoglu-Barnum, S.; Ellis, D.; Gillespie, R.A.; Hutchinson, G.B.; Park, Y.; Moin, S.M.; Acton, O.J.; Ravichandran, R.; Murphy, M.; Pettie, D.; et al. Quadrivalent Influenza Nanoparticle Vaccines Induce Broad Protection. Nature 2021, 592, 623–628. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. AlgPred: Prediction of Allergenic Proteins and Mapping of IgE Epitopes. Nucleic Acids Res. 2006, 34, 202–209. [Google Scholar] [CrossRef]
- Dong, R.; Chu, Z.; Yu, F.; Zha, Y. Contriving Multi-Epitope Subunit of Vaccine for COVID-19: Immunoinformatics Approaches. Front. Immunol. 2020, 11, 1784. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. Identifying Candidate Subunit Vaccines Using an Alignment-Independent Method Based on Principal Amino Acid Properties. Vaccine 2007, 25, 856–866. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A Server for Prediction of Protective Antigens, Tumour Antigens and Subunit Vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server; The Proteomics Protocols Handbook. Hum. Press 2005, 112, 531–552. [Google Scholar] [CrossRef]
- Baek, M.; Dimaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate Prediction of Protein Structures and Interactions Using a 3-Track Neural Network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein Structure Refinement Driven by Side-Chain Repacking. Nucleic Acids Res. 2013, 41, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L.; Cox, M.M.; Hoskins, A.A. Lehninger Principles of Biochemistry, 8th ed.; Macmillan Learning: Austin, TX, USA, 2021; ISBN 9781429234146. [Google Scholar]
- Ponomarenko, J.; Bui, H.H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A New Structure-Based Tool for the Prediction of Antibody Epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Rapin, N.; Lund, O.; Castiglione, F. Immune System Simulation Online. Bioinformatics 2011, 27, 2013–2014. [Google Scholar] [CrossRef] [PubMed]
- Rapin, N.; Lund, O.; Bernaschi, M.; Castiglione, F. Computational Immunology Meets Bioinformatics: The Use of Prediction Tools for Molecular Binding in the Simulation of the Immune System. PLoS ONE 2010, 5, e9862. [Google Scholar] [CrossRef]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S.; Biology, Q. The ClusPro Web Server for Protein-Protein Docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Ghafouri, F.; Cohan, R.A.; Samimi, H.; Hosseini, A.; Naderi, M.; Noorbakhsh, F.; Haghpanah, V. Development of a Multiepitope Vaccine Against SARS-CoV-2: Immunoinformatics Study. JMIR Bioinforma. Biotechnol. 2022, 3, e36100. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; White, A. Monkeypox Virus Emerges from The Shadow of Its More Infamous Cousin: Family Biology Matters. Emerg. Microbes Infect. 2022, 11, 1768–1777. [Google Scholar] [CrossRef] [PubMed]
- Durski, K.N.; Mccollum, A.M.; Nakazawa, Y.; Petersen, B.W.; Reynolds, M.G.; Briand, S.; Djingarey, M.H.; Olson, V.; Damon, I.K.; Khalakdina, A. Emergence of Monkeypox—West and Central Africa, 1970–2017. MMWR. Morb. Mortal. Wkly. Rep. 2018, 67, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Ladnyj, D.; Ziegler, P.; Kima, E. A Human Infection Caused by Monkeypox Virus in Basankusu Territory, Democratic Republic of the Congo. Bull. World Health Organ. 1972, 46, 593–597. [Google Scholar]
- Kmiec, D.; Kirchhoff, F. Monkeypox: A New Threat? Int. J. Mol. Sci. 2022, 23, 7866. [Google Scholar] [CrossRef] [PubMed]
- Osadebe, L.; Hughes, C.M.; Lushima, R.S.; Kabamba, J.; Nguete, B.; Malekani, J.; Pukuta, E.; Karhemere, S.; Tamfum, J.M.; Okitolonda, E.W.; et al. Enhancing Case Definitions for Surveillance of Human Monkeypox in the Democratic Republic of Congo. PLoS Negl. Trop. Dis. 2017, 11, e0005857. [Google Scholar] [CrossRef] [PubMed]
- Pfäfflin, F.; Wendisch, D.; Scherer, R.; Jürgens, L.; Godzick, G.; Eva, N.; Tober, P.; Miriam, L.; Stegemann, S.; Max, V.; et al. Monkeypox In-Patients with Severe Anal Pain. Infection 2022, 51, 483–487. [Google Scholar] [CrossRef]
- Jang, Y.R.; Lee, M.; Shin, H.; Kim, J.; Choi, M.; Kim, Y.M.; Lee, M.J.; Kim, J.; Na, H.K.; Kim, J.Y.; et al. The First Case of Monkeypox in the Republic of Korea. Infect. Dis. Microbiol. Parasitol. 2022, 37, e224. [Google Scholar] [CrossRef]
- Meyer, H.; Perrichot, M.; Stemmler, M.; Emmerich, P.; Schmitz, H.; Varaine, F.; Shungu, R.; Tshioko, F.; Formenty, P. Outbreaks of Disease Suspected of Being Due to Human Monkeypox Virus Infection in the Democratic Republic of Congo in 2001. J. Clin. Microbiol. 2002, 40, 2919–2921. [Google Scholar] [CrossRef]
- Tian, D.; Zheng, T. Comparison and Analysis of Biological Agent Category Lists Based on Biosafety and Biodefense. PLoS ONE 2014, 9, e101163. [Google Scholar] [CrossRef]
- Isidro, J.; Borges, V.; Pinto, M.; Sobral, D.; Santos, J.D.; Nunes, A.; Mixão, V.; Ferreira, R.; Santos, D.; Duarte, S.; et al. Phylogenomic Characterization and Signs of Microevolution in the 2022 Multi-Country Outbreak of Monkeypox Virus. Nat. Med. 2022, 28, 1569–1572. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.R.; Isaacs, S.N. Monkeypox Virus and Insights into Its Immunomodulatory Proteins. Immunol. Rev. 2009, 225, 96–113. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Ajisegiri, W.S.; Costantino, V.; Chughtai, A.A.; Macintyre, C.R. Reemergence of Human Monkeypox and Declining Population Immunity in the Context of Urbanization, Nigeria, 2017–2020. Emerg. Infect. Dis. 2021, 27, 1007–1014. [Google Scholar] [CrossRef]
- Burki, T. Investigating Monkeypox. Lancet 2022, 399, 2254–2255. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. Monkeypox: A Potential Global Threat? J. Med. Virol. 2022, 94, 4034–4036. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.E.; Sisler, J.R.; Chandran, D.; Moss, B. Vaccinia Virus Entry/Fusion Complex Subunit A28 Is a Target of Neutralizing and Protective Antibodies. Virology 2008, 380, 394–401. [Google Scholar] [CrossRef]
- Rizk, J.G.; Lippi, G.; Henry, B.M.; Forthal, D.N.; Rizk, Y. Prevention and Treatment of Monkeypox. Drugs 2022, 82, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Kandeel, M.; Morsy, M.A.; El-lateef, H.M.A.; Marzok, M.; El-Beltagi, H.S.; Khodair, K.M.A.; Albokhadaim, I.; Venugopala, K.N. Efficacy of the Modified Vaccinia Ankara Virus Vaccine and the Replication-Competent Vaccine ACAM2000 in Monkeypox Prevention Mahmoud. Int. Immunopharmacol. 2023, 119, 110206. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA). Use of JYNNEOS (Smallpox and Monkeypox Vaccine, Live, Nonreplicating) for Preexposure Vaccination of Persons at Risk for Occupational Exposure to Orthopoxviruses: Recommendations of the Advisory Committee on Immunization Practices—United States. 2022. Available online: https://www.fda.gov/media/131078/download#:~:text=JYNNEOS is an attenuated%2C live,prevention of smallpox and monkeypox (accessed on 4 May 2023).
- Turner, J.S.; Halloran, J.A.O.; Kalaidina, E.; Kim, W.; Schmitz, A.J.; Zhou, J.Q.; Lei, T.; Thapa, M.; Chen, R.E.; Case, J.B.; et al. SARS-CoV-2 MRNA Vaccines Induce Persistent Human Germinal Centre Responses. Nature 2021, 596, 109–113. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Haas, E.J.; Angulo, F.J.; McLaughlin, J.M.; Anis, E.; Singer, S.R.; Khan, F.; Brooks, N.; Smaja, M.; Mircus, G.; Pan, K.; et al. Impact and Effectiveness of MRNA BNT162b2 Vaccine against SARS-CoV-2 Infections and COVID-19 Cases, Hospitalisations, and Deaths Following a Nationwide Vaccination Campaign in Israel: An Observational Study Using National Surveillance Data. Lancet 2021, 397, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Flynn, B.; Foulds, K.E.; Francica, J.R.; Boyoglu-Barnum, S.; Werner, A.P.; Flach, B.; O’Connell, S.; Bock, K.W.; Minai, M.; et al. Evaluation of the MRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N. Engl. J. Med. 2020, 383, 1544–1555. [Google Scholar] [CrossRef]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. MRNA Vaccines for Infectious Diseases: Principles, Delivery and Clinical Translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef] [PubMed]
- Damase, T.R.; Sukhovershin, R.; Boada, C.; Taraballi, F.; Pettigrew, R.I.; Cooke, J.P. The Limitless Future of RNA Therapeutics. Front. Bioeng. Biotechnol. 2021, 9, 628137. [Google Scholar] [CrossRef] [PubMed]
- Osterholm, M.T.; Gellin, B. Confronting 21st-Century Monkeypox. Science 2022, 377, 349. [Google Scholar] [CrossRef]
- Belongia, E.A.; Clinic, M. Smallpox Vaccine: The Good, the Bad, and the Ugly. Clin. Med. Res. 2003, 1, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Cui, L.; Gu, M.; Shi, C.; Sun, C.; Zhao, K. Outbreak of Vaccinia Virus Infection. Emerg. Infect. Dis. 2019, 25, 2017–2020. [Google Scholar]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; Mcwilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega for Making Accurate Alignments of Many Protein Sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef]
- Sievers, F.; Barton, G.J.; Higgins, D.G. Multiple Sequence Alignments. Bioinformatics, 4th ed.; Baxevanis, A.D., Bader, G.D., Wishart, D.S., Eds.; Wiley: Hoboken, NJ, USA, 2020; Volume 227, ISBN 978-1-119-33558-0. [Google Scholar]
- González-Pech, R.A.; Stephens, T.G.; Chan, C.X. Commonly Misunderstood Parameters of NCBI BLAST and Important Considerations for Users. Bioinformatics 2019, 35, 2697–2698. [Google Scholar] [CrossRef]
- Zhang, J.; Liang, Y.; Zhang, Y. Atomic-Level Protein Structure Refinement Using Fragment Guided Molecular Dynamics Conformation Sampling. Structure 2011, 19, 1784–1795. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Brenke, R.; Comeau, S.R.; Vajda, S. PIPER: An FFT-Based Protein Docking Program with Pairwise Potentials. Proteins 2006, 65, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.C.; Magalhães, M.T.Q. De; Homan, E.J. Immunoinformatic Analysis of SARS-CoV-2 Nucleocapsid Protein and Identi Fi Cation of COVID-19 Vaccine Targets. Front. Immunol. 2020, 11, 587615. [Google Scholar] [CrossRef]
- Sharma, S.; Kumari, V.; Vasant, B.; Mukherjee, A.; Pandey, R.; Kondabagil, K. Immunoinformatics Approach for a Novel Multi-Epitope Subunit Vaccine Design against Various Subtypes of Influenza A Virus. Immunobiology 2021, 226, 152053. [Google Scholar] [CrossRef] [PubMed]
- Behbahani, M.; Moradi, M.; Mohabatkar, H. In Silico Design of a Multi-epitope Peptide Construct as a Potential Vaccine Candidate for Influenza A Based on Neuraminidase Protein. In Silico Pharmacol. 2021, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Rcheulishvili, N.; Mao, J.; Papukashvili, D.; Liu, C.; Wang, Z.; Zhao, J.; Xie, F.; Pan, X.; Ji, Y.; He, Y.; et al. Designing multi-epitope mRNA construct as a universal influenza vaccine candidate for future epidemic/pandemic preparedness. Int J Biol Macromol. 2023, 226, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Ros-Lucas, A.; Correa-Fiz, F.; Bosch-Camós, L.; Rodriguez, F.; Alonso-Padilla, J. Computational Analysis of African Swine Fever Virus Protein Space for the Design of an Epitope-Based Vaccine Ensemble. Pathogens 2020, 9, 1078. [Google Scholar] [CrossRef]
- Mahmudul, H.; Shiful, I.; Sourav, C.; Abu, H.M.; Kazi, F.A.; Ziaul, F.J.; Nazmul, H.; Shakhawat, H.F.; Nazmul, H. Contriving a Chimeric Polyvalent Vaccine to Prevent Infections Caused by Herpes Simplex Virus (Type-1 and Type-2): An Exploratory Immunoinformatic Approach. J. Biomol. Struct. Dyn. 2020, 38, 2898–2915. [Google Scholar] [CrossRef]
- Ali, A.; Khan, A.; Kaushik, A.C.; Wang, Y.; Ali, S.S.; Junaid, M.; Saleem, S.; Cho, W.C.S.; Mao, X.; Wei, D. Immunoinformatic and Systems Biology Approaches to Predict and Validate Peptide Vaccines against Epstein–Barr Virus (EBV). Sci. Rep. 2019, 9, 720. [Google Scholar] [CrossRef]
- Papukashvili, D.; Rcheulishvili, N.; Liu, C.; Wang, X.; He, Y.; Wang, P.G. Strategy of developing nucleic acid-based universal monkeypox vaccine candidates. Front Immunol. 2022, 13, 1050309. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (MPXV) | Name (VARV) | Name (VACV) | Location | Function and Characteristics |
|---|---|---|---|---|
| A29 | A30 | A27 | MV | Surface membrane fusion protein; Binds to cell surface heparan; Neutralizing antibody target |
| A30 | A31 | A28 | MV | Envelope protein; Virus entry into a host; Cell–cell fusion (syncytial formation); Neutralizing antibody target |
| A35 | A36 | A33 | EV | Envelope glycoprotein; Formation of actin-containing microvilli and cell-to-cell spread of virion; Neutralizing antibody target; Target of complement-mediated cytolysis |
| B6 | B7 | B5 | EV | Palmitylated glycoprotein; Required for efficient cell spread; Complement control |
| M1 | M1 | L1 | MV | Myristylated surface membrane protein; Virus entry into a host; Neutralizing antibody target |
| # | Epitopes | Residue Number | Score |
|---|---|---|---|
| 1 | A:K287, A:E289, A:Q290, A:T291, A:S292, A:K293, A:K294, A:V295, A:S296, A:E297, A:L298, A:Y299, A:N300, A:K301, A:P302, A:L303, A:Y304, A:K305, A:K306, A:E307, A:E308, A:K309, A:N310, A:G311, A:N312, A:T313, A:S314, A:W315, A:N316, A:D317, A:T318, A:V319, A:K320, A:K321, A:P322, A:D323, A:D324, A:E325, A:T326, A:D327, A:L328, A:S329, A:K330, A:L331 | 44 | 0.859 |
| 2 | A:I511, A:V512, A:R513, A:L514, A:N515, A:Q516, A:C517, A:M518, A:S519, A:A520, A:N521, A:G522, A:G523, A:G524, A:S525, A:A526, A:S527, A:Y528, A:I529, A:S530, A:C531, A:T532, A:A533, A:N534, A:S535, A:N537, A:I539, A:G546, A:V547, A:I548, A:H549, A:L550, A:S551, A:C552, A:K553, A:S554, A:G555, A:F556, A:I557, A:L558, A:T559, A:G560, A:G561, A:G562, A:S563, A:M564, A:K565, A:T566, A:I567, A:S568, A:V569, A:V570, A:S589, A:V592, A:L593, A:V594, A:C595, A:S596, A:C597, A:N598, A:G599, A:G600, A:G601, A:S602, A:K603, A:I604, A:Q605, A:N606, A:V607, A:I608, A:I609, A:D610, A:E611, A:C612, A:Y613, A:G614, A:G615, A:G616, A:S617, A:A618, A:A619, A:L620, A:F621, A:M622, A:Y623, A:Y624, A:A625, A:K626, A:R627, A:G628, A:G629, A:G630, A:S631, A:N632, A:T633, A:L634, A:S635, A:E636, A:R637, A:I638, A:S639, A:S640, A:K641, A:H642, A:H643, A:H644, A:H645, A:H646, A:H647 | 109 | 0.761 |
| 3 | A:K212, A:D216, A:N217, A:K218, A:K221, A:G224, A:G225, A:T226, A:P227, A:A228, A:K229, A:K230, A:Q231, A:D232, A:V233, A:N234, A:D235, A:T236, A:I237, A:S238, A:D239, A:K240, A:K241, A:G242, A:P243, A:N244, A:N245, A:T246, A:R247, A:K248, A:K249, A:S250, A:T251, A:H252, A:R253, A:K254, A:V255, A:F401, A:I402, A:V403, A:V404, A:A405, A:T406, A:A407, A:A408, A:V409, A:C410, A:L411, A:L412, A:F413, A:I414, A:G415, A:G416, A:G417, A:S418, A:M419, A:N420, A:S421, A:L422, A:S423, A:I424, A:F425, A:F426, A:I427, A:V428, A:V429, A:A430, A:T431, A:A432, A:A433 | 70 | 0.733 |
| 4 | A:D9, A:V10, A:A11, A:A12, A:P13, A:H14, A:R15, A:Q16, A:P17, A:L18, A:T19, A:S20, A:S21, A:E22, A:R23, A:I24, A:D25, A:M66, A:A67, A:E68, A:K69, A:D70, A:G71, A:C72, A:F73, A:Q74, A:S75, A:G76, A:F77, A:N78, A:E79, A:E80, A:T81, A:C82, A:L83, A:V84, A:K85, A:I86, A:I87, A:T88, A:L91, A:V120, A:L121, A:I122, A:Q123, A:F124, A:L125, A:Q126, A:K128, A:A129, A:K130, A:N131, A:L132, A:D133, A:A134, A:I135, A:T136, A:T137, A:P138, A:D139, A:P140, A:T141, A:T142, A:A144, A:S145, A:T148, A:K149, A:S176, A:L177, A:R178, A:A179, A:L180, A:R181, A:Q182, A:M183, A:G184, A:G185, A:G186, A:S187, A:A188, A:K189, A:F190, A:V191, A:A192, A:A193, A:W194, A:T195, A:K197, A:A198, A:E201 | 90 | 0.69 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rcheulishvili, N.; Mao, J.; Papukashvili, D.; Feng, S.; Liu, C.; Yang, X.; Lin, J.; He, Y.; Wang, P.G. Development of a Multi-Epitope Universal mRNA Vaccine Candidate for Monkeypox, Smallpox, and Vaccinia Viruses: Design and In Silico Analyses. Viruses 2023, 15, 1120. https://doi.org/10.3390/v15051120
Rcheulishvili N, Mao J, Papukashvili D, Feng S, Liu C, Yang X, Lin J, He Y, Wang PG. Development of a Multi-Epitope Universal mRNA Vaccine Candidate for Monkeypox, Smallpox, and Vaccinia Viruses: Design and In Silico Analyses. Viruses. 2023; 15(5):1120. https://doi.org/10.3390/v15051120
Chicago/Turabian StyleRcheulishvili, Nino, Jiawei Mao, Dimitri Papukashvili, Shunping Feng, Cong Liu, Xidan Yang, Jihui Lin, Yunjiao He, and Peng George Wang. 2023. "Development of a Multi-Epitope Universal mRNA Vaccine Candidate for Monkeypox, Smallpox, and Vaccinia Viruses: Design and In Silico Analyses" Viruses 15, no. 5: 1120. https://doi.org/10.3390/v15051120
APA StyleRcheulishvili, N., Mao, J., Papukashvili, D., Feng, S., Liu, C., Yang, X., Lin, J., He, Y., & Wang, P. G. (2023). Development of a Multi-Epitope Universal mRNA Vaccine Candidate for Monkeypox, Smallpox, and Vaccinia Viruses: Design and In Silico Analyses. Viruses, 15(5), 1120. https://doi.org/10.3390/v15051120