
Area Wide Monitoring of Plant and Honey Bee (Apis mellifera) Viruses in Blueberry (Vaccinium corymbosum) Agroecosystems Facilitated by Honey Bee Pollination

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Sample Collection and RNA Extraction
2.2. RNA Sequencing
2.3. Bioinformatics
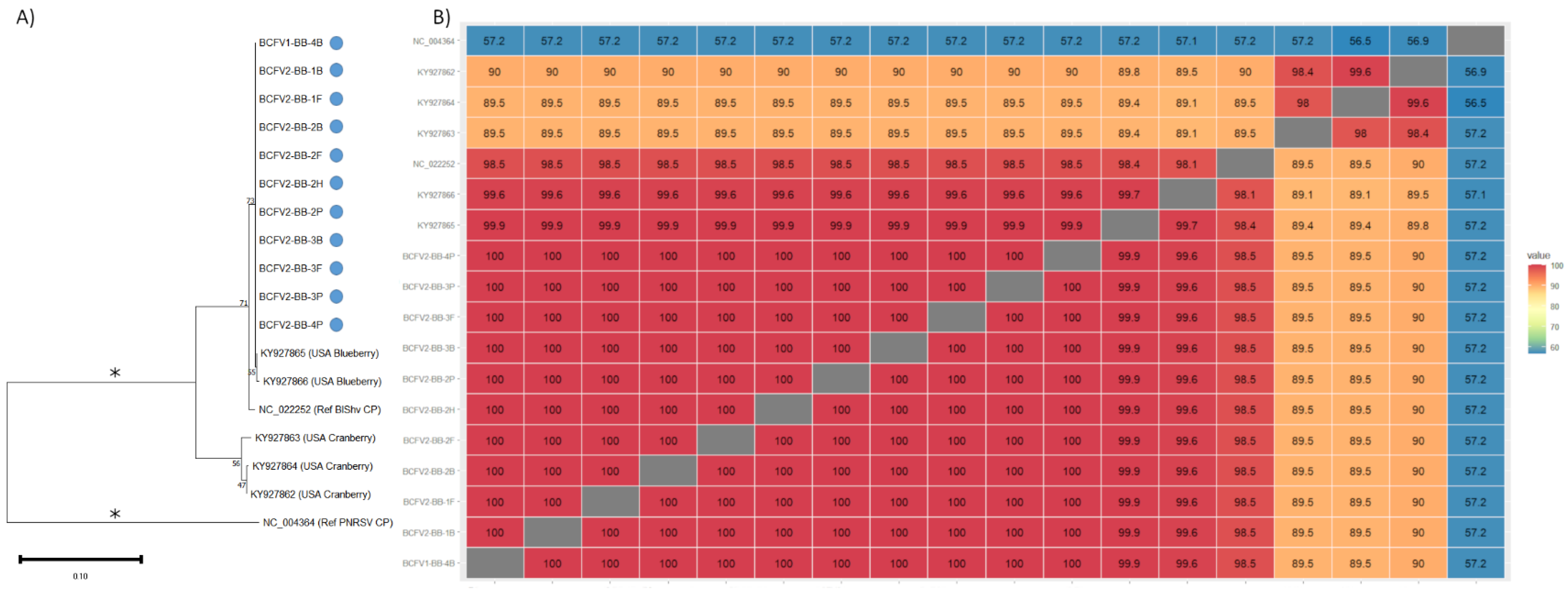
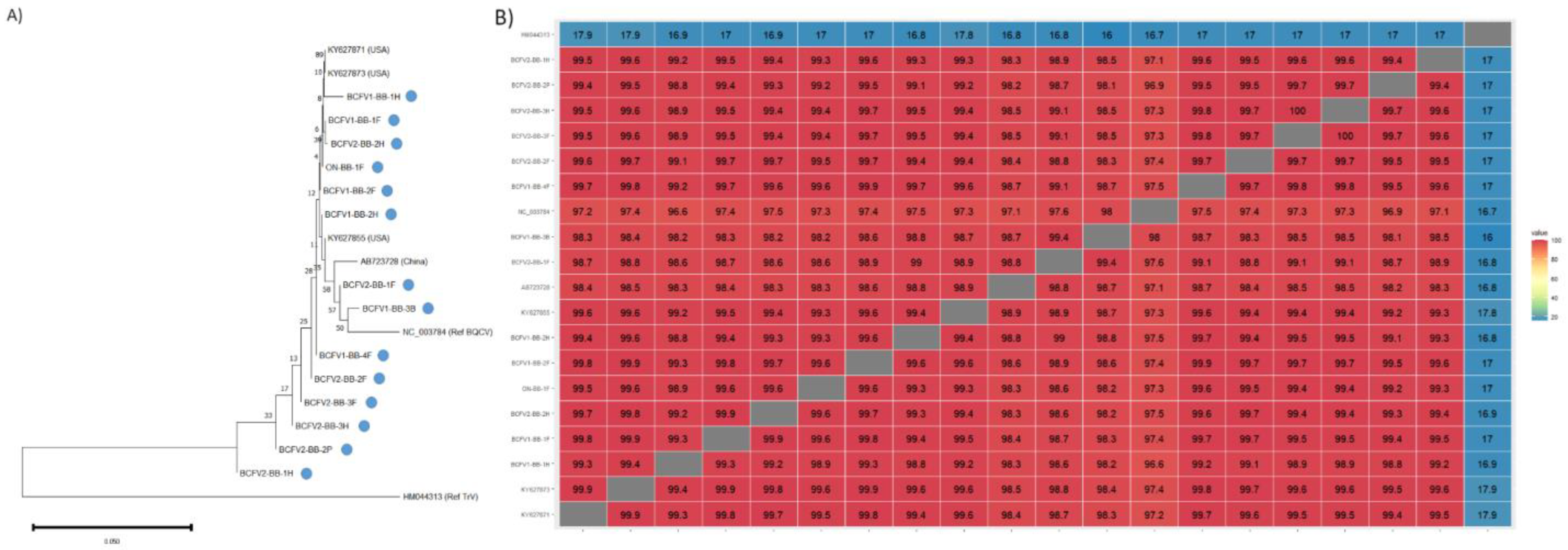
2.4. Phylogenetics and Sequence Analysis
3. Results
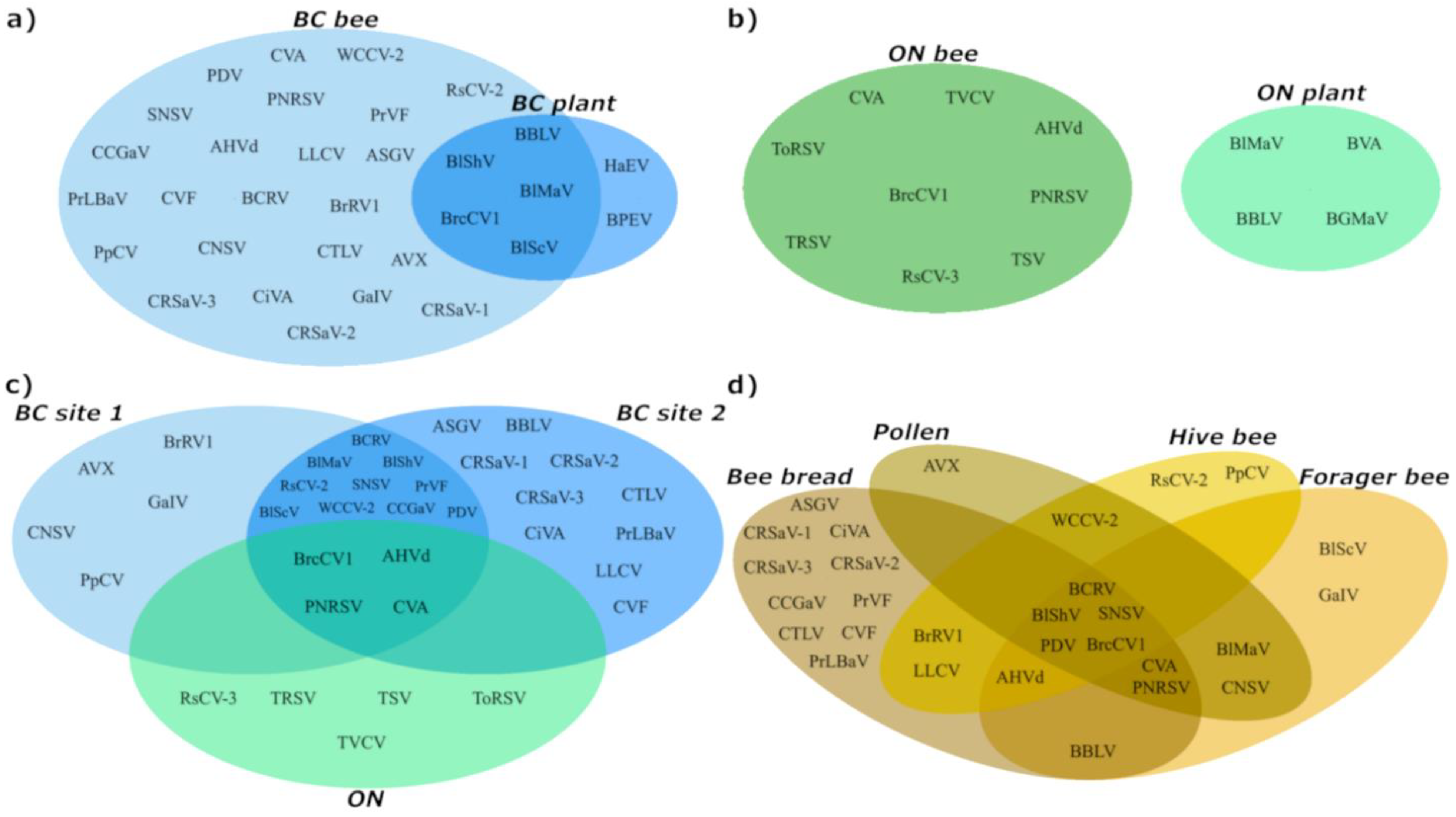
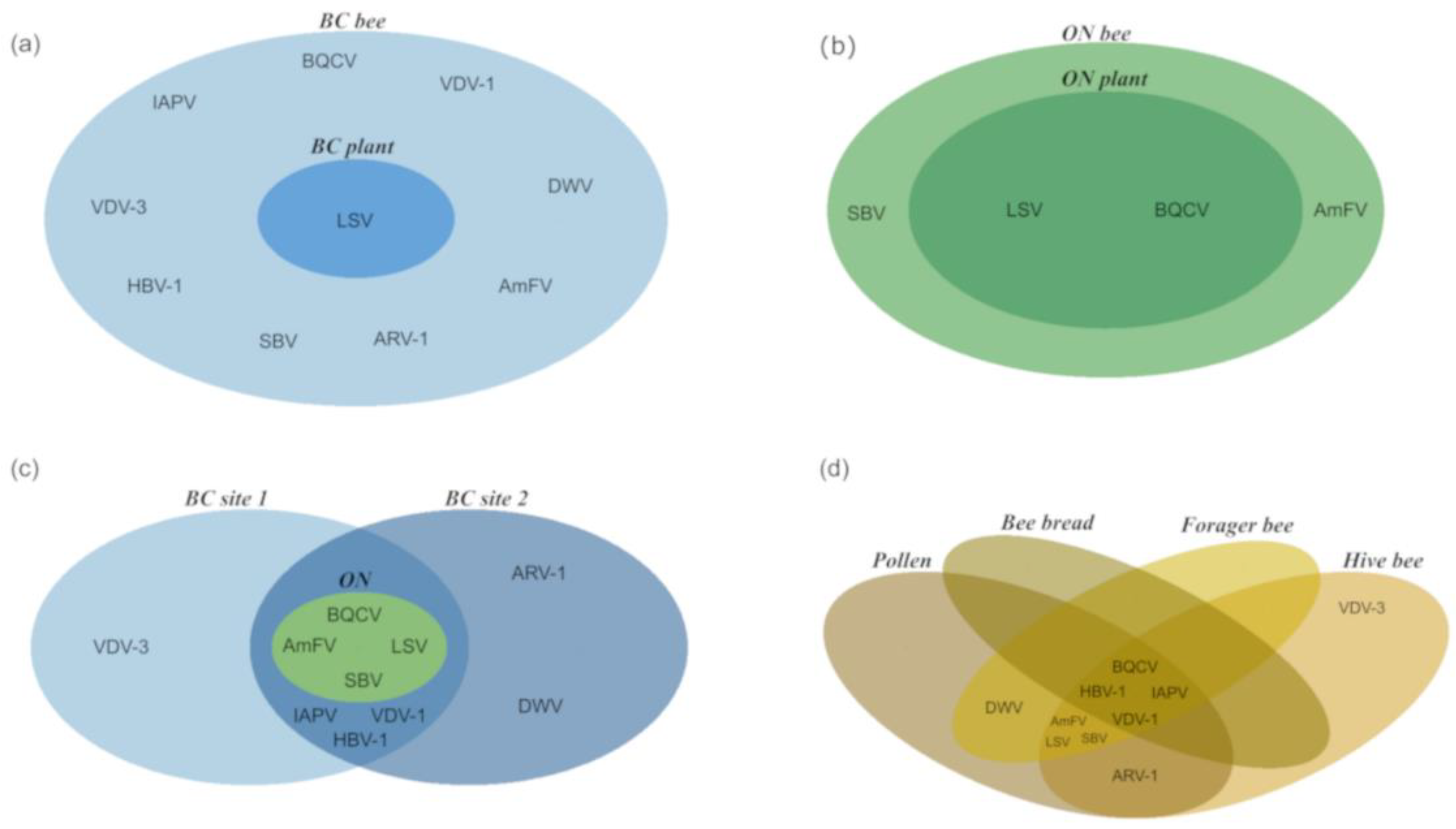
3.1. Virus Detection from Managed Bee Hives in Canadian Blueberry Farms
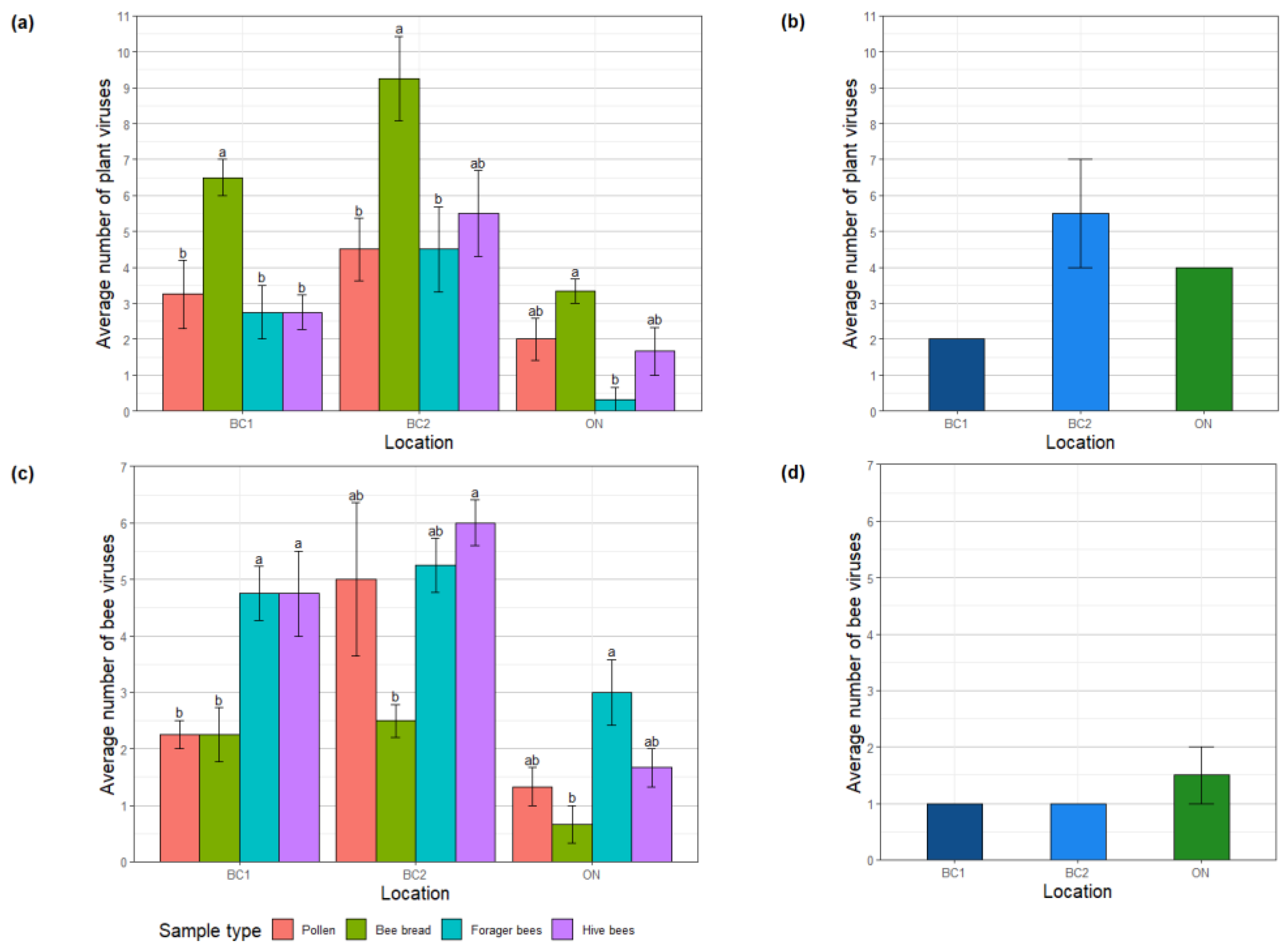
3.2. Comparison of Viral Profiles from Bee Pollen, Forager Bees, Hive Bees and Bee Bread
3.3. Viral Diversity in Bee-Collected Samples
4. Discussion
4.1. Plant Virome
4.2. Bee Virome
4.3. Metagenomics-based Monitoring of Plant Viruses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, R.R.; Constable, F.; Tzanetakis, I.E. Quarantine regulations and the impact of modern detection methods. Ann. Rev. Phytopathol. 2016, 54, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Villamor, D.E.V.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. Comparison of High Throughput Sequencing to Standard Protocols for Virus Detection in Berry Crops. Plant Dis. 2022, 106, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J.; Martin, D.P.; Roumagnac, P. Plant Virus Metagenomics: Advances in Virus Discovery. Phytopathology 2015, 105, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Piombo, E.; Abdelfattah, A.; Droby, S.; Wisniewski, M.; Spadaro, D.; Schena, L. Metagenomics Approaches for the Detection and Surveillance of Emerging and Recurrent Plant Pathogens. Microorganisms 2021, 9, 188. [Google Scholar] [CrossRef]
- Saad, N.; Olmstead, J.W.; Varsani, A.; Polston, J.E.; Jones, J.B.; Folimonova, S.Y.; Harmon, P.F. Discovery of Known and Novel Viruses in Wild and Cultivated Blueberry in Florida through Viral Metagenomic Approaches. Viruses 2021, 13, 1165. [Google Scholar] [CrossRef]
- Cunningham, M.M.; Tran, L.; McKee, C.G.; Polo, R.O.; Newman, T.; Lansing, L.; Griffiths, J.S.; Bilodeau, G.J.; Rott, M.; Guarna, M.M. Honey bees as biomonitors of environmental contaminants, pathogens, and climate change. Ecol. Indic. 2022, 134, 108457. [Google Scholar] [CrossRef]
- Roberts, J.M.K.; Ireland, K.B.; Tay, W.T.; Paini, D. Honey bee-assisted surveillance for early plant virus detection. Ann. Appl. Biol. 2018, 173, 285–293. [Google Scholar] [CrossRef]
- Granberg, F.; Vicente-Rubiano, M.; Rubio-Guerri, C.; Karlsson, O.E.; Kukielka, D.; Belák, S.; Sánchez-Vizcaíno, J.M. Metagenomic detection of viral pathogens in Spanish honeybees: Co-infection by aphid lethal paralysis, Israel acute paralysis and Lake Sinai viruses. PLoS ONE 2013, 8, e57459. [Google Scholar] [CrossRef]
- Roberts, J.; Anderson, D.L.; Durr, P. Metagenomic analysis of Varroa-free Australian honey bees (Apis mellifera) shows a diverse Picornavirales virome. J. Gen. Virol. 2018, 99, 818–826. [Google Scholar] [CrossRef]
- Kavalappara, S.R.; Milner, H.; Konakalla, N.C.; Morgan, K.; Sparks, A.N.; McGregor, C.; Culbreath, A.K.; Wintermantel, W.M.; Bag, S. High Throughput Sequencing-Aided Survey Reveals Widespread Mixed Infections of Whitefly-Transmitted Viruses in Cucurbits in Georgia, USA. Viruses 2021, 13, 988. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Mayer, D.F. Crop Pollination by Bees; CABI Publishing: New York, NY, USA, 2000. [Google Scholar] [CrossRef]
- Isaacs, R.; Kirk, A.K. Pollination services provided to small and large highbush blueberry fields by wild and managed bees. J. Appl. Ecol. 2010, 47, 841–849. [Google Scholar] [CrossRef]
- Aizen, M.A.; Harder, L.D. The Global Stock of Domesticated Honey Bees Is Growing Slower Than Agricultural Demand for Pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef]
- Kleijn, D.; Winfree, R.; Bartomeus, I.; Carvalheiro, L.G.; Henry, M.; Isaacs, R.; Klein, A.-M.; Kremen, C.; M’Gonigle, L.K.; Rader, R.; et al. Delivery of crop pollination services is an insufficient argument for wild pollinator conservation. Nat. Commun. 2015, 6, 7414. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.K.; Seeley, T.D. Foraging Strategy of Honeybee Colonies in a Temperate Deciduous Forest. Ecology 1982, 63, 1790–1801. [Google Scholar] [CrossRef]
- Beekman, M.; Ratnieks, F.L.W. Long-range foraging by the honey-bee, Apis mellifera L. Funct. Ecol. 2000, 14, 490–496. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Kuhn, A. Honeybee foraging in differentially structured landscapes. Proc. R. Soc. B Boil. Sci. 2003, 270, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Couvillon, M.J.; Schürch, R.; Ratnieks, F.L.W. Waggle Dance Distances as Integrative Indicators of Seasonal Foraging Challenges. PLoS ONE 2014, 9, e93495. [Google Scholar] [CrossRef]
- Balfour, N.J.; Ratnieks, F.L.W. Using the waggle dance to determine the spatial ecology of honey bees during commercial crop pollination. Agric. For. Entomol. 2017, 19, 210–216. [Google Scholar] [CrossRef]
- Tremblay, D.; Duceppe, M.; Thurston, G.B.; Gagnon, M.; Côté, M.; Bilodeau, G.J. High-resolution biomonitoring of plant pathogens and plant species using metabarcoding of pollen pellet contents collected from a honey bee hive. Environ. DNA 2019, 1, 155–175. [Google Scholar] [CrossRef]
- AAFC—Agriculture and Agri-Food Canada. Statistical Overview of the Canadian Fruit Industry 2021. Horticulture Section, Crops and Horticulture Division (2022). Available online: https://agriculture.canada.ca/sites/default/files/documents/2022-12/Fruit%20Report_2021_ENG.pdf (accessed on 22 March 2023).
- Moisan-DeSerres, J.; Chagnon, M.; Fournier, V. Influence of windbreaks and forest borders on abundance and species richness of native pollinators in lowbush blueberry fields in Québec, Canada. Can. Entomol. 2015, 147, 432–442. [Google Scholar] [CrossRef]
- Gibbs, J.; Elle, E.; Bobiwash, K.; Haapalainen, T.; Isaacs, R. Contrasting Pollinators and Pollination in Native and Non-Native Regions of Highbush Blueberry Production. PLoS ONE 2016, 11, e0158937. [Google Scholar] [CrossRef] [PubMed]
- Gruszka, J. Beekeeping in Western Canada; Alberta Agriculture, Food and Rural Development, Publishing Branch: Edmonton, AB, USA, 1998. [Google Scholar] [CrossRef]
- Pernal, S.F. The Social Life of Honey Bees. Veter Clin. North Am. Food Anim. Pr. 2021, 37, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Winston, M.L. The Biology of the Honey Bee; Harvard Univ. Press: Cambridge, MA, USA, 1987. [Google Scholar]
- Vasquez, A.; Olofsson, R.C. The lactic acid bacteria involved in the production of bee pollen and bee bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- Anderson, K.E.; Carroll, M.J.; Sheehan, T.; Mott, B.M.; Maes, P.; Corby-Harris, V. Hive-stored pollen of honey bees: Many lines of evidence are consistent with pollen preservation, not nutrient conversion. Mol. Ecol. 2014, 23, 5904–5917. [Google Scholar] [CrossRef] [PubMed]
- De-Melo, A.A.M.; de Almeida-Muradian, L.B. Chemical Composition of Bee Pollen. Bee Prod.-Chem. Biol. Prop. 2017, 221–259. [Google Scholar] [CrossRef]
- Card, S.D.; Pearson, M.N.; Clover, G.R.G. Plant pathogens transmitted by pollen. Australas. Plant Pathol. 2007, 36, 455–461. [Google Scholar] [CrossRef]
- Jones, R.A. Plant and Insect Viruses in Managed and Natural Environments: Novel and Neglected Transmission Pathways. Adv. Virus Res. 2018, 101, 149–187. [Google Scholar] [CrossRef]
- Spivak, M.; Cariveau, D.P. Flowers as parasite transmission hubs. Nat. Ecol. Evol. 2020, 4, 1298–1299. [Google Scholar] [CrossRef]
- Proesmans, W.; Albrecht, M.; Gajda, A.; Neumann, P.; Paxton, R.J.; Pioz, M.; Polzin, C.; Schweiger, O.; Settele, J.; Szentgyörgyi, H.; et al. Pathways for novel epidemiology: Plant-pollinator-pathogen networks and global change. Trends Ecol. Evol. 2021, 36, 623–636. [Google Scholar] [CrossRef]
- Desai, S.D.; Currie, R.W. Effects of Wintering Environment and Parasite–Pathogen Interactions on Honey Bee Colony Loss in North Temperate Regions. PLoS ONE 2016, 11, e0159615. [Google Scholar] [CrossRef]
- Gisder, S.; Genersch, E. Viruses of commercialized insect pollinators. J. Invertebr. Pathol. 2017, 147, 51–59. [Google Scholar] [CrossRef]
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, Pathogenicity, and Impacts. Annu. Rev. Entomol. 2019, 64, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Beaurepaire, A.; Piot, N.; Doublet, V.; Antunez, K.; Campbell, E.; Chantawannakul, P.; Chejanovsky, N.; Gajda, A.; Heerman, M.; Panziera, D.; et al. Diversity and Global Distribution of Viruses of the Western Honey Bee, Apis mellifera. Insects 2020, 11, 239. [Google Scholar] [CrossRef]
- Harwood, G.P.; Dolezal, A.G. Pesticide–Virus Interactions in Honey Bees: Challenges and Opportunities for Understanding Drivers of Bee Declines. Viruses 2020, 12, 566. [Google Scholar] [CrossRef] [PubMed]
- Borba, R.S.; Hoover, S.E.; Currie, R.W.; Giovenazzo, P.; Guarna, M.M.; Foster, L.J.; Zayed, A.; Pernal, S.F. Phenomic analysis of the honey bee pathogen-web and its dynamics on colony productivity, health and social immunity behaviors. PLoS ONE 2022, 17, e0263273. [Google Scholar] [CrossRef]
- Alger, S.A.; Burnham, P.A.; Brody, A.K. Flowers as viral hot spots: Honey bees (Apis mellifera) unevenly deposit viruses across plant species. PLoS ONE 2019, 14, e0221800. [Google Scholar] [CrossRef] [PubMed]
- Fetters, A.M.; Cantalupo, P.G.; Na Wei, N.; Robles, M.T.S.; Stanley, A.; Stephens, J.D.; Pipas, J.M.; Ashman, T.-L. The pollen virome of wild plants and its association with variation in floral traits and land use. Nat. Commun. 2022, 13, 523. [Google Scholar] [CrossRef]
- Bristow, P.R.; Martin, R.R. Transmission and the Role of Honeybees in Field Spread of Blueberry Shock Ilarvirus, a Pollen-Borne Virus of Highbush Blueberry. Phytopathology 1999, 89, 124–130. [Google Scholar] [CrossRef]
- Thomas-Sharma, S.; Wells-Hansen, L.; Page, R.; Kartanos, V.; Saalau-Rojas, E.; Lockhart, B.E.; McManus, P.S. Characterization of Blueberry shock virus, an emerging ilarvirus in Cranberry. Plant Dis. 2018, 102, 91–97. [Google Scholar] [CrossRef]
- Martin, R.R.; Tzanetakis, I.E. High risk blueberry viruses by region in North America; implication for certifications, nurseries, and fruit production. Viruses 2018, 10, 342. [Google Scholar] [CrossRef]
- Saad, N.; Olmstead, J.W.; Jones, J.B.; Varsani, A.; Harmon, P.F. Known and New Emerging Viruses Infecting Blueberry. Plants 2021, 10, 2172. [Google Scholar] [CrossRef]
- Massart, S.; Adams, I.; Al Rwahnih, M.; Baeyen, S.; Bilodeau, G.J.; Blouin, A.G.; Boonham, N.; Candresse, T.; Chandellier, A.; De Jonghe, K.; et al. Guidelines for the reliable use of high throughput sequencing technologies to detect plant pathogens and pests. Peer Community J. 2022, 2, e62. [Google Scholar] [CrossRef]
- Lebas, B.; Adams, I.; Al Rwahnih, M.; Baeyen, S.; Bilodeau, G.J.; Blouin, A.G.; Boonham, N.; Candresse, T.; Chandelier, A.; De Jonghe, K.; et al. Facilitating the adoption of high-throughput sequencing technologies as a plant pest diagnostic test in laboratories: A step-by-step description. EPPO Bull. 2022, 52, 394–418. [Google Scholar] [CrossRef]
- Kesanakurti, P.; Belton, M.; Saeed, H.; Rast, H.; Boyes, I.; Rott, M. Screening for plant viruses by next generation sequencing using a modified double strand RNA extraction protocol with an internal amplification control. J. Virol. Methods 2016, 236, 35–40. [Google Scholar] [CrossRef]
- Hong, C.; Manimaran, S.; Shen, Y.; Perez-Rogers, J.F.; Byrd, A.L.; Castro-Nallar, E.; A Crandall, K.; Johnson, W.E. PathoScope 2.0: A complete computational framework for strain identification in environmental or clinical sequencing samples. Microbiome 2014, 2, 33. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Rott, M.; Xiang, Y.; Boyes, I.; Belton, M.; Saeed, H.; Kesanakurti, P.; Hayes, S.; Lawrence, T.; Birch, C.; Bhagwat, B.; et al. Application of Next Generation Sequencing for Diagnostic Testing of Tree Fruit Viruses and Viroids. Plant Dis. 2017, 101, 1489–1499. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Kinoti, W.M.; Constable, F.E.; Nancarrow, N.; Plummer, K.; Rodoni, B. Generic Amplicon Deep Sequencing to Determine Ilarvirus Species Diversity in Australian Prunus. Front. Microbiol. 2017, 8, 1219. [Google Scholar] [CrossRef] [PubMed]
- Simkovich, A.J.; Li, Y.; Kohalmi, S.E.; Griffiths, J.S.; Wang, A. Molecular Identification of Prune Dwarf Virus (PDV) Infecting Sweet Cherry in Canada and Development of a PDV Full-Length Infectious cDNA Clone. Viruses 2021, 13, 2025. [Google Scholar] [CrossRef] [PubMed]
- Childress, A.M.; Ramsdell, D.C. Bee-mediated transmission of blueberry leaf mottle virus via infected pollen in highbush blueberry. Phytopathology 1987, 77, 167–172. [Google Scholar] [CrossRef]
- Bristow, P.R.; Martin, R.R.; Windom, G.E. Transmission, field spread, cultivar response and impact on high bush blueberry infected with Blueberry scorch virus. Phytopathology 2000, 90, 474–479. [Google Scholar] [CrossRef]
- Isogai, M.; Miyoshi, K.; Watanabe, M.; Yoshikawa, N. Characterization of horizontal transmission of blueberry latent spherical virus by pollen. Arch. Virol. 2020, 165, 2807–2815. [Google Scholar] [CrossRef]
- Drummond, F.A.; Lund, J.; Eitzer, B. Honey Bee Health in Maine Wild Blueberry Production. Insects 2022, 12, 523. [Google Scholar] [CrossRef]
- Mitra, A.; Jarugula, S.; Hoheisel, G.-A.; Naidu, R.A. First Report of Tobacco Ringspot Virus in Highbush Blueberry in Washington State. Plant Dis. 2021, 105, 2739. [Google Scholar] [CrossRef] [PubMed]
- Rivera, L.; Zamorano, A.; Fiore, N. Genetic divergence of tomato ringspot virus. Arch. Virol. 2016, 161, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- Isogai, M.; Muramatu, S.; Watanabe, M.; Yoshikawa, N. Complete nucleotide sequence and latency of a novel blueberry-infecting closterovirus. J. Gen. Plant Pathol. 2013, 79, 123–127. [Google Scholar] [CrossRef]
- Thekke-Veetil, T.; Ho, T. Molecular characterization of a new vitivirus discovered in a blueberry plant with green mosaic symptoms. Arch. Virol. 2019, 164, 2609–2611. [Google Scholar] [CrossRef]
- Tzanetakis, I.; Martin, R.R.; Scott, S.W. Genomic sequences of blackberry chlorotic ringspot virus and strawberry necrotic shock virus and the phylogeny of viruses in subgroup 1 of the genus Ilarvirus. Arch. Virol. 2010, 155, 557–561. [Google Scholar] [CrossRef]
- Tzanetakis, I.E.; Postman, J.D.; Martin, R.R. First report of Blackberry chlorotic ringspot virus in Rubus sp. in the United States. Plant Dis. 2007, 91, 463. [Google Scholar] [CrossRef]
- Chiapello, M.; Rodríguez-Romero, J.; Nerva, L.; Forgia, M.; Chitarra, W.; Ayllón, M.A.; Turina, M. Putative new plant viruses associated with Plasmopara viticola-infected grapevine samples. Ann. Appl. Biol. 2020, 176, 180–191. [Google Scholar] [CrossRef]
- Herranz, M.C.; Al Rwahnih, M.; Sánchez-Navarro, J.A.; Elena, S.F.; Choueiri, E.; Myrta, A.; Pallás, V. Low genetic variability in the coat and movement proteins of American plum line pattern virus isolates from different geographic origins. Arch. Virol. 2008, 153, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Daughenbaugh, K.F.; Kahnonitch, I.; Carey, C.C.; McMenamin, A.J.; Wiegand, T.; Erez, T.; Arkin, N.; Ross, B.; Wiedenheft, B.; Sadeh, A.; et al. Metatranscriptome Analysis of Sympatric Bee Species Identifies Bee Virus Variants and a New Virus, Andrena-Associated Bee Virus-1. Viruses 2021, 13, 291. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, C.; Hu, T.; Li, J.; Zhou, H.; Ji, J.; Wu, J.; Kang, W.; Holmes, E.C.; Shi, W.; et al. Nationwide genomic surveillance reveals the prevalence and evolution of honeybee viruses in China. Microbiome 2023, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Pettis, J.S.; Corona, M.; Chen, W.P.; Li, C.J.; Spivak, M.; Visscher, P.K.; DeGrandi-Hoffman, G.; Boncristiani, H.; Zhao, Y.; et al. Israeli Acute Paralysis Virus: Epidemiology, Pathogenesis and Implications for Honey Bee Health. PLOS Pathog. 2014, 10, e1004261. [Google Scholar] [CrossRef]
- Amiri, E.; Seddon, G.; Zuluaga Smith, W.; Strand, M.K.; Tarpy, D.R.; Rueppell, O. Israeli Acute Paralysis Virus: Honey Bee Queen–Worker Interaction and Potential Virus Transmission Pathways. Insects 2019, 10, 123. [Google Scholar] [CrossRef]
- Singh, R.; Levitt, A.L.; Rajotte, E.G.; Holmes, E.C.; Ostiguy, N.; Vanengelsdorp, D.; Lipkin, W.I.; Depamphilis, C.W.; Toth, A.L.; Cox-Foster, D.L. RNA Viruses in Hymenopteran Pollinators: Evidence of Inter-Taxa Virus Transmission via Pollen and Potential Impact on Non-Apis Hymenopteran Species. PLoS ONE 2010, 5, e14357. [Google Scholar] [CrossRef]
- Ravoet, J.; De Smet, L.; Wenseleers, T.; de Graaf, D.C. Genome sequence heterogeneity of Lake Sinai Virus found in honey bees and Orf1/RdRP-based polymorphisms in a single host. Virus Res. 2015, 201, 67–72. [Google Scholar] [CrossRef]
- Yañez, O.; Piot, N.; Dalmon, A.; de Miranda, J.R.; Chantawannakul, P.; Panziera, D.; Amiri, E.; Smagghe, G.; Schroeder, D.; Chejanovsky, N. Bee Viruses: Routes of Infection in Hymenoptera. Front. Microbiol. 2020, 11, 943. [Google Scholar] [CrossRef]
- Daughenbaugh, K.F.; Martin, M.; Brutscher, L.M.; Cavigli, I.; Garcia, E.; Lavin, M.; Flenniken, M.L. Honey Bee Infecting Lake Sinai Viruses. Viruses 2015, 7, 3285–3309. [Google Scholar] [CrossRef]
- Rosseel, T.; Pardon, B.; De Clercq, K.; Ozhelvaci, O.; Van Borm, S. False-Positive Results in Metagenomic Virus Discovery: A Strong Case for Follow-Up Diagnosis. Transbound. Emerg. Dis. 2014, 61, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Beaver-Kanuya, E.; Harper, S. Detection and quantification of four viruses in Prunus pollen: Implications for biosecurity. J. Virol. Methods 2019, 271, 113673. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Virus | Genus | BC Site 1 | BC Site 2 | Average Frequency of Detection (%) * | Average Genome Coverage (%) * | Average VRPM * | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bee Bread | Forager Bee | Hive Bee | Pollen | Bee Bread | Forager Bee | Hive Bee | Pollen | |||||
| n = 4 | n = 4 | n = 4 | n = 4 | n = 4 | n = 4 | n = 4 | n = 4 | |||||
| Frequency (%) | ||||||||||||
| Blueberry shock virus | Ilarvirus | 75 | 50 | 50 | 50 | 100 | 100 | 100 | 100 | 78 | 57.5 | 425 |
| Prune dwarf virus | Ilarvirus | 100 | 50 | 75 | 50 | 100 | 50 | 100 | 50 | 72 | 52.3 | 356 |
| Strawberry necrotic shock virus | Ilarvirus | 75 | 25 | 0 | 75 | 50 | 50 | 50 | 75 | 50 | 56.6 | 281 |
| Cherry virus A | Capillovirus | 75 | 25 | 25 | 0 | 100 | 50 | 75 | 0 | 44 | 63.2 | 176 |
| Brassica campestris chrysovirus 1 | Alphachrysovirus | 50 | 0 | 0 | 0 | 50 | 25 | 75 | 75 | 34 | 52.8 | 24 |
| White clover cryptic virus 2 | Betapartitivirus | 0 | 0 | 50 | 50 | 0 | 0 | 75 | 25 | 25 | 24.5 | 43 |
| Prunus necrotic ringspot virus | Ilarvirus | 50 | 25 | 25 | 0 | 100 | 0 | 0 | 0 | 25 | 62.9 | 290 |
| Blackberry chlorotic ringspot virus | Ilarvirus | 25 | 0 | 0 | 0 | 25 | 25 | 50 | 50 | 22 | 17.4 | 0 |
| Apple hammerhead viroid | Pelamoviroid | 75 | 0 | 0 | 25 | 25 | 25 | 0 | 25 | 22 | 58.6 | 5 |
| Blueberry mosaic-associated virus | Ophiovirus | 0 | 25 | 0 | 0 | 0 | 75 | 25 | 0 | 16 | 34.3 | 40 |
| Citrus concave gum-associated virus | Coguvirus | 25 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 9 | 29.6 | 143 |
| Prunus virus F | Fabavirus | 50 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 9 | 77.7 | 16 |
| Lilac leaf chlorosis virus | Ilarvirus | 0 | 0 | 0 | 0 | 50 | 0 | 0 | 25 | 9 | 35.0 | 230 |
| Brassica rapa virus 1 | unclassified Rhabdoviridae | 50 | 0 | 0 | 25 | 0 | 0 | 0 | 0 | 9 | 17.2 | 93 |
| Blueberry latent virus | Amalgavirus | 0 | 0 | 0 | 0 | 25 | 25 | 0 | 0 | 6 | 25.5 | 3 |
| Blueberry scorch virus | Carlavirus | 0 | 25 | 0 | 0 | 0 | 25 | 0 | 0 | 6 | 42.3 | 147 |
| Cherry virus F | Fabavirus | 0 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 6 | 27.1 | 0 |
| Cycas necrotic stunt virus | Nepovirus | 0 | 25 | 25 | 0 | 0 | 0 | 0 | 0 | 6 | 34.3 | 23 |
| Raphanus sativus cryptic virus 2 | unclassified Partitiviridae | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 25 | 6 | 31.3 | 44 |
| Apple stem grooving virus | Capillovirus | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 3 | 24.0 | 0 |
| Citrus tatter leaf virus | Capillovirus | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 3 | 12.9 | 0 |
| Citrus virus A | Coguvirus | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 3 | 28.7 | 1 |
| Pyrus pyrifolia cryptic virus | Deltapartitivirus | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 0 | 3 | 12.4 | 0 |
| Privet leaf blotch-associated virus | Idaeovirus | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 3 | 77.9 | 44 |
| Grapevine-associated ilarvirus | Ilarvirus | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 10.1 | 36 |
| Actinidia virus X | Potexvirus | 0 | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 3 | 10.0 | 57 |
| Camellia ringspot-associated virus 1 | unclassified Betaflexiviridae | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 3 | 40.2 | 0 |
| Camellia ringspot-associated virus 2 | unclassified Betaflexiviridae | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 3 | 34.3 | 0 |
| Camellia ringspot-associated virus 3 | unclassified Betaflexiviridae | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 0 | 3 | 58.8 | 2 |
| Plant Virus | Genus | BC Site 1 | BC Site 2 | Average Frequency of Detection (%) * | Average Genome Coverage (%) * | Average VRPM* |
|---|---|---|---|---|---|---|
| Plant Tissue | Plant Tissue | |||||
| n = 2 | n = 2 | |||||
| Frequency (%) | ||||||
| Blueberry latent virus | Amalgavirus | 100 | 100 | 100 | 99.9 | 55,109 |
| Blueberry shock virus | Ilarvirus | 100 | 100 | 100 | 82.7 | 20,711 |
| Blueberry mosaic-associated virus | Ophiovirus | 0 | 100 | 50 | 99.6 | 625 |
| Blueberry scorch virus | Carlavirus | 0 | 100 | 50 | 66.3 | 900 |
| Bell pepper endornavirus | Alphaendornavirus | 0 | 50 | 25 | 25.2 | 1 |
| Brassica campestris chrysovirus 1 | Alphachrysovirus | 0 | 50 | 25 | 17.2 | 0 |
| Helianthus annuus alphaendornavirus | Alphaendornavirus | 0 | 50 | 25 | 100.0 | 338 |
| Plant Virus | Genus | ON Site 1 | Avearge Frequency of Detection (%) * | Average Genome Coverage (%) * | Average VRPM * | |||
|---|---|---|---|---|---|---|---|---|
| Bee Bread | Forager Bee | Hive Bee | Pollen | |||||
| n = 3 | n = 3 | n = 3 | n = 3 | |||||
| Frequency (%) | ||||||||
| Tomato ringspot virus | Nepovirus | 100 | 0 | 33 | 33 | 42 | 33.3 | 75 |
| Tobacco ringspot virus | Nepovirus | 67 | 0 | 67 | 0 | 33 | 22.9 | 61 |
| Raphanus sativus cryptic virus 3 | unclassified Partitiviridae | 67 | 0 | 0 | 33 | 25 | 40.2 | 22 |
| Brassica campestris chrysovirus 1 | Alphachrysovirus | 0 | 0 | 0 | 100 | 25 | 22.8 | 57 |
| Cherry virus A | Capillovirus | 33 | 0 | 0 | 33 | 17 | 16.8 | 60 |
| Prunus necrotic ringspot virus | Ilarvirus | 33 | 33 | 0 | 0 | 17 | 13.8 | 41 |
| Apple hammerhead viroid-like circular RNA | Pelamoviroid | 33 | 0 | 0 | 0 | 8 | 17.5 | 0 |
| Turnip vein-clearing virus | Tobamovirus | 0 | 0 | 33 | 0 | 8 | 11.0 | 67 |
| Tobacco streak virus | Ilarvirus | 0 | 0 | 33 | 0 | 8 | 10.7 | 128 |
| Plant Virus | Genus | ON Site 1 | Average Frequency of Detection (%) * | Average Genome Coverage (%) * | Average VRPM * |
|---|---|---|---|---|---|
| Plant Tissue | |||||
| n = 2 | |||||
| Frequency (%) | |||||
| Blueberry green mosaic-associated virus | Vitivirus | 100 | 100 | 98.9 | 34 |
| Blueberry latent virus | Amalgavirus | 100 | 100 | 99.7 | 61,635 |
| Blueberry mosaic-associated virus | Ophiovirus | 100 | 100 | 68.8 | 7 |
| Blueberry virus A | Unassigned (Closteroviridae family) | 100 | 100 | 61.9 | 922 |
| Bee Virus | Genus | Bee Bread | Forager Bee | Hive Bee | Pollen | Bee Bread | Forager Bee | Hive Bee | Pollen | Average Frequency of Detection (%) * | Average Genome Coverage (%) * | Average VRPM * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BC Site 1 | BC Site 2 | |||||||||||
| n = 4 | n = 4 | n = 4 | n = 4 | n = 4 | n = 4 | n = 4 | n = 4 | |||||
| Frequency (%) | ||||||||||||
| Black Queen Cell Virus | Triatovirus | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 81.1 | 1301 |
| Varroa destructor virus 1 | Iflavirus | 75 | 100 | 100 | 75 | 100 | 100 | 100 | 100 | 94 | 59.5 | 484 |
| Israel Acute Paralysis Virus | Aparavirus | 25 | 25 | 75 | 25 | 50 | 100 | 100 | 100 | 63 | 68.1 | 911 |
| Lake Sinai Virus | Sinaivirus | 0 | 100 | 100 | 0 | 0 | 100 | 100 | 75 | 59 | 88.6 | 5564 |
| Apis mellifera filamentous virus | Unclassified dsDNA virus | 0 | 75 | 25 | 0 | 0 | 50 | 75 | 25 | 31 | 46.8 | 16 |
| Hobart bee virus 1 | Unclassified Picornavirales | 25 | 50 | 50 | 0 | 0 | 0 | 50 | 25 | 25 | 26.1 | 0 |
| Sacbrood Virus | Iflavirus | 0 | 25 | 0 | 25 | 0 | 50 | 50 | 25 | 22 | 51.6 | 762 |
| Apis rhabdovirus 1 | Unclassified Rhabdoviridae | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 25 | 6 | 70.1 | 0 |
| Deformed Wing Virus | Iflavirus | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 25 | 6 | 65.3 | 4316 |
| Varroa destructor virus 3 | Iflavirus | 0 | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 3 | 12.7 | 0 |
| Bee Virus | Genus | ON Site 1 | Average Frequency of Detection (%) * | Average Genome Coverage (%) * | Average VRPM * | |||
|---|---|---|---|---|---|---|---|---|
| Bee Bread | Forager Bee | HIVE BEE | Pollen | |||||
| n = 3 | n = 3 | n = 3 | n = 3 | |||||
| Frequency (%) | ||||||||
| Black Queen Cell Virus | Triatovirus | 67 | 100 | 100 | 100 | 92 | 64.7 | 140 |
| Lake Sinai Virus | Sinaivirus | 0 | 100 | 33 | 0 | 42 | 58.2 | 626 |
| Sacbrood Bee Virus | Iflavirus | 0 | 67 | 33 | 0 | 25 | 42.6 | 158 |
| Apis mellifera filamentous virus | Unclassified dsDNA virus | 0 | 33 | 0 | 0 | 8 | 79.6 | 31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the Crown in Canada. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.; Vansia, R.; Phelan, J.; Lofano, A.; Smith, A.; Wang, A.; Bilodeau, G.J.; Pernal, S.F.; Guarna, M.M.; Rott, M.; et al. Area Wide Monitoring of Plant and Honey Bee (Apis mellifera) Viruses in Blueberry (Vaccinium corymbosum) Agroecosystems Facilitated by Honey Bee Pollination. Viruses 2023, 15, 1209. https://doi.org/10.3390/v15051209
Lee E, Vansia R, Phelan J, Lofano A, Smith A, Wang A, Bilodeau GJ, Pernal SF, Guarna MM, Rott M, et al. Area Wide Monitoring of Plant and Honey Bee (Apis mellifera) Viruses in Blueberry (Vaccinium corymbosum) Agroecosystems Facilitated by Honey Bee Pollination. Viruses. 2023; 15(5):1209. https://doi.org/10.3390/v15051209
Chicago/Turabian StyleLee, Eunseo, Raj Vansia, James Phelan, Andrea Lofano, Adam Smith, Aiming Wang, Guillaume J. Bilodeau, Stephen F. Pernal, M. Marta Guarna, Michael Rott, and et al. 2023. "Area Wide Monitoring of Plant and Honey Bee (Apis mellifera) Viruses in Blueberry (Vaccinium corymbosum) Agroecosystems Facilitated by Honey Bee Pollination" Viruses 15, no. 5: 1209. https://doi.org/10.3390/v15051209
APA StyleLee, E., Vansia, R., Phelan, J., Lofano, A., Smith, A., Wang, A., Bilodeau, G. J., Pernal, S. F., Guarna, M. M., Rott, M., & Griffiths, J. S. (2023). Area Wide Monitoring of Plant and Honey Bee (Apis mellifera) Viruses in Blueberry (Vaccinium corymbosum) Agroecosystems Facilitated by Honey Bee Pollination. Viruses, 15(5), 1209. https://doi.org/10.3390/v15051209