
Genetic Variability in the E6/E7 Region of Human Papillomavirus 16 in Women from Ecuador

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Molecular Analysis
2.3. Genetic Characterization and Phylogenetic Analyses
2.4. GenBank Accession Numbers
3. Results
3.1. Sample Characteristics
3.2. E6 and E7 Genetic Characterization
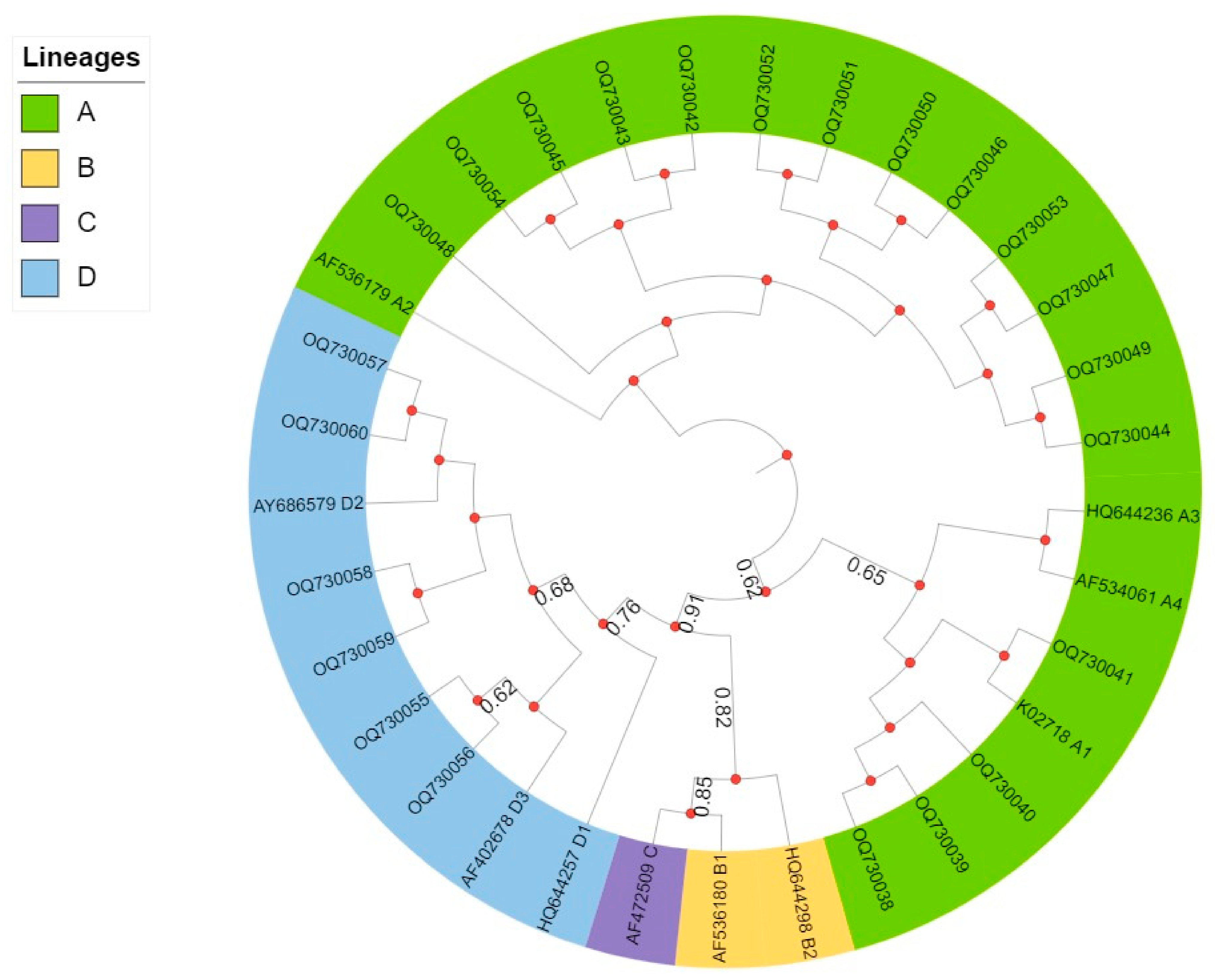
3.3. Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bermúdez, J.F.; Cabezas, C.P.; Rosales, K.L.; Gaitán, S.M.; Guerrero, M.X. Pruebas de detección del cáncer cervicouterino asociado al Virus del Papiloma Humano (VPH). Univ. Cienc. 2017, 12, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Laurent, J.S.; Luckett, R.; Feldman, S. HPV vaccination and the effects on rates of HPV-related cancers. Curr. Probl. Cancer 2018, 42, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Petca, A.; Borislavschi, A.; Zvanca, M.E.; Petca, R.C.; Sandru, F.; Dumitrascu, M.C. Non-sexual HPV transmission and role of vaccination for a better future. Exp. Ther. Med. 2020, 20, 186. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.W.; Mirabello, L. Human papillomavirus genomics: Understanding carcinogenicity. Tumour Virus Res. 2023, 15, 200258. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, N.; Bosch, F.X.; de Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K.V.; Snijders, P.J.; Meijer, C.J.; International Agency for Research on Cancer Multicenter Cervical Cancer Study Group. Epidemiologic classification of human papillomavirus types associated with cervical cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gultekin, M.; Ramirez, P.; Broutet, N.; Hutubessy, R. World Health Organization call for action to eliminate cervical cancer globally. Int. J. Gynecol. Cancer 2020, 4, 427. [Google Scholar] [CrossRef] [Green Version]
- Burd, E.M. Human papillomavirus and cervical cancer. Clin. Microbiol. Rev. 2003, 16, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Terai, M.; Fu, L.; Herrero, R.; DeSalle, R.; Burk, R.D. Diversifying selection in human papillomavirus type 16 lineages based on complete genome analyses. J. Virol. 2005, 79, 7014–7023. [Google Scholar] [CrossRef] [Green Version]
- Kukimoto, I.; Muramatsu, M. Genetic variations of human papillomavirus type 16: Implications for cervical carcinogenesis. Jpn. J. Infect Dis. 2015, 3, 175. [Google Scholar] [CrossRef] [Green Version]
- Sichero, L.; Sobrinho, J.S.; Villa, L.L. Oncogenic potential diverge among human papillomavirus type 16 natural variants. Virology 2012, 432, 127–132. [Google Scholar] [CrossRef]
- Mirabello, L.; Yeager, M.; Yu, K.; Clifford, G.M.; Xiao, Y.; Zhu, B.; Cullen, M.; Boland, J.F.; Wentzensen, N.; Nelson, C.W.; et al. HPV16 E7 Genetic Conservation Is Critical to Carcinogenesis. Cell 2017, 170, 1164–1174.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.; Chen, Z.; Reimers, L.; van Doorslaer, K.; Schiffman, M.; Desalle, R.; Herrero, R.; Yu, K.; Wacholder, S.; Wang, T.; et al. Sequence imputation of HPV16 genomes for genetic association studies. PLoS ONE 2011, 6, e21375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornet, I.; Gheit, T.; Franceschi, S.; Vignat, J.; Burk, R.D.; Sylla, B.S.; Tommasino, M.; Clifford, G. HPV Variant Study Group. Human papillomavirus type 16 genetic variants: Phylogeny and classification based on E6 and LCR. J. Virol. 2012, 12, 6855–6861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, T.; Manos, M.M.; Peto, J.; Greer, C.E.; Munoz, N.; Bosch, F.X.; Wheeler, C.M. Human papillomavirus type 16 sequence variation in cervical cancers: A worldwide perspective. J. Virol. 1997, 71, 2463–2472. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; DeSalle, R.; Schiffman, M.; Herrero, R.; Wood, C.E.; Ruiz, J.C.; Clifford, G.M.; Chan, P.K.S.; Burk, R.D. Niche adaptation and viral transmission of human papillomaviruses from archaic hominins to modern humans. PLoS Pathog. 2018, 14, e1007352. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.D.; Harari, A.; Chen, Z. Human papillomavirus genome variants. Virology 2013, 445, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Clifford, G.M.; Tenet, V.; Georges, D.; Alemany, L.; Pavón, M.A.; Chen, Z.; Yeager, M.; Cullen, M.; Boland, J.F.; Bass, S.; et al. Human papilloma-virus 16 sub-lineage dispersal and cervical cancer risk worldwide: Whole viral genome sequences from 7116 HPV16-positive women. Papillomavirus Res. 2019, 7, 67–74. [Google Scholar] [CrossRef]
- Bruni, L.; Alberto, G.; Serrano, B.; Mena, M.; Gómez, D.; Muñoz, J.; Bosch, F.X.; de Sanjosé, S. Human Papillomavirus and Related Diseases in Ecuador. [Summary Report] ICO/IARC, Information Centre on HPV and Cancer. 2019. Available online: www.hpvcentre.net (accessed on 10 June 2020).
- Leverone, R.B.; Regalado, J.G.; Giler, S.; Rosales, V. Situación Epidemiológica del Cáncer Cérvicouterino en el Ecuador. Rev. Venez. Oncol. 2021, 33, 69–78. [Google Scholar]
- Bedoya-Pilozo, C.H.; Medina-Magües, L.G.; Espinosa-García, M.; Sánchez, M.; Parrales-Valdiviezo, J.V.; Molina, D.; Ibarra, M.A.; Quimis-Ponce, M.; España, K.; Párraga-Macias, K.E.; et al. Molecular epidemiology and phylogenetic analysis of human papillomavirus infection in women with cervical lesions and cancer from the coastal region of Ecuador. Rev. Argent Microbiol. 2018, 50, 136–146. [Google Scholar] [CrossRef]
- Zehbe, I.; Tachezy, R.; Mytilineos, J.; Voglino, G.; Mikyskova, I.; Delius, H.; Marongiu, A.; Gissmann, L.; Wilander, E.; Tommasino, M. Human papillomavirus 16 E6 polymorphisms in cervical lesions from different European populations and their correlation with human leukocyte antigen class II haplotypes. Int. J. Cancer 2001, 94, 711–716. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Y.; Li, L.; Yu, G.; He, Y.; Zhang, Y. Analysis of mutations in the E6/E7 oncogenes and L1 gene of human papillomavirus 16 cervical cancer isolates from China. J. Gen. Virol. 2006, 87, 1181–1188. [Google Scholar] [CrossRef]
- Bedoya-Pilozo, C.H. Infección por Virus del Papiloma Humano, sus Variantes Genéticas y Factores Asociados a Lesiones Cervicouterinas en Mujeres de la Región Litoral del Ecuador. 2012–2015. Ph.D. Thesis, Instituto de Medicina Tropical “Pedro Kouri”, La Habana, Cuba, 2022. Unpublished. [Google Scholar]
- BLAST (Basic Local Alignment Search Tool). Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 2 March 2023).
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. Available online: http://www.iqtree.org/ (accessed on 2 March 2023). [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed] [Green Version]
- Bruni, L.; Albero, G.; Serrano, B.; Mena, M.; Collado, J.J.; Gómez, D.; Muñoz, J.; Bosch, F.X.; de Sanjosé, S.; ICO/IARC Information Centre on HPV and Cancer (HPV Information Centre). Human Papillomavirus and Related Diseases in the World. Summary Report. 10 March 2023. Available online: https://hpvcentre.net/datastatistics.php (accessed on 6 April 2023).
- Koenraad Van Doorslaer and others, The Papillomavirus Episteme: A major update to the papillomavirus sequence database. Nucleic Acids Res. 2017, 45, D499–D506. [CrossRef] [Green Version]
- Ghittoni, R.; Accardi, R.; Hasan, U.; Gheit, T.; Sylla, B.; Tommasino, M. The biological properties of E6 and E7 oncoproteins from human papillomaviruses. Virus Genes 2010, 40, 1–13. [Google Scholar] [CrossRef]
- Lee, C.W.; Bae, J.H.; Lee, S.J.; Ho, E.M.; Lee, I.H.; Park, Y.G.; Park, J.S. Distribution of human papillomavirus type 16 E6 and E7 gene variants in the progression of cervical dysplasia in Korean women. J. Obstet. Gynaecol. Res. 2011, 37, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Antaño-Arias, R.; Del Moral-Hernández, O.; Ortiz-Ortiz, J.; Alarcón-Romero, L.D.C.; Navor-Hernández, J.A.; Leyva-Vázquez, M.A.; Jiménez-López, M.A.; Organista-Nava, J.; Illades-Aguiar, B. E6/E7 Variants of Human Papillomavirus 16 Associated with Cervical Carcinoma in Women in Southern Mexico. Pathogens 2021, 10, 773. [Google Scholar] [CrossRef] [PubMed]
- Badano, I.; Totaro, M.E.; Culasso, A.C.; Sanabria, D.J.; Schurr, T.G.; Balette, I.C.; Roisman, A.; Basiletti, J.; Picconi, M.A.; Cam-pos, R.H.; et al. Genetic characterization and clinical implications of human papillomavirus type 16 (HPV16) variants from northeastern Argentina. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Ge-Netics Infect. Dis. 2015, 29, 103–109. [Google Scholar] [CrossRef]
- Mejía, L.; Muñoz, D.; Trueba, G.; Tinoco, L.; Zapata, S. Prevalence of human papillomavirus types in cervical cancer-ous and precancerous lesions of Ecuadorian women. J. Med. Virol. 2016, 88, 144–152. [Google Scholar] [CrossRef]
- Flores-Espinoza, R.; Paz-Cruz, E.; Ruiz-Pozo, V.A.; Lopez-Carrera, M.; Cabrera-Andrade, A.; Gusmão, L.; Burgos, G. Investigating genetic diversity in admixed populations from Ecuador. Am. J. Phys. Anthropol. 2021, 176, 109–119. [Google Scholar] [CrossRef]
- López-Revilla, R.; Pineda, M.A.; Ortiz-Valdez, J.; Sánchez-Garza, M.; Riego, L. Human papillomavirus type 16 variants in cervical intraepithelial neoplasia and invasive carcinoma in San Luis Potosí City, Mexico. Infect. Agents Cancer 2009, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobar-Escamilla, N.; González-Martínez, B.E.; Araiza-Rodríguez, A.; Fragoso-Fonseca, D.E.; Pedroza-Torres, A.; Landa-Flores, M.G.; Garcés-Ayala, F.; Mendieta-Condado, E.; Díaz-Quiñonez, J.A.; Castro-Escarpulli, G.; et al. Mutational landscape and intra-host diversity of human papillomavirus type 16 long control region and E6 variants in cervical samples. Arch. Virol. 2019, 164, 2953–2961. [Google Scholar] [CrossRef] [PubMed]
- Gheit, T.; Simoes, R.T.; Tommasino, M.; Donadi, E.A.; Gonçalves, M.A. HPV16 variants in squamous intraepithelial lesions in human immunodeficiency virus-negative and -positive Brazilian women. Viral Immunol. 2006, 19, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Totaro, M.E.; Gili, J.A.; Liotta, D.J.; Schurr, T.G.; Picconi, M.A.; Badano, I. Genetic variation in the E6 and E7 genes of human papillomavirus type 16 in northeastern Argentina. J. Med. Virol. 2022, 94, 745–751. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, C.M.; Bravo, I.G.; Santiago e Souza, N.C.; Genta, M.L.; Fregnani, J.H.; Tacla, M.; Carvalho, J.P.; Longatto-Filho, A.; Levi, J.E. High-level of viral genomic diversity in cervical cancers: A Brazilian study on human papillomavirus type 16. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2015, 34, 44–51. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| E6 | 145 | 183 | 286 | 289 | 335 | 350 | 532 | Amino Acids | n (%) | Cervical Lesions | Lineage | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genome Position | ||||||||||||||
| Ref SNPs | G | T | T | A | C | T | A | Ref | ASCUS | LSIL | HSIL | CC | ||
| OQ730038 | - | - | - | - | - | - | - | Ref | 4 (17.4) | 1 | 1 | 1 | 1 | A |
| OQ730042 | - | - | - | - | - | G | - | L83V | 13 (56.5) | 2 | - | 8 | 2 | A |
| OQ730057 | T | - | A | G | T | G | G | Q14H–H78Y–L83V | 4 (17.4) | 1 | - | 3 | - | D |
| OQ730055 | T | G | A | G | T | G | G | Q14H–I27R–H78Y–L83V | 2 (8.7) | - | - | - | 2 | D |
| Total | 23 (100) | 5 | 1 | 12 | 5 | |||||||||
| 29 | 1 | 2 | 1 | 1 | ||||||||||
| E7 | 678 | 732 | 789 | 795 | 828 | Amino Acids | n (%) | Cervical Lesions | Lineage | |||||
| Genome Position | ||||||||||||||
| Ref | T | T | T | T | T | Ref | ASCUS | LSIL | HSIL | CC | ||||
| OQ730063 | - | - | - | - | - | Ref | 13 (61.9) | 2 | 2 | 4 | 5 | A | ||
| OQ730062 | C | - | - | - | - | Ref | 2 (9.5) | 1 | - | 1 | - | A | ||
| OQ730076 | - | - | C | - | C | Ref | 1 (4.8) | 1 | - | - | - | A | ||
| OQ730077 | - | C | C | G | - | Ref | 5 (23.8) | 2 | 1 | 2 | - | D | ||
| Total | 21 (100) | 6 | 3 | 7 | 5 | |||||||||
| 29 | 0 | 0 | 5 | 2 | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhingre, A.; Bedoya-Pilozo, C.; Gutiérrez-Pallo, D.; Badano, I.; Herrera-Yela, A.; Salazar, Z.; Alarcón, D.; Argüello-Bravo, N.; Espinoza, M.; Ponce, P.; et al. Genetic Variability in the E6/E7 Region of Human Papillomavirus 16 in Women from Ecuador. Viruses 2023, 15, 1393. https://doi.org/10.3390/v15061393
Zhingre A, Bedoya-Pilozo C, Gutiérrez-Pallo D, Badano I, Herrera-Yela A, Salazar Z, Alarcón D, Argüello-Bravo N, Espinoza M, Ponce P, et al. Genetic Variability in the E6/E7 Region of Human Papillomavirus 16 in Women from Ecuador. Viruses. 2023; 15(6):1393. https://doi.org/10.3390/v15061393
Chicago/Turabian StyleZhingre, Alicia, César Bedoya-Pilozo, Diana Gutiérrez-Pallo, Inés Badano, Andrés Herrera-Yela, Zoila Salazar, Damaris Alarcón, Natali Argüello-Bravo, Maylen Espinoza, Patricio Ponce, and et al. 2023. "Genetic Variability in the E6/E7 Region of Human Papillomavirus 16 in Women from Ecuador" Viruses 15, no. 6: 1393. https://doi.org/10.3390/v15061393
APA StyleZhingre, A., Bedoya-Pilozo, C., Gutiérrez-Pallo, D., Badano, I., Herrera-Yela, A., Salazar, Z., Alarcón, D., Argüello-Bravo, N., Espinoza, M., Ponce, P., Soto, Y., & Carrazco-Montalvo, A. (2023). Genetic Variability in the E6/E7 Region of Human Papillomavirus 16 in Women from Ecuador. Viruses, 15(6), 1393. https://doi.org/10.3390/v15061393