

Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Compounds
2.2. Plasmids and Pseudovirus Production
2.3. Luciferase Assay on Pseudovirus Infection
2.4. Cytotoxicity Assay
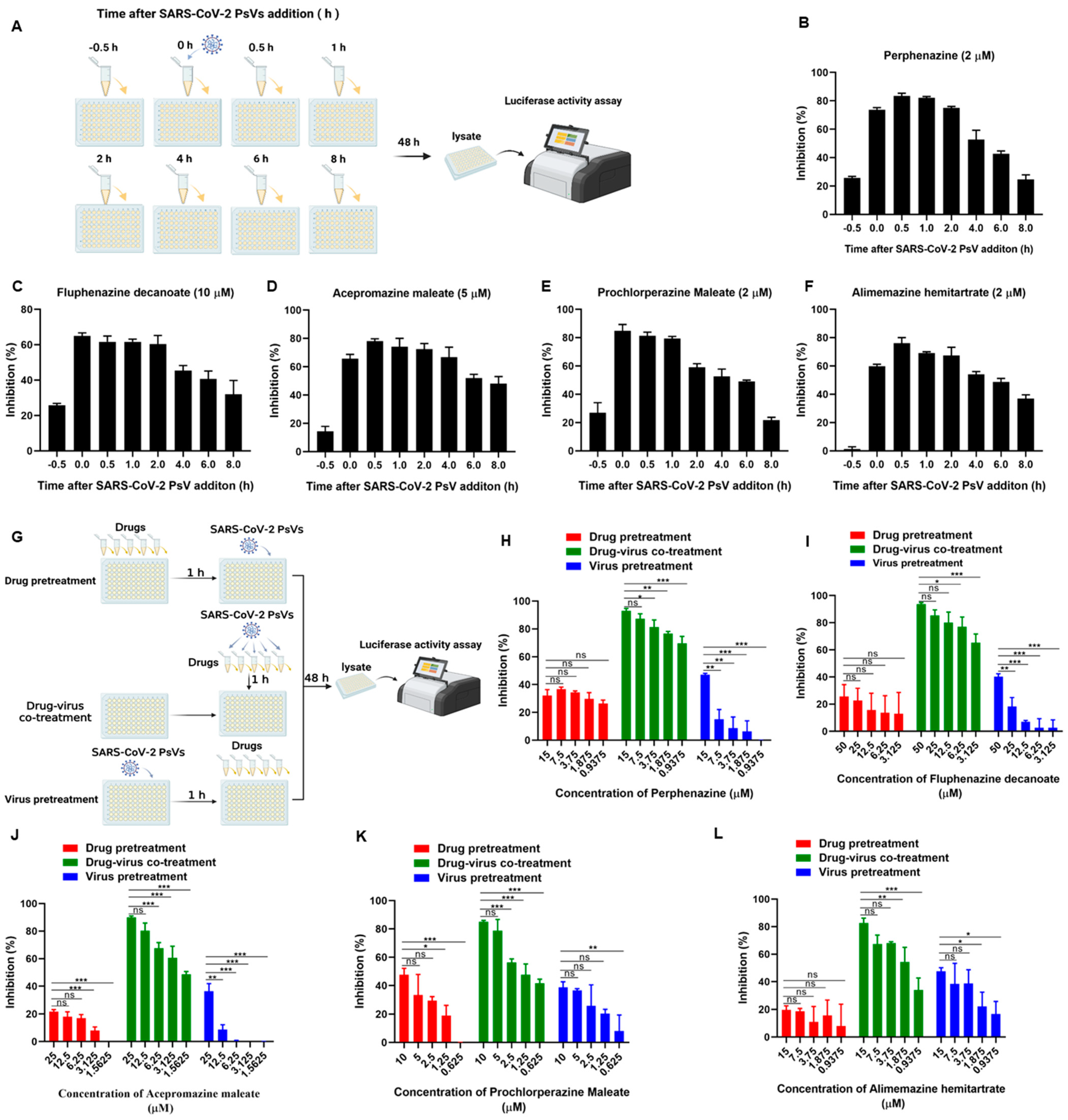
2.5. Time-of-Addition Assay
2.6. Drug or Virus Pretreatment Assay
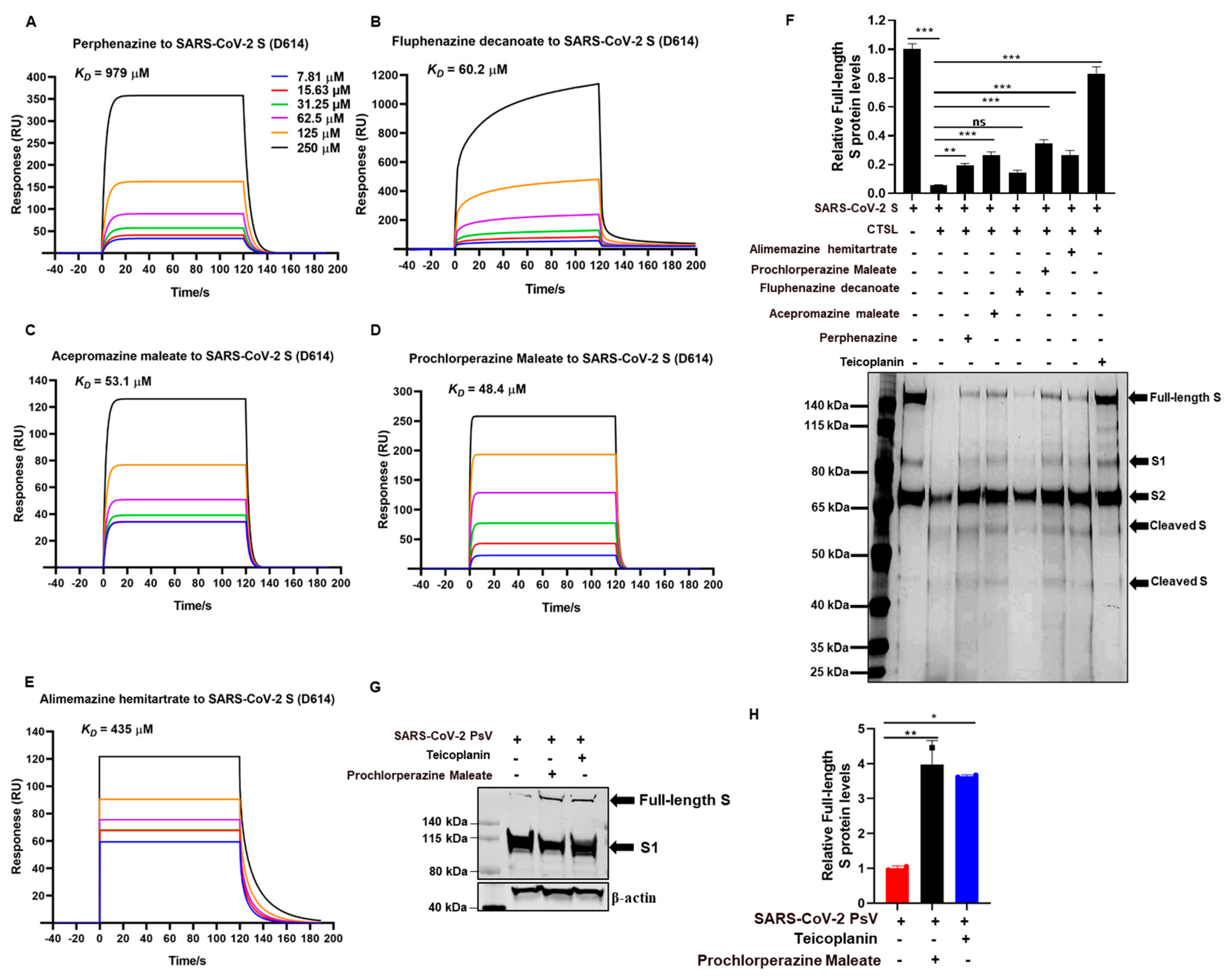
2.7. Surface Plasmon Resonance (SPR) Binding Analysis
2.8. CTSL-Mediated the Cleavage of SARS-CoV-2 Spike Protein In Vitro
2.9. The Cleavage of SARS-CoV-2 Spike Protein
2.10. Antiviral Experiments In Vivo
2.11. Real-Time Quantitative PCR (RT-qPCR)
2.12. Statistical Analysis
3. Results
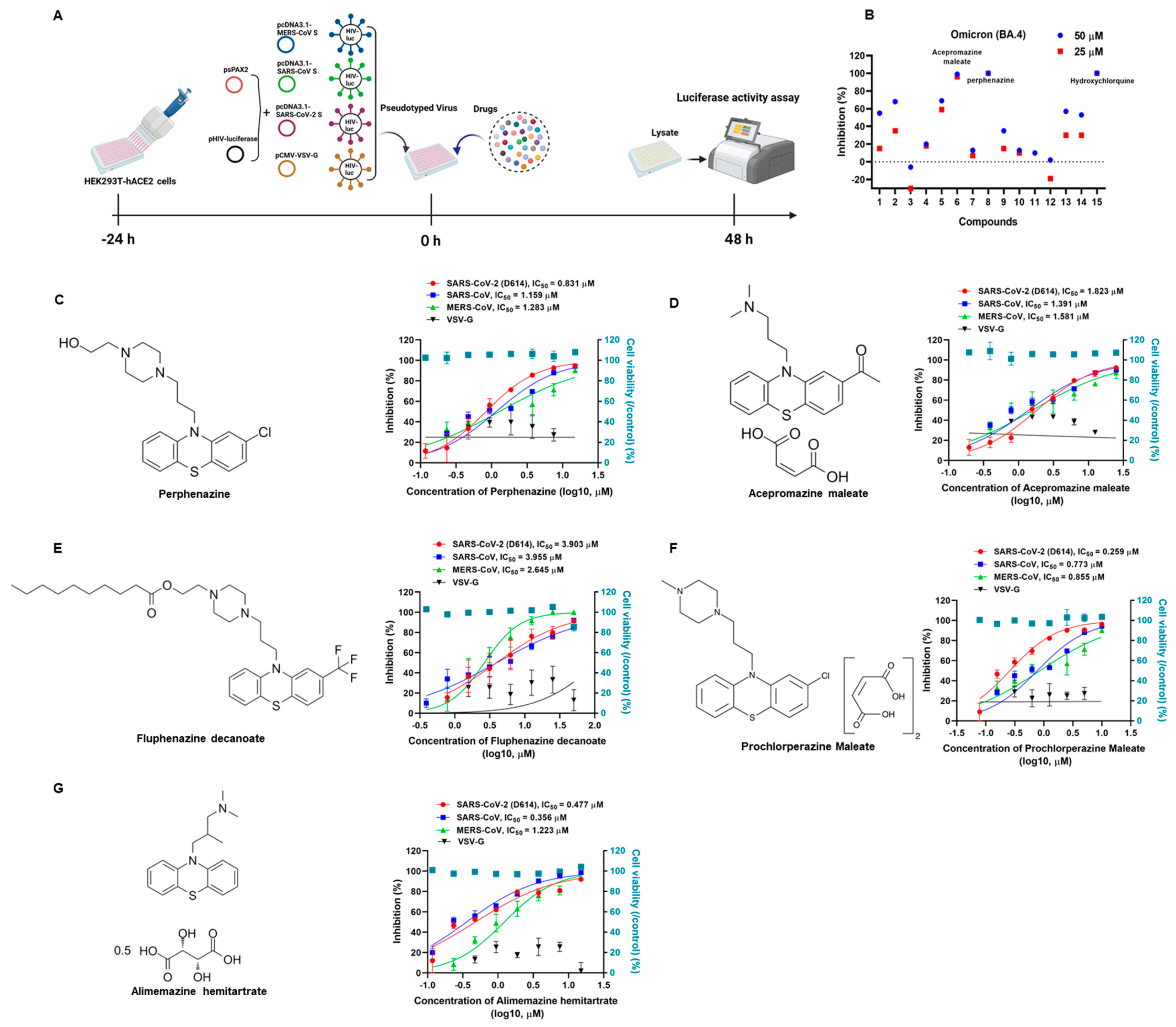
3.1. Phenothiazines Show Potent Broad-Spectrum Antiviral Activities
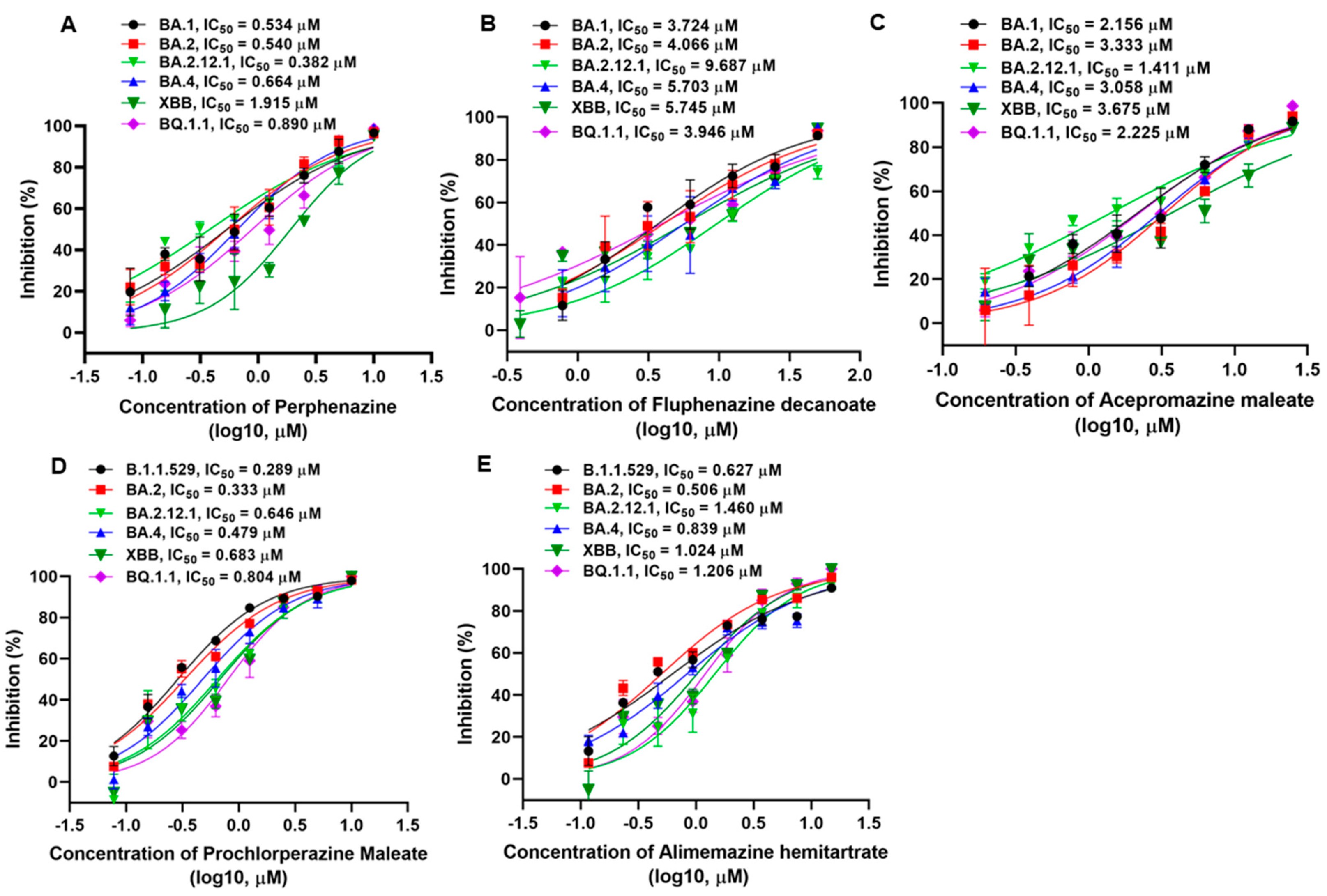
3.2. Phenothiazines Inhibit the Entry of Multiple SARS-CoV-2 Variants
3.3. Phenothiazines Predominantly Inhibit SARS-CoV-2 PsV Infection at the Early Stage
3.4. Phenothiazines Bind to S Protein and Interfere S Protein Cleavage
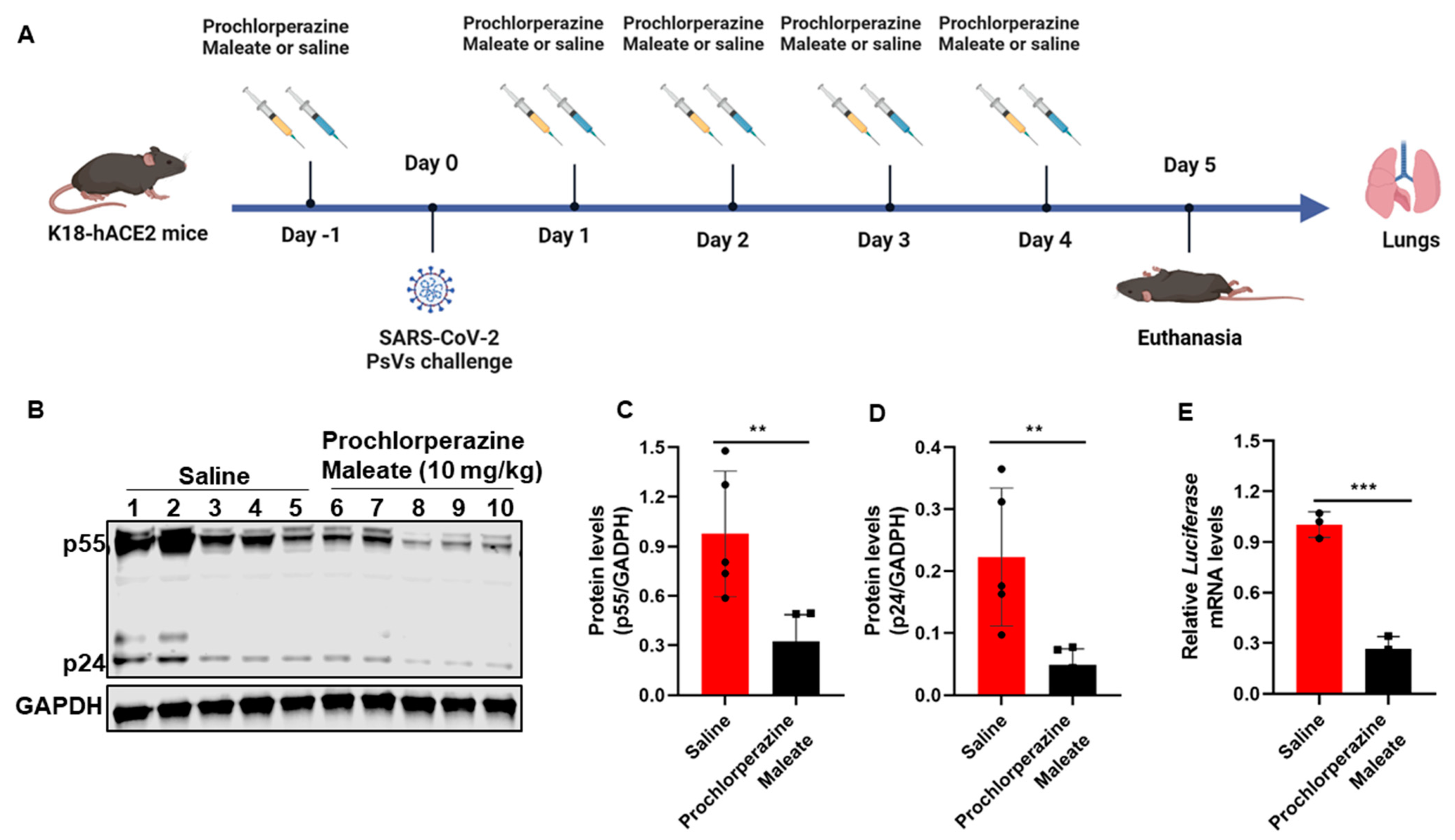
3.5. Prochlorperazine Maleate Inhibits SARS-CoV-2 PsV Infection in K18-hACE2 Mice

4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Li, X.; Li, T.; Zhang, S.; Wang, L.; Wu, X.; Liu, J. The genetic sequence, origin, and diagnosis of SARS-CoV-2. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1629–1635. [Google Scholar] [CrossRef]
- Telenti, A.; Arvin, A.; Corey, L.; Corti, D.; Diamond, M.S.; García-Sastre, A.; Garry, R.F.; Holmes, E.C.; Pang, P.S.; Virgin, H.W. After the pandemic: Perspectives on the future trajectory of COVID-19. Nature 2021, 596, 495–504. [Google Scholar] [CrossRef]
- Brady, D.K.; Gurijala, A.R.; Huang, L.; Hussain, A.A.; Lingan, A.L.; Pembridge, O.G.; Ratangee, B.A.; Sealy, T.T.; Vallone, K.T.; Clements, T.P. A guide to COVID-19 antiviral therapeutics: A summary and perspective of the antiviral weapons against SARS-CoV-2 infection. Febs. J. 2022. [Google Scholar] [CrossRef] [PubMed]
- Reina, J.; Iglesias, C. Nirmatrelvir plus ritonavir (Paxlovid) a potent SARS-CoV-2 3CLpro protease inhibitor combination. Rev. Esp. Quim. 2022, 35, 236–240. [Google Scholar] [CrossRef]
- Wen, W.; Chen, C.; Tang, J.; Wang, C.; Zhou, M.; Cheng, Y.; Zhou, X.; Wu, Q.; Zhang, X.; Feng, Z.; et al. Efficacy and safety of three new oral antiviral treatment (molnupiravir, fluvoxamine and Paxlovid) for COVID-19: A meta-analysis. Ann. Med. 2022, 54, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Vitiello, A.; Troiano, V.; La Porta, R. What will be the role of molnupiravir in the treatment of COVID-19 infection? Drugs Ther. Perspect. 2021, 37, 579–580. [Google Scholar] [CrossRef] [PubMed]
- WHO Solidarity Trial Consortium. Remdesivir and three other drugs for hospitalised patients with COVID-19: Final results of the WHO Solidarity randomised trial and updated meta-analyses. Lancet 2022, 399, 1941–1953. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of molnupiravir-induced SARS-CoV-2 mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef]
- Gandhi, S.; Klein, J.; Robertson, A.J.; Peña-Hernández, M.A.; Lin, M.J.; Roychoudhury, P.; Lu, P.; Fournier, J.; Ferguson, D.; Mohamed Bakhash, S.A.K.; et al. De novo emergence of a remdesivir resistance mutation during treatment of persistent SARS-CoV-2 infection in an immunocompromised patient: A case report. Nat. Commun. 2022, 13, 1547. [Google Scholar] [CrossRef]
- Najjar-Debbiny, R.; Gronich, N.; Weber, G.; Khoury, J.; Amar, M.; Stein, N.; Goldstein, L.H.; Saliba, W. Effectiveness of Paxlovid in Reducing Severe Coronavirus Disease 2019 and Mortality in High-Risk Patients. Clin. Infect. Dis. 2023, 76, e342–e349. [Google Scholar] [CrossRef]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simón-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with COVID-19. N. Engl. J. Med. 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Sacco, M.D.; Hu, Y.; Gongora, M.V.; Meilleur, F.; Kemp, M.T.; Zhang, X.; Wang, J.; Chen, Y. The P132H mutation in the main protease of Omicron SARS-CoV-2 decreases thermal stability without compromising catalysis or small-molecule drug inhibition. Cell Res. 2022, 32, 498–500. [Google Scholar] [CrossRef]
- Mótyán, J.A.; Mahdi, M.; Hoffka, G.; Tőzsér, J. Potential Resistance of SARS-CoV-2 Main Protease (Mpro) against Protease Inhibitors: Lessons Learned from HIV-1 Protease. Int. J. Mol. Sci. 2022, 23, 3507. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Lan, Q.; Wang, L.; Jiao, F.; Lu, L.; Xia, S.; Jiang, S. Pan-coronavirus fusion inhibitors to combat COVID-19 and other emerging coronavirus infectious diseases. J. Med. Virol. 2023, 95, e28143. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Yan, L.; Xu, W.; Agrawal, A.S.; Algaissi, A.; Tseng, C.K.; Wang, Q.; Du, L.; Tan, W.; Wilson, I.A.; et al. A pan-coronavirus fusion inhibitor targeting the HR1 domain of human coronavirus spike. Sci. Adv. 2019, 5, eaav4580. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Pan, T.; Huang, F.; Ying, R.; Liu, J.; Fan, H.; Zhang, J.; Liu, W.; Lin, Y.; Yuan, Y.; et al. Glycopeptide Antibiotic Teicoplanin Inhibits Cell Entry of SARS-CoV-2 by Suppressing the Proteolytic Activity of Cathepsin L. Front. Microbiol. 2022, 13, 884034. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Pan, T.; Zhang, J.; Li, Q.; Zhang, X.; Bai, C.; Huang, F.; Peng, T.; Zhang, J.; Liu, C.; et al. Glycopeptide Antibiotics Potently Inhibit Cathepsin L in the Late Endosome/Lysosome and Block the Entry of Ebola Virus, Middle East Respiratory Syndrome Coronavirus (MERS-CoV), and Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV). J. Biol. Chem. 2016, 291, 9218–9232. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.M.; Zhu, Y.; Zhang, L.; Zhong, G.; Tai, L.; Liu, S.; Yin, G.; Lu, J.; He, Q.; Li, M.J.; et al. Novel cleavage sites identified in SARS-CoV-2 spike protein reveal mechanism for cathepsin L-facilitated viral infection and treatment strategies. Cell Discov. 2022, 8, 53. [Google Scholar] [CrossRef]
- Song, W.; Gui, M.; Wang, X.; Xiang, Y. Cryo-EM structure of the SARS coronavirus spike glycoprotein in complex with its host cell receptor ACE2. PLoS Pathog. 2018, 14, e1007236. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.M.; Yang, W.L.; Yang, F.Y.; Zhang, L.; Huang, W.J.; Hou, W.; Fan, C.F.; Jin, R.H.; Feng, Y.M.; Wang, Y.C.; et al. Cathepsin L plays a key role in SARS-CoV-2 infection in humans and humanized mice and is a promising target for new drug development. Signal Transduct. Target. Ther. 2021, 6, 134. [Google Scholar] [CrossRef]
- Simmons, G.; Zmora, P.; Gierer, S.; Heurich, A.; Pöhlmann, S. Proteolytic activation of the SARS-coronavirus spike protein: Cutting enzymes at the cutting edge of antiviral research. Antivir. Res. 2013, 100, 605–614. [Google Scholar] [CrossRef]
- McCray, P.B., Jr.; Pewe, L.; Wohlford-Lenane, C.; Hickey, M.; Manzel, L.; Shi, L.; Netland, J.; Jia, H.P.; Halabi, C.; Sigmund, C.D.; et al. Lethal infection of K18-hACE2 mice infected with severe acute respiratory syndrome coronavirus. J. Virol. 2007, 81, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Muralidar, S.; Gopal, G.; Visaga Ambi, S. Targeting the viral-entry facilitators of SARS-CoV-2 as a therapeutic strategy in COVID-19. J. Med. Virol. 2021, 93, 5260–5276. [Google Scholar] [CrossRef]
- Pu, J.; Zhou, J.T.; Liu, P.; Yu, F.; He, X.; Lu, L.; Jiang, S. Viral Entry Inhibitors Targeting Six-Helical Bundle Core against Highly Pathogenic Enveloped Viruses with Class I Fusion Proteins. Curr. Med. Chem. 2022, 29, 700–718. [Google Scholar] [CrossRef]
- Li, L.; He, L.; Tan, S.; Guo, X.; Lu, H.; Qi, Z.; Pan, C.; An, X.; Jiang, S.; Liu, S. 3-hydroxyphthalic anhydride-modified chicken ovalbumin exhibits potent and broad anti-HIV-1 activity: A potential microbicide for preventing sexual transmission of HIV-1. Antimicrob. Agents Chemother. 2010, 54, 1700–1711. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Duan, J.; Qiu, J.; Yu, F.; Che, X.; Jiang, S.; Li, L. 3-hydroxyphthalic anhydride-modified human serum albumin as a microbicide candidate against HIV type 1 entry by targeting both viral envelope glycoprotein gp120 and cellular receptor CD4. AIDS Res. Hum. Retroviruses 2013, 29, 1455–1464. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Li, P.; Xu, W.; Liu, Z.; Wang, C.; Wang, Q.; Tang, J.; Li, W.; Lu, L.; Jiang, S. Chemically Modified Bovine β-Lactoglobulin as a Broad-Spectrum Influenza Virus Entry Inhibitor with the Potential to Combat Influenza Outbreaks. Viruses 2022, 14, 55. [Google Scholar] [CrossRef]
- Lu, L.; Yang, X.; Li, Y.; Jiang, S. Chemically modified bovine beta-lactoglobulin inhibits human papillomavirus infection. Microbes Infect. 2013, 15, 506–510. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, Q.; Jia, R.; Xia, S.; Li, Y.; Liu, Q.; Xu, W.; Xu, J.; Du, L.; Lu, L.; et al. Intranasal administration of maleic anhydride-modified human serum albumin for pre-exposure prophylaxis of respiratory syncytial virus infection. Viruses 2015, 7, 798–819. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Du, L.; Ma, C.; Li, Y.; Li, L.; Poon, V.K.; Wang, L.; Yu, F.; Zheng, B.J.; Jiang, S.; et al. A safe and convenient pseudovirus-based inhibition assay to detect neutralizing antibodies and screen for viral entry inhibitors against the novel human coronavirus MERS-CoV. Virol. J. 2013, 10, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, T.; Qiu, J.; Niu, X.; Ma, Q.; Zhou, C.; Chen, P.; Zhang, Q.; Chen, M.; Yang, Z.; Liu, S.; et al. 3-Hydroxyphthalic Anhydride-Modified Chicken Ovalbumin as a Potential Candidate Inhibits SARS-CoV-2 Infection by Disrupting the Interaction of Spike Protein With Host ACE2 Receptor. Front. Pharmacol. 2020, 11, 603830. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, M.; Takashima, A.; Ono, C.; Okamoto, T.; Iwasaki, M. Phenothiazines inhibit SARS-CoV-2 cell entry via a blockade of spike protein binding to neuropilin-1. Antivir. Res. 2023, 209, 105481. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yu, D.; Yan, H.; Chong, H.; He, Y. Design of Potent Membrane Fusion Inhibitors against SARS-CoV-2, an Emerging Coronavirus with High Fusogenic Activity. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef]
- Candurra, N.A.; Maskin, L.; Damonte, E.B. Inhibition of arenavirus multiplication in vitro by phenotiazines. Antivir. Res. 1996, 31, 149–158. [Google Scholar] [CrossRef]
- Hewlett, I.; Lee, S.; Molnar, J.; Foldeak, S.; Pine, P.S.; Weaver, J.L.; Aszalos, A. Inhibition of HIV infection of H9 cells by chlorpromazine derivatives. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol 1997, 15, 16–20. [Google Scholar] [CrossRef]
- Ochiai, H.; Kurokawa, M.; Niwayama, S. Influence of trifluoperazine on the late stage of influenza virus infection in MDCK cells. Antivir. Res. 1991, 15, 149–160. [Google Scholar] [CrossRef]
- Piccini, L.E.; Castilla, V.; Damonte, E.B. Inhibition of dengue virus infection by trifluoperazine. Arch. Virol. 2022, 167, 2203–2212. [Google Scholar] [CrossRef]
- Ruiz de Pellón Santamaría, Á. Psychosis Treatment During COVID-19 Pandemic and the Potential Role of Phenothiazines: A Call for Research Studies. J. Clin. Psychopharmacol. 2020, 40, 641–642. [Google Scholar] [CrossRef]
- Amaral, L.; Viveiros, M.; Molnar, J. Antimicrobial activity of phenothiazines. In Vivo 2004, 18, 725–731. [Google Scholar]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef]
- Koch, J.; Uckeley, Z.M.; Doldan, P.; Stanifer, M.; Boulant, S.; Lozach, P.Y. TMPRSS2 expression dictates the entry route used by SARS-CoV-2 to infect host cells. Embo J. 2021, 40, e107821. [Google Scholar] [CrossRef]
- Forrestall, K.L.; Burley, D.E.; Cash, M.K.; Pottie, I.R.; Darvesh, S. Phenothiazines as dual inhibitors of SARS-CoV-2 main protease and COVID-19 inflammation. Can. J. Chem. 2021, 99, 801–811. [Google Scholar] [CrossRef]
- Machado-Vieira, R.; Quevedo, J.; Shahani, L.; Soares, J.C. Convergent evidence for the antiviral effects of several FDA-approved phenothiazine antipsychotics against SARS-CoV-2 and other coronaviruses. Braz. J. Psychiatry 2021, 43, 462–464. [Google Scholar] [CrossRef]
- Plaze, M.; Attali, D.; Petit, A.C.; Blatzer, M.; Simon-Loriere, E.; Vinckier, F.; Cachia, A.; Chrétien, F.; Gaillard, R. Repurposing chlorpromazine to treat COVID-19: The reCoVery study. Encephale 2020, 46, 169–172. [Google Scholar] [CrossRef]
- Lu, J.; Hou, Y.; Ge, S.; Wang, X.; Wang, J.; Hu, T.; Lv, Y.; He, H.; Wang, C. Screened antipsychotic drugs inhibit SARS-CoV-2 binding with ACE2 in vitro. Life Sci. 2021, 266, 118889. [Google Scholar] [CrossRef]
- Varga, B.; Csonka, Á.; Csonka, A.; Molnár, J.; Amaral, L.; Spengler, G. Possible Biological and Clinical Applications of Phenothiazines. Anticancer. Res. 2017, 37, 5983–5993. [Google Scholar] [CrossRef] [Green Version]
- Jaszczyszyn, A.; Gąsiorowski, K.; Świątek, P.; Malinka, W.; Cieślik-Boczula, K.; Petrus, J.; Czarnik-Matusewicz, B. Chemical structure of phenothiazines and their biological activity. Pharmacol. Rep. 2012, 64, 16–23. [Google Scholar] [CrossRef]
- Galea, M.; Agius, M.; Vassallo, N. Neurological manifestations and pathogenic mechanisms of COVID-19. Neurol. Res. 2022, 44, 571–582. [Google Scholar] [CrossRef]
- Jaffry, M.; Faiz, I.; Jaffry, K.; Souayah, N. Neurological Manifestations of SARS-CoV-2 Infection and the Role of Complement Activation. US Neurol. 2022. [Google Scholar] [CrossRef]
- Chen, J.; Li, S.; Lei, Z.; Tang, Q.; Mo, L.; Zhao, X.; Xie, F.; Zi, D.; Tan, J. Inhibition of SARS-CoV-2 pseudovirus invasion by ACE2 protecting and Spike neutralizing peptides: An alternative approach to COVID19 prevention and therapy. Int. J. Biol. Sci. 2021, 17, 2957–2969. [Google Scholar] [CrossRef] [PubMed]
- Simanjuntak, Y.; Liang, J.-J.; Lee, Y.-L.; Lin, Y.-L. Repurposing of Prochlorperazine for Use Against Dengue Virus Infection. J. Infect. Dis. 2014, 211, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Scovino, A.M.; Dahab, E.C.; Vieira, G.F.; Freire-de-Lima, L.; Freire-de-Lima, C.G.; Morrot, A. SARS-CoV-2’s Variants of Concern: A Brief Characterization. Front. Immunol. 2022, 13, 834098. [Google Scholar] [CrossRef] [PubMed]
- Kaufer, A.M.; Theis, T.; Lau, K.A.; Gray, J.L.; Rawlinson, W.D. Laboratory biosafety measures involving SARS-CoV-2 and the classification as a Risk Group 3 biological agent. Pathology 2020, 52, 790–795. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (μM) | CC50 (μM) | SI b |
|---|---|---|---|
| Perphenazine | 0.831 ± 0.120 | 18.120 ± 1.450 | 21.805 |
| Acepromazine maleate | 1.823 ± 0.230 | 93.300 ± 11.520 | 51.179 |
| Fluphenazine decanoate | 3.903 ± 1.090 | 61.570 ± 28.840 | 15.775 |
| Prochlorperazine maleate | 0.259 ± 0.048 | 85.160 ± 13.915 | 328.803 |
| Alimemazine hemitartrate | 0.477 ± 0.124 | 53.390 ± 10.945 | 111.929 |
| SARS-CoV-2 Variants | IC50 (μM, Mean ± SD) | ||||
|---|---|---|---|---|---|
| Perphenazine | Fluphenazine Decanoate | Acepromazine Maleate | Prochlorperazine Maleate | Alimemazine Hemitartrate | |
| B.1.1.7 (Alpha) | 1.315 ± 0.283 | 2.680 ± 0.370 | 2.580 ± 0.494 | 0.526 ± 0.103 | 0.580 ± 0.084 |
| B.1.351 (Beta) | 1.215 ± 0.225 | 1.736 ± 0.518 | 3.152 ± 0.602 | 0.293 ± 0.027 | 0.293 ± 0.059 |
| P.1 (Gamma) | 0.487 ± 0.169 | 6.214 ± 1.211 | 1.719 ± 0.348 | 0.762 ± 0.100 | 1.526 ± 0.339 |
| B.1.429 (Epsilon) | 0.802 ± 0.125 | 1.200 ± 0.425 | 4.605 ± 1.068 | 0.113 ± 0.034 | 0.342 ± 0.103 |
| B.1.526 (lota) | 0.315 ± 0.042 | 4.525 ± 2.615 | 1.298 ± 0.196 | 0.31 ± 0.045 | 1.040 ± 0.574 |
| B.1.617.1 (Kappa) | 0.343 ± 0.075 | 3.957 ± 0.762 | 0.661 ± 0.161 | 0.249 ± 0.057 | 0.609 ± 0.082 |
| B.1.617.2 (Delta) | 0.903 ± 0.211 | 9.318 ± 3.236 | 2.496 ± 0.583 | 0.609 ± 0.128 | 1.776 ± 0.660 |
| C.37 (Lambda) | 0.482 ± 0.059 | 2.270 ± 0.419 | 1.993 ± 0.576 | 0.423 ± 0.096 | 0.342 ± 0.097 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, T.; Xiao, S.; Wu, Z.; Lv, X.; Liu, S.; Hu, M.; Li, G.; Li, P.; Ma, X. Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein. Viruses 2023, 15, 1666. https://doi.org/10.3390/v15081666
Liang T, Xiao S, Wu Z, Lv X, Liu S, Hu M, Li G, Li P, Ma X. Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein. Viruses. 2023; 15(8):1666. https://doi.org/10.3390/v15081666
Chicago/Turabian StyleLiang, Taizhen, Shiqi Xiao, Ziyao Wu, Xi Lv, Sen Liu, Meilin Hu, Guojie Li, Peiwen Li, and Xiancai Ma. 2023. "Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein" Viruses 15, no. 8: 1666. https://doi.org/10.3390/v15081666
APA StyleLiang, T., Xiao, S., Wu, Z., Lv, X., Liu, S., Hu, M., Li, G., Li, P., & Ma, X. (2023). Phenothiazines Inhibit SARS-CoV-2 Entry through Targeting Spike Protein. Viruses, 15(8), 1666. https://doi.org/10.3390/v15081666