
The Antiviral Potential of AdipoRon, an Adiponectin Receptor Agonist, Reveals the Ability of Zika Virus to Deregulate Adiponectin Receptor Expression

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus, Cell Culture and Reagents
2.2. Lactate Dehydrogenase Assay (LDH)
2.3. Neutral Red Assay
2.4. Cell Infection and Treatments with AdipoRon or Adiponectin
2.5. Flow Cytometry and Antibody
2.6. RNA Extraction and qRT-PCR
2.7. Statistical Analyses
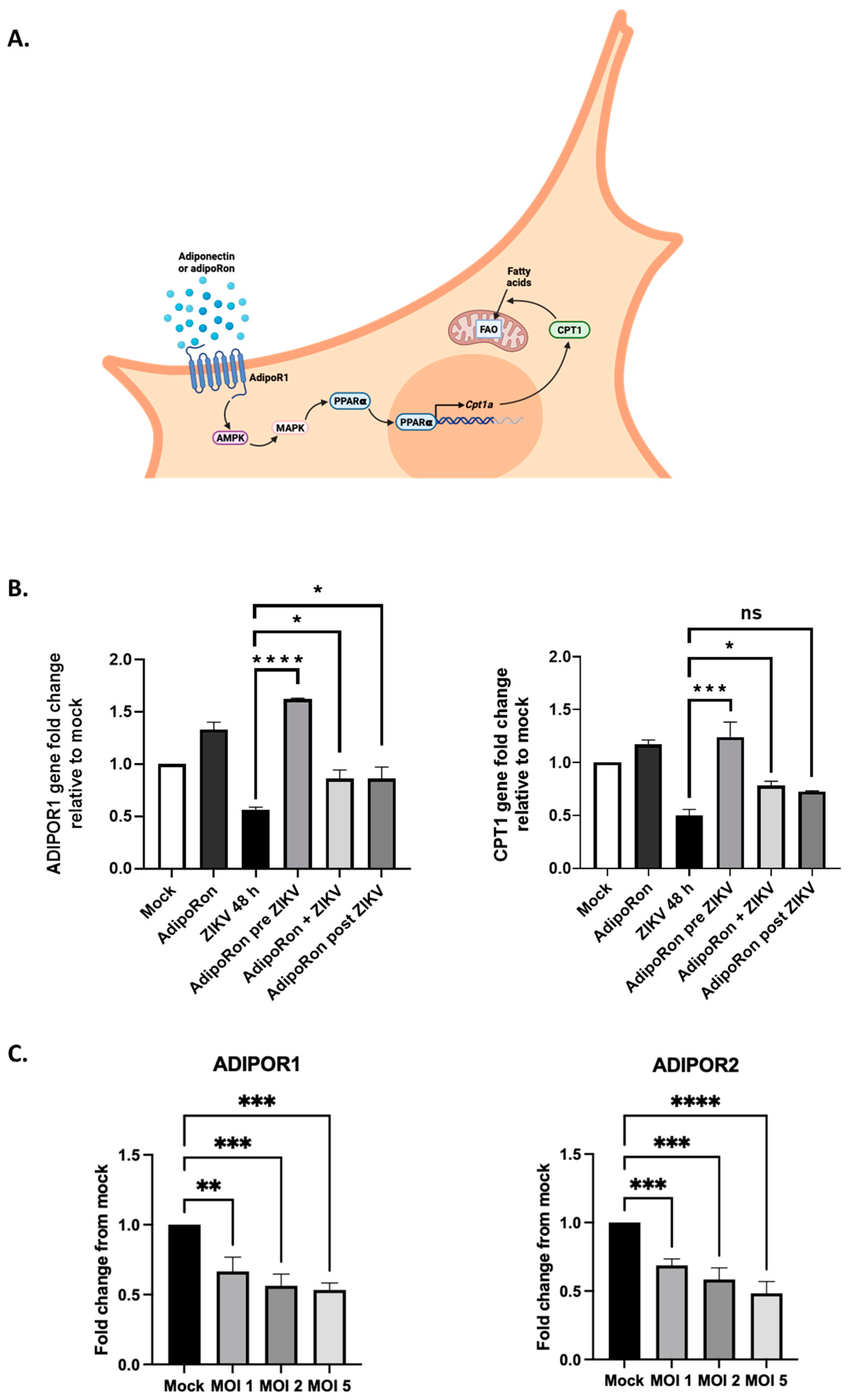
3. Results
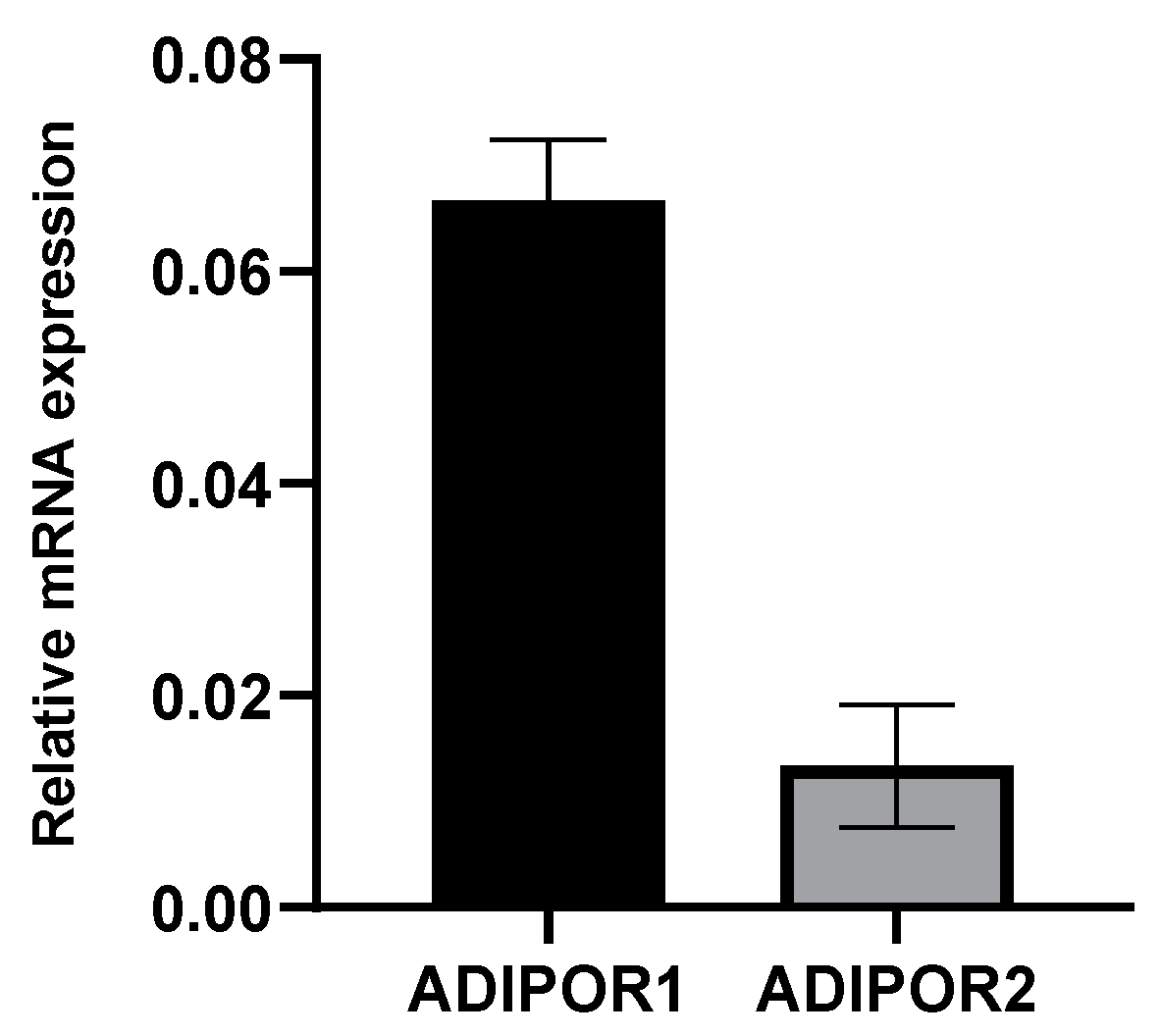
3.1. A549 Cells Express Both Adiponectin Receptor Genes, ADIPOR1 and ADIPOR2
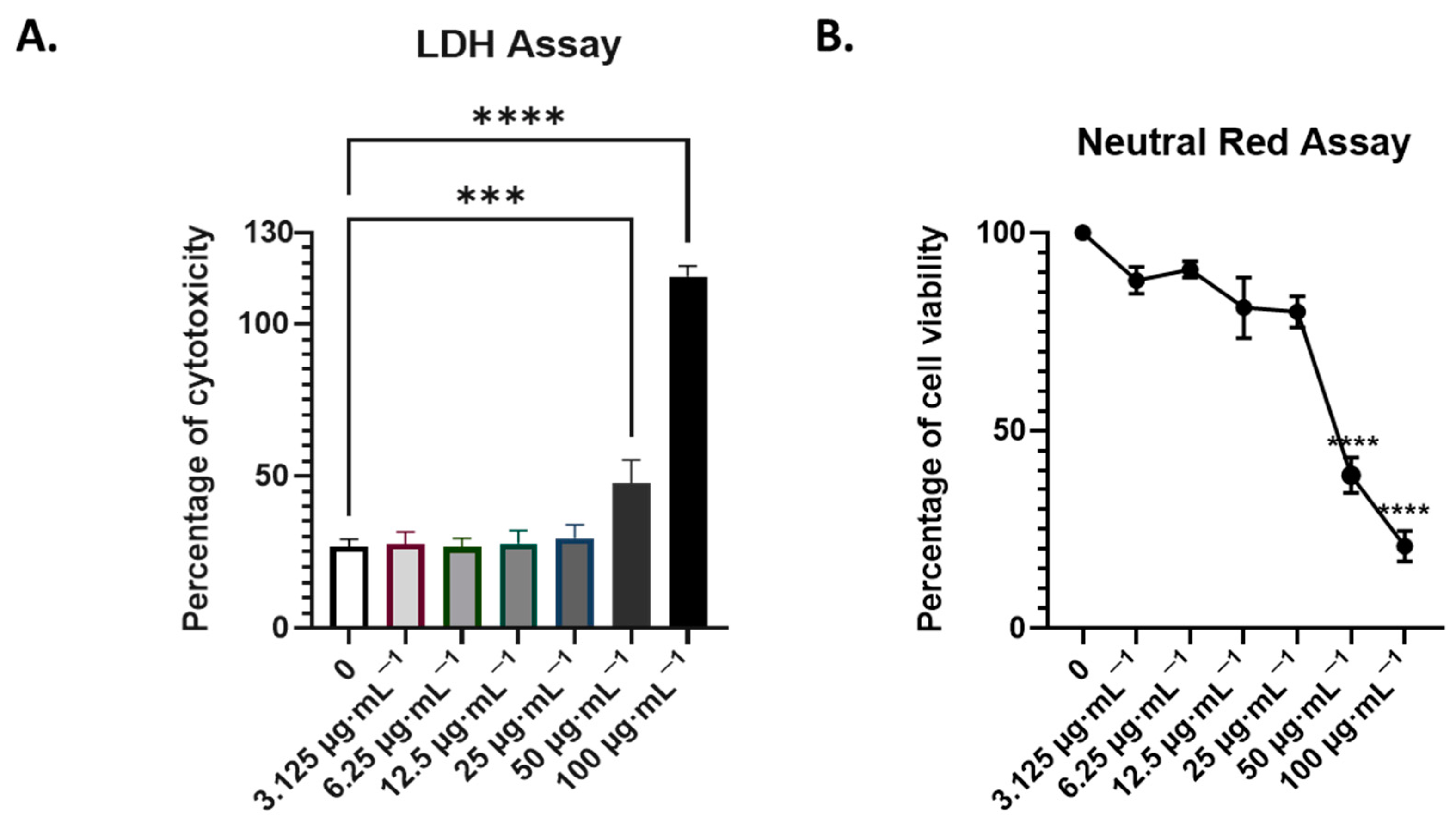
3.2. Assessment of the Non-Toxic Concentration of AdipoRon in A549 Cells
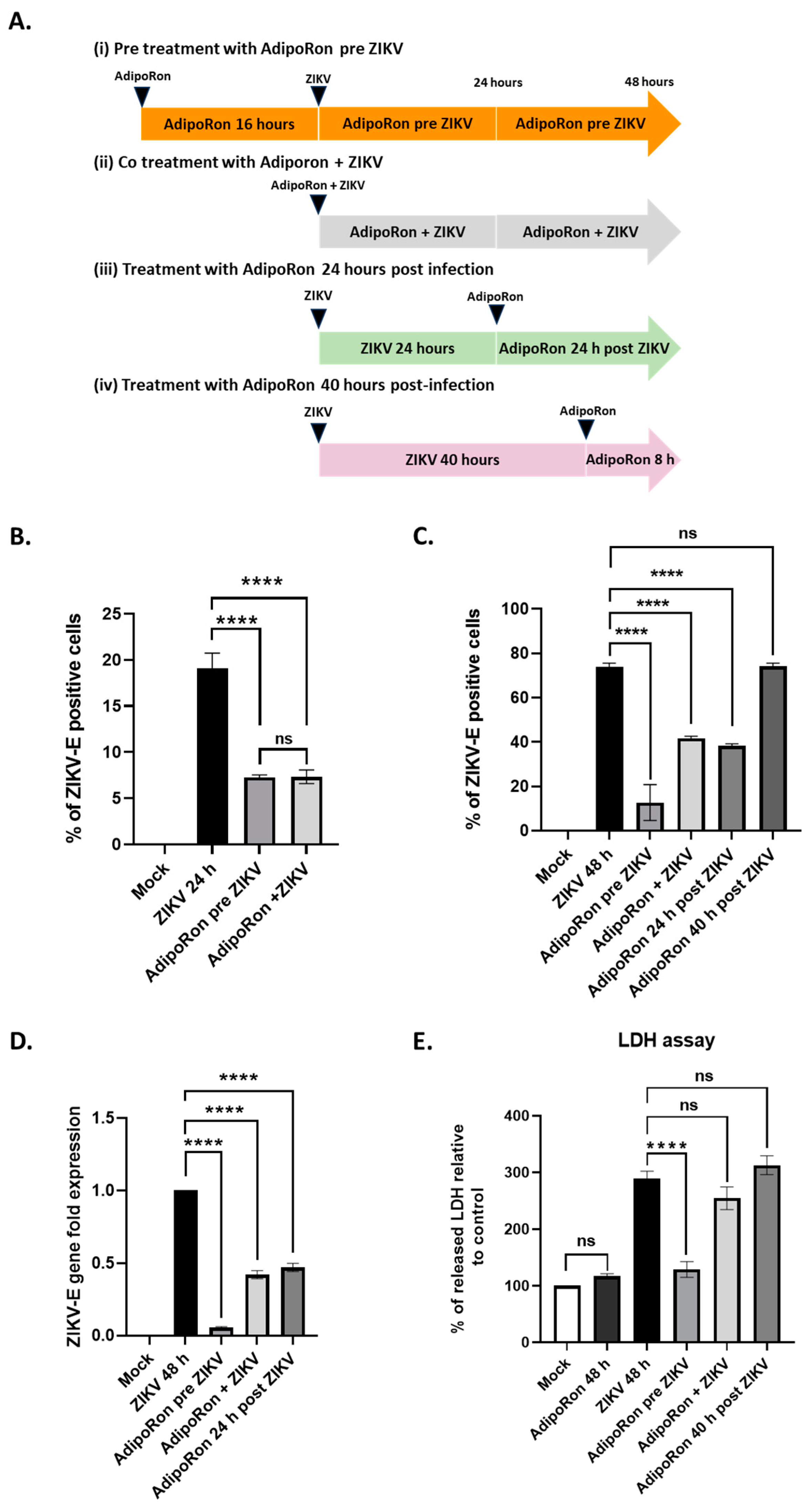
3.3. AdipoRon Displays Antiviral and Cytoprotective Effects
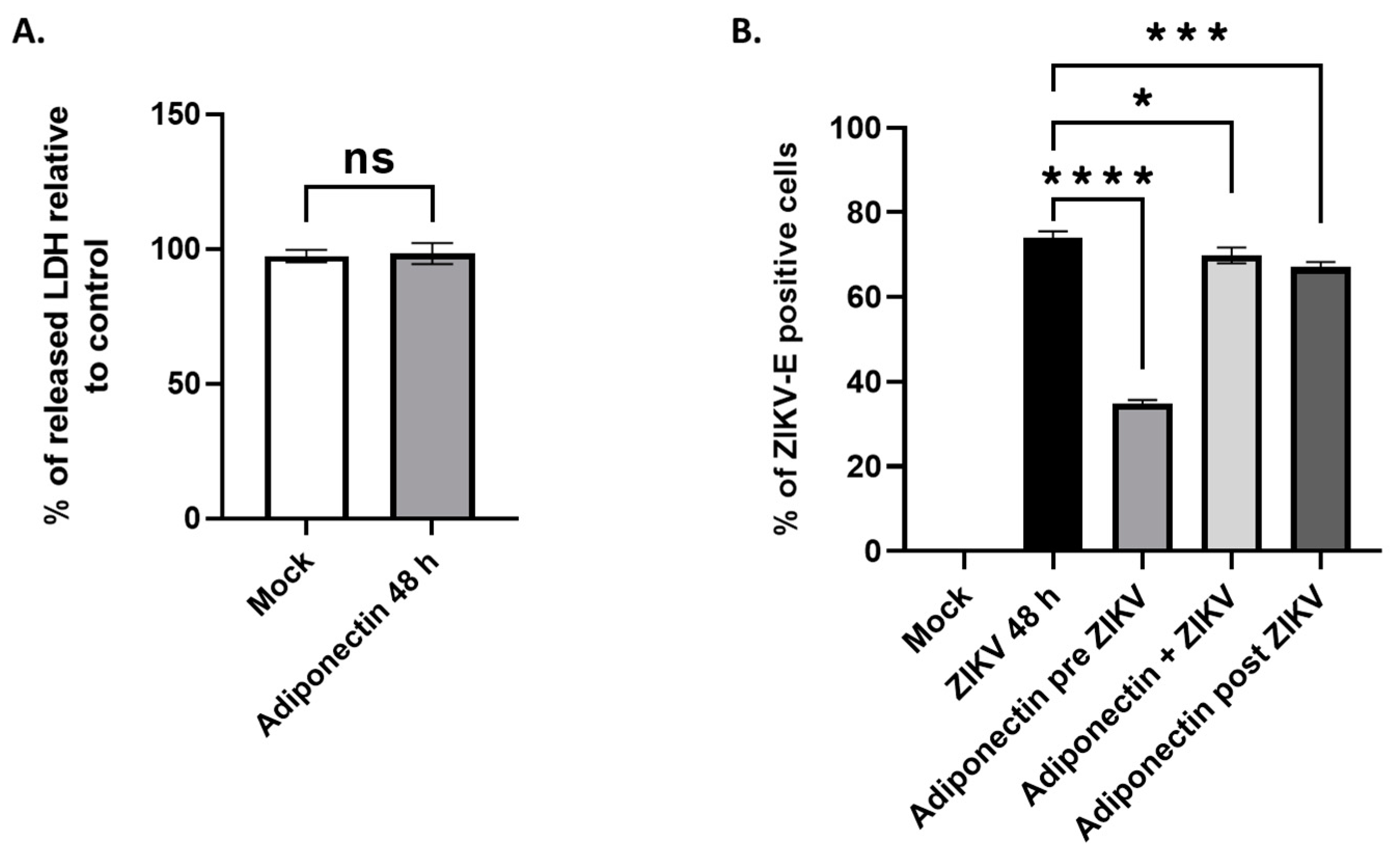
3.4. Adiponectin, like Its Pharmacological Analog AdipoRon, Has Anti-ZIKV Properties
3.5. ZIKV Downregulates ADIPOR1 and CPT1 Gene Expression
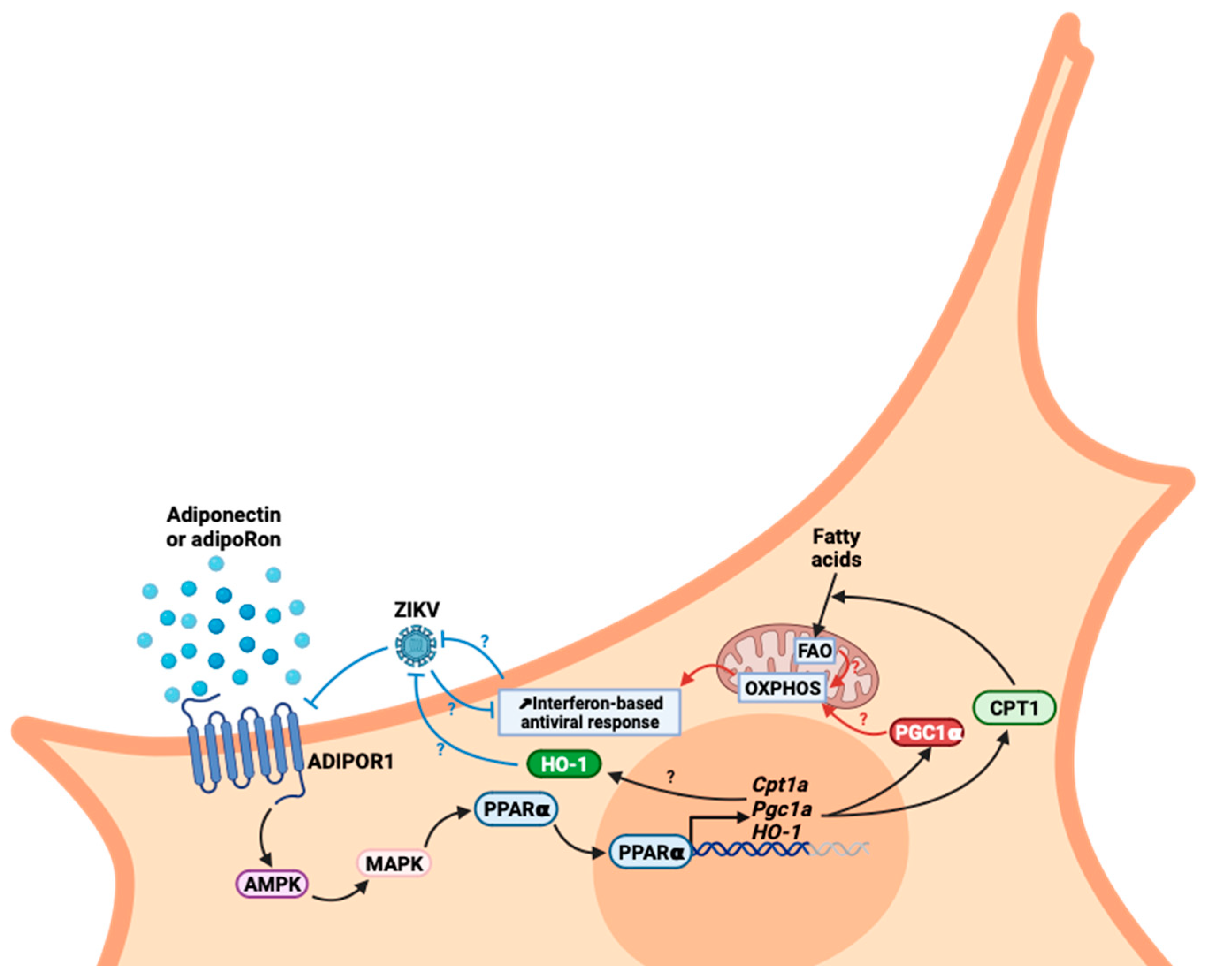
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, B.-H.; Yun, S.-I.; Woolley, M.; Lee, Y.-M. Zika Virus: History, Epidemiology, Transmission, and Clinical Presentation. J. Neuroimmunol. 2017, 308, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Vasilakis, N.; Musso, D. History and Emergence of Zika Virus. J. Infect. Dis. 2017, 216, S860–S867. [Google Scholar] [CrossRef] [PubMed]
- White, M.K.; Wollebo, H.S.; David Beckham, J.; Tyler, K.L.; Khalili, K. Zika Virus: An Emergent Neuropathological Agent. Ann. Neurol. 2016, 80, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.-I.; Lee, Y.-M. Zika Virus: An Emerging Flavivirus. J. Microbiol. 2017, 55, 204–219. [Google Scholar] [CrossRef] [PubMed]
- Beaver, J.T.; Lelutiu, N.; Habib, R.; Skountzou, I. Evolution of Two Major Zika Virus Lineages: Implications for Pathology, Immune Response, and Vaccine Development. Front. Immunol. 2018, 9, 1640. [Google Scholar] [CrossRef] [PubMed]
- Baud, D.; Gubler, D.J.; Schaub, B.; Lanteri, M.C.; Musso, D. An Update on Zika Virus Infection. Lancet 2017, 390, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Marbán-Castro, E.; Goncé, A.; Fumadó, V.; Romero-Acevedo, L.; Bardají, A. Zika Virus Infection in Pregnant Women and Their Children: A Review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 265, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Runge-Ranzinger, S.; Morrison, A.C.; Manrique-Saide, P.; Horstick, O. Zika Transmission Patterns: A Meta-review. Trop. Med. Int. Health 2019, 24, 523–529. [Google Scholar] [CrossRef]
- Wen, Z.; Song, H.; Ming, G. How Does Zika Virus Cause Microcephaly? Genes Dev. 2017, 31, 849–861. [Google Scholar] [CrossRef]
- Christian, K.M.; Song, H.; Ming, G. Pathophysiology and Mechanisms of Zika Virus Infection in the Nervous System. Annu. Rev. Neurosci. 2019, 42, 249–269. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Kadowaki, T. Adiponectin Receptors: A Review of Their Structure, Function and How They Work. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Heiker, J.T.; Kosel, D.; Beck-Sickinger, A.G. Molecular Mechanisms of Signal Transduction via Adiponectin and Adiponectin Receptors. Biol. Chem. 2010, 391, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Tsao, T.-S. Assembly of Adiponectin Oligomers. Rev. Endocr. Metab. Disord. 2014, 15, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; Jiang, L.; Xu, F.; Lyu, J.; Jiang, X.; He, M.; Liu, Y.; Yang, T.; Leak, R.K.; Stetler, R.A.; et al. Adiponectin Ameliorates Hypoperfusive Cognitive Deficits by Boosting a Neuroprotective Microglial Response. Prog. Neurobiol. 2021, 205, 102125. [Google Scholar] [CrossRef]
- Chan, K.-H.; Lam, K.S.-L.; Cheng, O.-Y.; Kwan, J.S.-C.; Ho, P.W.-L.; Cheng, K.K.-Y.; Chung, S.K.; Ho, J.W.-M.; Guo, V.Y.; Xu, A. Adiponectin Is Protective against Oxidative Stress Induced Cytotoxicity in Amyloid-Beta Neurotoxicity. PLoS ONE 2012, 7, e52354. [Google Scholar] [CrossRef]
- Qiu, G.; Wan, R.; Hu, J.; Mattson, M.P.; Spangler, E.; Liu, S.; Yau, S.-Y.; Lee, T.M.C.; Gleichmann, M.; Ingram, D.K.; et al. Adiponectin Protects Rat Hippocampal Neurons against Excitotoxicity. Age 2011, 33, 155–165. [Google Scholar] [CrossRef]
- Bloemer, J.; Pinky, P.D.; Govindarajulu, M.; Hong, H.; Judd, R.; Amin, R.H.; Moore, T.; Dhanasekaran, M.; Reed, M.N.; Suppiramaniam, V. Role of Adiponectin in Central Nervous System Disorders. Neural Plast. 2018, 2018, 4593530. [Google Scholar] [CrossRef]
- Rizzo, M.R.; Fasano, R.; Paolisso, G. Adiponectin and Cognitive Decline. Int. J. Mol. Sci. 2020, 21, 2010. [Google Scholar] [CrossRef]
- Thundyil, J.; Pavlovski, D.; Sobey, C.G.; Arumugam, T.V. Adiponectin Receptor Signalling in the Brain. Br. J. Pharmacol. 2012, 165, 313–327. [Google Scholar] [CrossRef]
- Liu, J.; Guo, M.; Zhang, D.; Cheng, S.-Y.; Liu, M.; Ding, J.; Scherer, P.E.; Liu, F.; Lu, X.-Y. Adiponectin Is Critical in Determining Susceptibility to Depressive Behaviors and Has Antidepressant-like Activity. Proc. Natl. Acad. Sci. USA 2012, 109, 12248–12253. [Google Scholar] [CrossRef] [PubMed]
- Wędrychowicz, A. Peptides from Adipose Tissue in Mental Disorders. WJP 2014, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Huo, T.; Guo, F.; Wang, H.; Wei, H.; Yang, Q.; Dong, H.; Wang, Q.; Xiong, L. Globular Adiponectin Elicits Neuroprotection by Inhibiting NADPH Oxidase-Mediated Oxidative Damage in Ischemic Stroke. Neuroscience 2013, 248, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Zhao, L.; Liu, H.; Guo, H.; Guo, W.; Zheng, L.; Liu, X.; Wu, X.; Luo, J.; Li, X.; et al. Adiponectin Confers Neuroprotection against Cerebral Ischemia-Reperfusion Injury through Activating the cAMP/PKA-CREB-BDNF Signaling. Brain Res. Bull. 2018, 143, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Straub, L.G.; Scherer, P.E. Metabolic Messengers: Adiponectin. Nat. Metab. 2019, 1, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Clain, J.; Couret, D.; Planesse, C.; Krejbich-Trotot, P.; Meilhac, O.; Lefebvre d’Hellencourt, C.; Viranaicken, W.; Diotel, N. Distribution of Adiponectin Receptors in the Brain of Adult Mouse: Effect of a Single Dose of the Adiponectin Receptor Agonist, AdipoRON, on Ischemic Stroke. Brain Sci. 2022, 12, 680. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Kang, S.M.; Kim, E.; Kim, C.-H.; Song, H.-T.; Lee, J.E. Adiponectin Receptor-Mediated Signaling Ameliorates Cerebral Cell Damage and Regulates the Neurogenesis of Neural Stem Cells at High Glucose Concentrations: An In Vivo and In Vitro Study. Cell Death Dis. 2015, 6, e1844. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Daniele, A.; Salzillo, A.; Ragone, A.; Naviglio, S.; Sapio, L. AdipoRon and Other Adiponectin Receptor Agonists as Potential Candidates in Cancer Treatments. Int. J. Mol. Sci. 2021, 22, 5569. [Google Scholar] [CrossRef]
- Okada-Iwabu, M.; Yamauchi, T.; Iwabu, M.; Honma, T.; Hamagami, K.; Matsuda, K.; Yamaguchi, M.; Tanabe, H.; Kimura-Someya, T.; Shirouzu, M.; et al. A Small-Molecule AdipoR Agonist for Type 2 Diabetes and Short Life in Obesity. Nature 2013, 503, 493–499. [Google Scholar] [CrossRef]
- Holland, W.L.; Scherer, P.E. Ronning After the Adiponectin Receptors. Science 2013, 342, 1460–1461. [Google Scholar] [CrossRef]
- Formolo, D.A.; Lee, T.H.; Yu, J.; Lin, K.; Chen, G.; Kranz, G.S.; Yau, S.-Y. Increasing Adiponectin Signaling by Sub-Chronic AdipoRon Treatment Elicits Antidepressant- and Anxiolytic-Like Effects Independent of Changes in Hippocampal Plasticity. Biomedicines 2023, 11, 249. [Google Scholar] [CrossRef]
- Ng, R.C.-L.; Jian, M.; Ma, O.K.-F.; Bunting, M.; Kwan, J.S.-C.; Zhou, G.-J.; Senthilkumar, K.; Iyaswamy, A.; Chan, P.-K.; Li, M.; et al. Chronic Oral Administration of adipoRon Reverses Cognitive Impairments and Ameliorates Neuropathology in an Alzheimer’s Disease Mouse Model. Mol. Psychiatry 2021, 26, 5669–5689. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, S.; Debayle, D.; Béchade, C.; Maroteaux, L.; Gay, A.-S.; Bayer, P.; Heurteaux, C.; Guyon, A.; Chabry, J. Adiporon, an Adiponectin Receptor Agonist Acts as an Antidepressant and Metabolic Regulator in a Mouse Model of Depression. Transl. Psychiatry 2018, 8, 159. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Christie, B.R.; van Praag, H.; Lin, K.; Siu, P.M.-F.; Xu, A.; So, K.-F.; Yau, S. AdipoRon Treatment Induces a Dose-Dependent Response in Adult Hippocampal Neurogenesis. Int. J. Mol. Sci. 2021, 22, 2068. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Luo, J.; Liu, H.; Cui, W.; Guo, K.; Zhao, L.; Bai, H.; Guo, W.; Guo, H.; Feng, D.; et al. Recombinant Adiponectin Peptide Ameliorates Brain Injury Following Intracerebral Hemorrhage by Suppressing Astrocyte-Derived Inflammation via the Inhibition of Drp1-Mediated Mitochondrial Fission. Transl. Stroke Res. 2020, 11, 924–939. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zheng, J.; Lu, J.; Sun, Z.; Wang, Z.; Zhang, J. AdipoRon Protects Against Secondary Brain Injury After Intracerebral Hemorrhage via Alleviating Mitochondrial Dysfunction: Possible Involvement of AdipoR1–AMPK–PGC1α Pathway. Neurochem. Res. 2019, 44, 1678–1689. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, X.; Wang, J.; Shi, Y.; Hu, Q.; Cui, W.; Bai, H.; Zhou, J.; Du, Y.; Han, L.; et al. Adiponectin/AdiopR1 Signaling Prevents Mitochondrial Dysfunction and Oxidative Injury after Traumatic Brain Injury in a SIRT3 Dependent Manner. Redox Biol. 2022, 54, 102390. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Guo, H.; Li, X.; Yue, L.; Liu, H.; Zhao, L.; Bai, H.; Liu, X.; Wu, X.; Qu, Y. Adiponectin Attenuates Oxygen–Glucose Deprivation-Induced Mitochondrial Oxidative Injury and Apoptosis in Hippocampal HT22 Cells via the JAK2/STAT3 Pathway. Cell Transpl. 2018, 27, 1731–1743. [Google Scholar] [CrossRef]
- Okada-Iwabu, M.; Iwabu, M.; Ueki, K.; Yamauchi, T.; Kadowaki, T. Perspective of Small-Molecule AdipoR Agonist for Type 2 Diabetes and Short Life in Obesity. Diabetes Metab. J. 2015, 39, 363. [Google Scholar] [CrossRef]
- Zatorski, H.; Salaga, M.; Zielińska, M.; Majchrzak, K.; Binienda, A.; Kordek, R.; Małecka-Panas, E.; Fichna, J. AdipoRon, an Orally Active, Synthetic Agonist of AdipoR1 and AdipoR2 Receptors Has Gastroprotective Effect in Experimentally Induced Gastric Ulcers in Mice. Molecules 2021, 26, 2946. [Google Scholar] [CrossRef]
- Kim, Y.; Park, C.W. Mechanisms of Adiponectin Action: Implication of Adiponectin Receptor Agonism in Diabetic Kidney Disease. Int. J. Mol. Sci. 2019, 20, 1782. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, M.; Manglani, K.; Upadhyay, P.; Azad, M.; Gupta, S. AdipoRon Induces AMPK Activation and Ameliorates Alzheimer’s like Pathologies and Associated Cognitive Impairment in APP/PS1 Mice. Neurobiol. Dis. 2022, 174, 105876. [Google Scholar] [CrossRef] [PubMed]
- Frumence, E.; Roche, M.; Krejbich-Trotot, P.; El-Kalamouni, C.; Nativel, B.; Rondeau, P.; Missé, D.; Gadea, G.; Viranaicken, W.; Desprès, P. The South Pacific Epidemic Strain of Zika Virus Replicates Efficiently in Human Epithelial A549 Cells Leading to IFN-β Production and Apoptosis Induction. Virology 2016, 493, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Oeyen, M.; Meyen, E.; Doijen, J.; Schols, D. In-Depth Characterization of Zika Virus Inhibitors Using Cell-Based Electrical Impedance. Microbiol. Spectr. 2022, 10, e00491-22. [Google Scholar] [CrossRef] [PubMed]
- Morales Vasquez, D.; Park, J.-G.; Ávila-Pérez, G.; Nogales, A.; De La Torre, J.C.; Almazan, F.; Martinez-Sobrido, L. Identification of Inhibitors of ZIKV Replication. Viruses 2020, 12, 1041. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.S.; Santos, F.C.P.; Campana, P.R.V.; Costa, V.V.; de Pádua, R.M.; Souza, D.G.; Teixeira, M.M.; Braga, F.C. Natural Products and Derivatives as Potential Zika Virus Inhibitors: A Comprehensive Review. Viruses 2023, 15, 1211. [Google Scholar] [CrossRef] [PubMed]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of Zika Virus Infection in Human Skin Cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef]
- Repetto, G.; Del Peso, A.; Zurita, J.L. Neutral Red Uptake Assay for the Estimation of Cell Viability/Cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Turpin, J.; Frumence, E.; Harrabi, W.; Haddad, J.G.; El Kalamouni, C.; Desprès, P.; Krejbich-Trotot, P.; Viranaïcken, W. Zika Virus Subversion of Chaperone GRP78/BiP Expression in A549 Cells during UPR Activation. Biochimie 2020, 175, 99–105. [Google Scholar] [CrossRef]
- Gobillot, T.A.; Humes, D.; Sharma, A.; Kikawa, C.; Overbaugh, J. The Robust Restriction of Zika Virus by Type-I Interferon in A549 Cells Varies by Viral Lineage and Is Not Determined by IFITM3. Viruses 2020, 12, 503. [Google Scholar] [CrossRef]
- Turpin, J.; Frumence, E.; Desprès, P.; Viranaicken, W.; Krejbich-Trotot, P. The ZIKA Virus Delays Cell Death Through the Anti-Apoptotic Bcl-2 Family Proteins. Cells 2019, 8, 1338. [Google Scholar] [CrossRef] [PubMed]
- Turpin, J.; El-Safadi, D.; Lebeau, G.; Frumence, E.; Desprès, P.; Viranaïcken, W.; Krejbich-Trotot, P. CHOP Pro-Apoptotic Transcriptional Program in Response to ER Stress Is Hacked by Zika Virus. Int. J. Mol. Sci. 2021, 22, 3750. [Google Scholar] [CrossRef] [PubMed]
- Vicenti, I.; Boccuto, A.; Giannini, A.; Dragoni, F.; Saladini, F.; Zazzi, M. Comparative Analysis of Different Cell Systems for Zika Virus (ZIKV) Propagation and Evaluation of Anti-ZIKV Compounds in Vitro. Virus Res. 2018, 244, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Cho, J.Y.; Pham, A.; Ramsdell, J.; Broide, D.H. Adiponectin and Functional Adiponectin Receptor 1 Are Expressed by Airway Epithelial Cells in Chronic Obstructive Pulmonary Disease. J. Immunol. 2009, 182, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Ito, Y.; Tsuchida, A.; Yokomizo, T.; Kita, S.; Sugiyama, T.; Miyagishi, M.; Hara, K.; Tsunoda, M.; et al. Cloning of Adiponectin Receptors That Mediate Antidiabetic Metabolic Effects. Nature 2003, 423, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-M.; Yang, C.-M.; Chang, J.-F.; Wu, C.-S.; Sia, K.-C.; Lin, W.-N. AdipoR-Increased Intracellular ROS Promotes cPLA2 and COX-2 Expressions via Activation of PKC and P300 in Adiponectin-Stimulated Human Alveolar Type II Cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L255–L269. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.J.; Lee, G.Y.; Chung, J.-J.; Ahn, Y.H.; Hong, S.H.; Kim, J.B. Adiponectin Increases Fatty Acid Oxidation in Skeletal Muscle Cells by Sequential Activation of AMP-Activated Protein Kinase, P38 Mitogen-Activated Protein Kinase, and Peroxisome Proliferator–Activated Receptor α. Diabetes 2006, 55, 2562–2570. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, T.V.B.; de Alencar Ximenes, R.A.; Miranda-Filho, D.d.B.; Souza, W.V.; Montarroyos, U.R.; de Melo, A.P.L.; Valongueiro, S.; de Albuquerque, M. de F.P.M.; Braga, C.; Filho, S.P.B.; et al. Association between Microcephaly, Zika Virus Infection, and Other Risk Factors in Brazil: Final Report of a Case-Control Study. Lancet Infect. Dis. 2018, 18, 328–336. [Google Scholar] [CrossRef]
- Gilbert, R.K.; Petersen, L.R.; Honein, M.A.; Moore, C.A.; Rasmussen, S.A. Zika Virus as a Cause of Birth Defects: Were the Teratogenic Effects of Zika Virus Missed for Decades? Birth Defects Res. 2023, 115, 265–274. [Google Scholar] [CrossRef]
- Zorrilla, C.D.; García García, I.; García Fragoso, L.; De La Vega, A. Zika Virus Infection in Pregnancy: Maternal, Fetal, and Neonatal Considerations. J. Infect. Dis. 2017, 216, S891–S896. [Google Scholar] [CrossRef]
- Watts, J.L.; Ralston, A. The Fetal Lineage Is Susceptible to Zika Virus Infection within Days of Fertilization. Development 2022, 149, dev200501. [Google Scholar] [CrossRef] [PubMed]
- Bonifay, T.; Le Turnier, P.; Epelboin, Y.; Carvalho, L.; De Thoisy, B.; Djossou, F.; Duchemin, J.-B.; Dussart, P.; Enfissi, A.; Lavergne, A.; et al. Review on Main Arboviruses Circulating on French Guiana, An Ultra-Peripheric European Region in South America. Viruses 2023, 15, 1268. [Google Scholar] [CrossRef] [PubMed]
- Noisumdaeng, P.; Dangsagul, W.; Sangsiriwut, K.; Prasertsopon, J.; Changsom, D.; Yoksan, S.; Ajawatanawong, P.; Buathong, R.; Puthavathana, P. Molecular Characterization and Geographical Distribution of Zika Virus Worldwide from 1947 to 2022. Int. J. Infect. Dis. 2023, 136, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Marquine, S.; Durand, G.A.; Modenesi, G.; Khouadhria, S.; Piorkowski, G.; Badaut, C.; Canivez, T.; De Lamballerie, X.; Grard, G.; Klitting, R. Sequence Data from a Travel-Associated Case of Microcephaly Highlight a Persisting Risk Due to Zika Virus Circulation in Thailand. J. Infect. Dis. 2023, jiad322. [Google Scholar] [CrossRef] [PubMed]
- Braga, C.; Martelli, C.M.T.; Souza, W.V.; Luna, C.F.; Albuquerque, M.D.F.P.M.; Mariz, C.A.; Morais, C.N.L.; Brito, C.A.A.; Melo, C.F.C.A.; Lins, R.D.; et al. Seroprevalence of Dengue, Chikungunya and Zika at the Epicenter of the Congenital Microcephaly Epidemic in Northeast Brazil: A Population-Based Survey. PLoS Negl. Trop. Dis. 2023, 17, e0011270. [Google Scholar] [CrossRef] [PubMed]
- Aubry, F.; Jacobs, S.; Darmuzey, M.; Lequime, S.; Delang, L.; Fontaine, A.; Jupatanakul, N.; Miot, E.F.; Dabo, S.; Manet, C.; et al. Recent African Strains of Zika Virus Display Higher Transmissibility and Fetal Pathogenicity than Asian Strains. Nat. Commun. 2021, 12, 916. [Google Scholar] [CrossRef]
- Salampe, M.; Mamada, S.S.; Evary, Y.M.; Mitra, S.; Bin Emran, T.; Harapan, H.; Nainu, F.; Simal-Gandara, J. Promising Marine Natural Products for Tackling Viral Outbreaks:A Focus on Possible Targets and Structure-Activity Relationship. Curr. Top. Med. Chem. 2023, 23, 1352–1379. [Google Scholar] [CrossRef]
- Bernatchez, J.A.; Tran, L.T.; Li, J.; Luan, Y.; Siqueira-Neto, J.L.; Li, R. Drugs for the Treatment of Zika Virus Infection. J. Med. Chem. 2020, 63, 470–489. [Google Scholar] [CrossRef]
- Han, Y.; Mesplède, T. Investigational Drugs for the Treatment of Zika Virus Infection: A Preclinical and Clinical Update. Expert Opin. Investig. Drugs 2018, 27, 951–962. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, D.; Jose, J.; Giri, R.; Mysorekar, I.U. Drugs to Limit Zika Virus Infection and Implication for Maternal-Fetal Health. Front. Virol. 2022, 2, 928599. [Google Scholar] [CrossRef]
- Martinello, M.; Naggie, S.; Rockstroh, J.K.; Matthews, G.V. Direct-Acting Antiviral Therapy for Treatment of Acute and Recent Hepatitis C Virus Infection: A Narrative Review. Clin. Infect. Dis. 2023, 77, S238–S244. [Google Scholar] [CrossRef] [PubMed]
- Mayberry, J.; Lee, W.M. The Revolution in Treatment of Hepatitis C. Med. Clin. N. Am. 2019, 103, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Sun, Z.; Liang, F.; Xu, W.; Lu, J.; Shi, L.; Shao, A.; Yu, J.; Zhang, J. AdipoRon Attenuates Neuroinflammation After Intracerebral Hemorrhage Through AdipoR1-AMPK Pathway. Neuroscience 2019, 412, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, A.; Yamauchi, T.; Ito, Y.; Hada, Y.; Maki, T.; Takekawa, S.; Kamon, J.; Kobayashi, M.; Suzuki, R.; Hara, K.; et al. Insulin/Foxo1 Pathway Regulates Expression Levels of Adiponectin Receptors and Adiponectin Sensitivity. J. Biol. Chem. 2004, 279, 30817–30822. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.K.; Dang, J.; Qin, Y.; Lichinchi, G.; Bansal, V.; Rana, T.M. Zika Virus Infection Reprograms Global Transcription of Host Cells to Allow Sustained Infection. Emerg. Microbes Infect. 2017, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Kinney, B.; Yoo, H.S.; Lee, B.; Schaack, J.; Shao, J. Adiponectin Increases Skeletal Muscle Mitochondrial Biogenesis by Suppressing Mitogen-Activated Protein Kinase Phosphatase-1. Diabetes 2012, 61, 1463–1470. [Google Scholar] [CrossRef]
- Majeed, Y.; Halabi, N.; Madani, A.Y.; Engelke, R.; Bhagwat, A.M.; Abdesselem, H.; Agha, M.V.; Vakayil, M.; Courjaret, R.; Goswami, N.; et al. SIRT1 Promotes Lipid Metabolism and Mitochondrial Biogenesis in Adipocytes and Coordinates Adipogenesis by Targeting Key Enzymatic Pathways. Sci. Rep. 2021, 11, 8177. [Google Scholar] [CrossRef]
- Pal, S.; Singh, M.; Porwal, K.; Rajak, S.; Das, N.; Rajput, S.; Trivedi, A.K.; Maurya, R.; Sinha, R.A.; Siddiqi, M.I.; et al. Adiponectin Receptors by Increasing Mitochondrial Biogenesis and Respiration Promote Osteoblast Differentiation: Discovery of Isovitexin as a New Class of Small Molecule Adiponectin Receptor Modulator with Potential Osteoanabolic Function. Eur. J. Pharmacol. 2021, 913, 174634. [Google Scholar] [CrossRef]
- Thaker, S.K.; Ch’ng, J.; Christofk, H.R. Viral Hijacking of Cellular Metabolism. BMC Biol. 2019, 17, 59. [Google Scholar] [CrossRef]
- Lebeau, G.; Paulo-Ramos, A.; Hoarau, M.; El Safadi, D.; Meilhac, O.; Krejbich-Trotot, P.; Roche, M.; Viranaicken, W. Metabolic Dependency Shapes Bivalent Antiviral Response in Host Cells: The Role of Glutamine. bioRxiv 2023. [Google Scholar] [CrossRef]
- Lebeau, G.; El Safadi, D.; Paulo-Ramos, A.; Hoareau, M.; Desprès, P.; Krejbich-Trotot, P.; Chouchou, F.; Roche, M.; Viranaicken, W. The Efficient Antiviral Response of A549 Cells Is Enhanced When Mitochondrial Respiration Is Promoted. Pathogens 2022, 11, 1168. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gouilly, J.; Ferrat, Y.J.; Espino, A.; Glaziou, Q.; Cartron, G.; El Costa, H.; Al-Daccak, R.; Jabrane-Ferrat, N. Metabolic Reprogramming by Zika Virus Provokes Inflammation in Human Placenta. Nat. Commun. 2020, 11, 2967. [Google Scholar] [CrossRef] [PubMed]
- Gilbert-Jaramillo, J.; Purnama, U.; Molnár, Z.; James, W.S. Zika Virus-Induces Metabolic Alterations in Fetal Neuronal Progenitors That Could Influence in Neurodevelopment during Early Pregnancy. Biol. Open 2023, 12, bio059889. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, K.A.; Sanchez, E.L.; Camarda, R.; Lagunoff, M. Dengue Virus Induces and Requires Glycolysis for Optimal Replication. J. Virol. 2015, 89, 2358–2366. [Google Scholar] [CrossRef] [PubMed]
- Jordan, T.X.; Randall, G. Flavivirus Modulation of Cellular Metabolism. Curr. Opin. Virol. 2016, 19, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Mingo-Casas, P.; Blázquez, A.-B.; Gómez de Cedrón, M.; San-Félix, A.; Molina, S.; Escribano-Romero, E.; Calvo-Pinilla, E.; Jiménez de Oya, N.; Ramírez de Molina, A.; Saiz, J.-C.; et al. Glycolytic Shift during West Nile Virus Infection Provides New Therapeutic Opportunities. J. Neuroinflamm. 2023, 20, 217. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Yu, C.-H.; Jen, C.-Y.; Cheng, C.-F.; Chou, Y.; Chang, C.-C.; Juan, S.-H. Adiponectin-Mediated Heme Oxygenase-1 Induction Protects Against Iron-Induced Liver Injury via a PPARα-Dependent Mechanism. Am. J. Pathol. 2010, 177, 1697–1709. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.A.; León, M.A.; Céspedes, P.F.; Gómez, R.S.; Canedo-Marroquín, G.; Riquelme, S.A.; Salazar-Echegarai, F.J.; Blancou, P.; Simon, T.; Anegon, I.; et al. Heme Oxygenase-1 Modulates Human Respiratory Syncytial Virus Replication and Lung Pathogenesis during Infection. J. Immunol. 2017, 199, 212–223. [Google Scholar] [CrossRef]
- El Kalamouni, C.; Frumence, E.; Bos, S.; Turpin, J.; Nativel, B.; Harrabi, W.; Wilkinson, D.; Meilhac, O.; Gadea, G.; Desprès, P.; et al. Subversion of the Heme Oxygenase-1 Antiviral Activity by Zika Virus. Viruses 2018, 11, 2. [Google Scholar] [CrossRef]
- Barroso, E.; Eyre, E.; Palomer, X.; Vázquez-Carrera, M. The Peroxisome Proliferator-Activated Receptor β/δ (PPARβ/δ) Agonist GW501516 Prevents TNF-α-Induced NF-κB Activation in Human HaCaT Cells by Reducing P65 Acetylation through AMPK and SIRT1. Biochem. Pharmacol. 2011, 81, 534–543. [Google Scholar] [CrossRef]
- Salminen, A.; Hyttinen, J.M.T.; Kaarniranta, K. AMP-Activated Protein Kinase Inhibits NF-κB Signaling and Inflammation: Impact on Healthspan and Lifespan. J. Mol. Med. 2011, 89, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Antonia, R.J.; Baldwin, A.S. IKK Promotes Cytokine-Induced and Cancer-Associated AMPK Activity and Attenuates Phenformin-Induced Cell Death in LKB1-Deficient Cells. Sci. Signal. 2018, 11, eaan5850. [Google Scholar] [CrossRef] [PubMed]
- Jiménez De Oya, N.; Blázquez, A.-B.; Casas, J.; Saiz, J.-C.; Martín-Acebes, M.A. Direct Activation of Adenosine Monophosphate-Activated Protein Kinase (AMPK) by PF-06409577 Inhibits Flavivirus Infection through Modification of Host Cell Lipid Metabolism. Antimicrob. Agents Chemother. 2018, 62, e00360-18. [Google Scholar] [CrossRef] [PubMed]
- Aktepe, T.E.; Pham, H.; Mackenzie, J.M. Differential Utilisation of Ceramide during Replication of the Flaviviruses West Nile and Dengue Virus. Virology 2015, 484, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Stijn, C.M.W.; Kim, J.; Lusis, A.J.; Barish, G.D.; Tangirala, R.K. Macrophage Polarization Phenotype Regulates Adiponectin Receptor Expression and Adiponectin Anti-inflammatory Response. FASEB J. 2015, 29, 636–649. [Google Scholar] [CrossRef] [PubMed]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yin, Z.; Qiu, T.; Chen, J.; Liu, J.; Zhang, X.; Xu, J. Revealing the Characteristics of ZIKV Infection through Tissue-Specific Transcriptome Sequencing Analysis. BMC Genom. 2022, 23, 697. [Google Scholar] [CrossRef] [PubMed]
- Barbalho, S.M.; Méndez-Sánchez, N.; Fornari Laurindo, L. AdipoRon and ADP355, Adiponectin Receptor Agonists, in Metabolic-Associated Fatty Liver Disease (MAFLD) and Nonalcoholic Steatohepatitis (NASH): A Systematic Review. Biochem. Pharmacol. 2023, 218, 115871. [Google Scholar] [CrossRef]
- Moyce Gruber, B.L.; Dolinsky, V.W. The Role of Adiponectin during Pregnancy and Gestational Diabetes. Life 2023, 13, 301. [Google Scholar] [CrossRef]
- Jones, H.N.; Jansson, T.; Powell, T.L. Full-Length Adiponectin Attenuates Insulin Signaling and Inhibits Insulin-Stimulated Amino Acid Transport in Human Primary Trophoblast Cells. Diabetes 2010, 59, 1161–1170. [Google Scholar] [CrossRef]
- Qiao, L.; Yoo, H.S.; Madon, A.; Kinney, B.; Hay, W.W.; Shao, J. Adiponectin Enhances Mouse Fetal Fat Deposition. Diabetes 2012, 61, 3199–3207. [Google Scholar] [CrossRef] [PubMed]
- Gázquez, A.; Rodríguez, F.; Sánchez-Campillo, M.; Martínez-Gascón, L.E.; Arnao, M.B.; Saura-Garre, P.; Albaladejo-Otón, M.D.; Larqué, E. Adiponectin Agonist Treatment in Diabetic Pregnant Rats. J. Endocrinol. 2021, 251, 1–13. [Google Scholar] [CrossRef]
- Qian, X.; Nguyen, H.N.; Jacob, F.; Song, H.; Ming, G. Using Brain Organoids to Understand Zika Virus-Induced Microcephaly. Development 2017, 144, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Boreland, A.J.; Li, X.; Erickson, C.; Jin, M.; Atkins, C.; Pang, Z.P.; Daniels, B.P.; Jiang, P. Developing Human Pluripotent Stem Cell-Based Cerebral Organoids with a Controllable Microglia Ratio for Modeling Brain Development and Pathology. Stem Cell Rep. 2021, 16, 1923–1937. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Buth, J.E.; Vishlaghi, N.; De La Torre-Ubieta, L.; Taxidis, J.; Khakh, B.S.; Coppola, G.; Pearson, C.A.; Yamauchi, K.; Gong, D.; et al. Self-Organized Cerebral Organoids with Human-Specific Features Predict Effective Drugs to Combat Zika Virus Infection. Cell Rep. 2017, 21, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.H.; Kim, K.; Shresta, S. Mouse Models of Zika Virus Transplacental Transmission. Antivir. Res. 2023, 210, 105500. [Google Scholar] [CrossRef]
- Lewandowski, D.; Foik, A.T.; Smidak, R.; Choi, E.H.; Zhang, J.; Hoang, T.; Tworak, A.; Suh, S.; Leinonen, H.; Dong, Z.; et al. Inhibition of Ceramide Accumulation in AdipoR1–/– Mice Increases Photoreceptor Survival and Improves Vision. JCI Insight 2022, 7, e156301. [Google Scholar] [CrossRef]
- Bjursell, M.; Ahnmark, A.; Bohlooly-Y, M.; William-Olsson, L.; Rhedin, M.; Peng, X.-R.; Ploj, K.; Gerdin, A.-K.; Arnerup, G.; Elmgren, A.; et al. Opposing Effects of Adiponectin Receptors 1 and 2 on Energy Metabolism. Diabetes 2007, 56, 583–593. [Google Scholar] [CrossRef]
- Muraleedharan, R.; Gawali, M.V.; Tiwari, D.; Sukumaran, A.; Oatman, N.; Anderson, J.; Nardini, D.; Bhuiyan, M.A.N.; Tkáč, I.; Ward, A.L.; et al. AMPK-Regulated Astrocytic Lactate Shuttle Plays a Non-Cell-Autonomous Role in Neuronal Survival. Cell Rep. 2020, 32, 108092. [Google Scholar] [CrossRef]
- Morant-Ferrando, B.; Jimenez-Blasco, D.; Alonso-Batan, P.; Agulla, J.; Lapresa, R.; Garcia-Rodriguez, D.; Yunta-Sanchez, S.; Lopez-Fabuel, I.; Fernandez, E.; Carmeliet, P.; et al. Fatty Acid Oxidation Organizes Mitochondrial Supercomplexes to Sustain Astrocytic ROS and Cognition. Nat. Metab. 2023, 5, 1290–1302. [Google Scholar] [CrossRef]
- El Safadi, D.; Paulo-Ramos, A.; Hoareau, M.; Roche, M.; Krejbich-Trotot, P.; Viranaïcken, W.; Lebeau, G. The Influence of Metabolism on Immune Response: A Journey to Understand Immunometabolism in the Context of Viral Infection. Viruses 2023, 15, 2399. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | 5′-CAAATTCCATGGCACCGTCA-3′ | 5′-GGAGTGGGTGTCGCTGTTGA-3′ |
| RNAPII | 5′-GAGAGCGTTGAGTTCCAGAACC-3′ | 5′-TGGATGTGTGCGTTGCTCAGCA-3′ |
| RPLP0 | 5′-AGATGCAGCAGATCCGCAT-3′ | 5′-GGATGGCCTTGCGCA-3′ |
| ADIPOR1 | 5′-AGCCTGCGGCTTAATTTGAC-3′ | 5′-CAACTAAGAACGGCCATGCA-3′ |
| ADIPOR2 | 5′-GCAGCCAAGTTTTACCGAAG-3′ | 5′-CACCTCAAATGTGGGCTTTT-3′ |
| CPT1a | 5′-GATCCTGGACAATACCTCGGAG-3′ | 5′-CTCCACAGCATCAAGAGACTGC-3′ |
| ZIKV-E | 5′-CTGGTCACCTGGGGAAACTA-3′ | 5′-GAGCCTTCTCAAAGCACACC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Safadi, D.; Lebeau, G.; Turpin, J.; Lefebvre d’Hellencourt, C.; Diotel, N.; Viranaicken, W.; Krejbich-Trotot, P. The Antiviral Potential of AdipoRon, an Adiponectin Receptor Agonist, Reveals the Ability of Zika Virus to Deregulate Adiponectin Receptor Expression. Viruses 2024, 16, 24. https://doi.org/10.3390/v16010024
El Safadi D, Lebeau G, Turpin J, Lefebvre d’Hellencourt C, Diotel N, Viranaicken W, Krejbich-Trotot P. The Antiviral Potential of AdipoRon, an Adiponectin Receptor Agonist, Reveals the Ability of Zika Virus to Deregulate Adiponectin Receptor Expression. Viruses. 2024; 16(1):24. https://doi.org/10.3390/v16010024
Chicago/Turabian StyleEl Safadi, Daed, Grégorie Lebeau, Jonathan Turpin, Christian Lefebvre d’Hellencourt, Nicolas Diotel, Wildriss Viranaicken, and Pascale Krejbich-Trotot. 2024. "The Antiviral Potential of AdipoRon, an Adiponectin Receptor Agonist, Reveals the Ability of Zika Virus to Deregulate Adiponectin Receptor Expression" Viruses 16, no. 1: 24. https://doi.org/10.3390/v16010024
APA StyleEl Safadi, D., Lebeau, G., Turpin, J., Lefebvre d’Hellencourt, C., Diotel, N., Viranaicken, W., & Krejbich-Trotot, P. (2024). The Antiviral Potential of AdipoRon, an Adiponectin Receptor Agonist, Reveals the Ability of Zika Virus to Deregulate Adiponectin Receptor Expression. Viruses, 16(1), 24. https://doi.org/10.3390/v16010024