

Japanese Encephalitis: Risk of Emergence in the United States and the Resulting Impact
{kind=link}
{kind=link}
Abstract
:1. Introduction
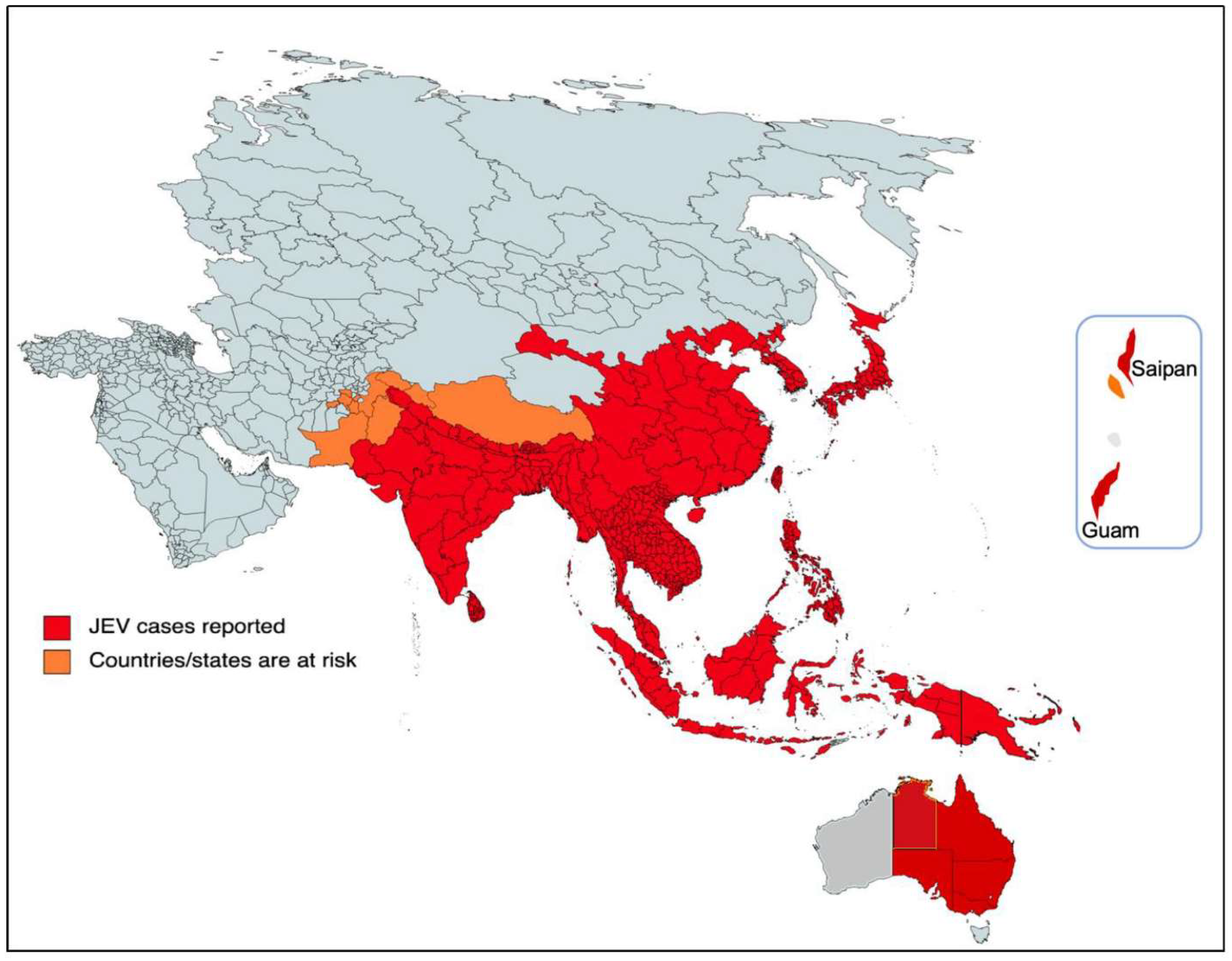
2. Historical Threat
3. The Current Threat
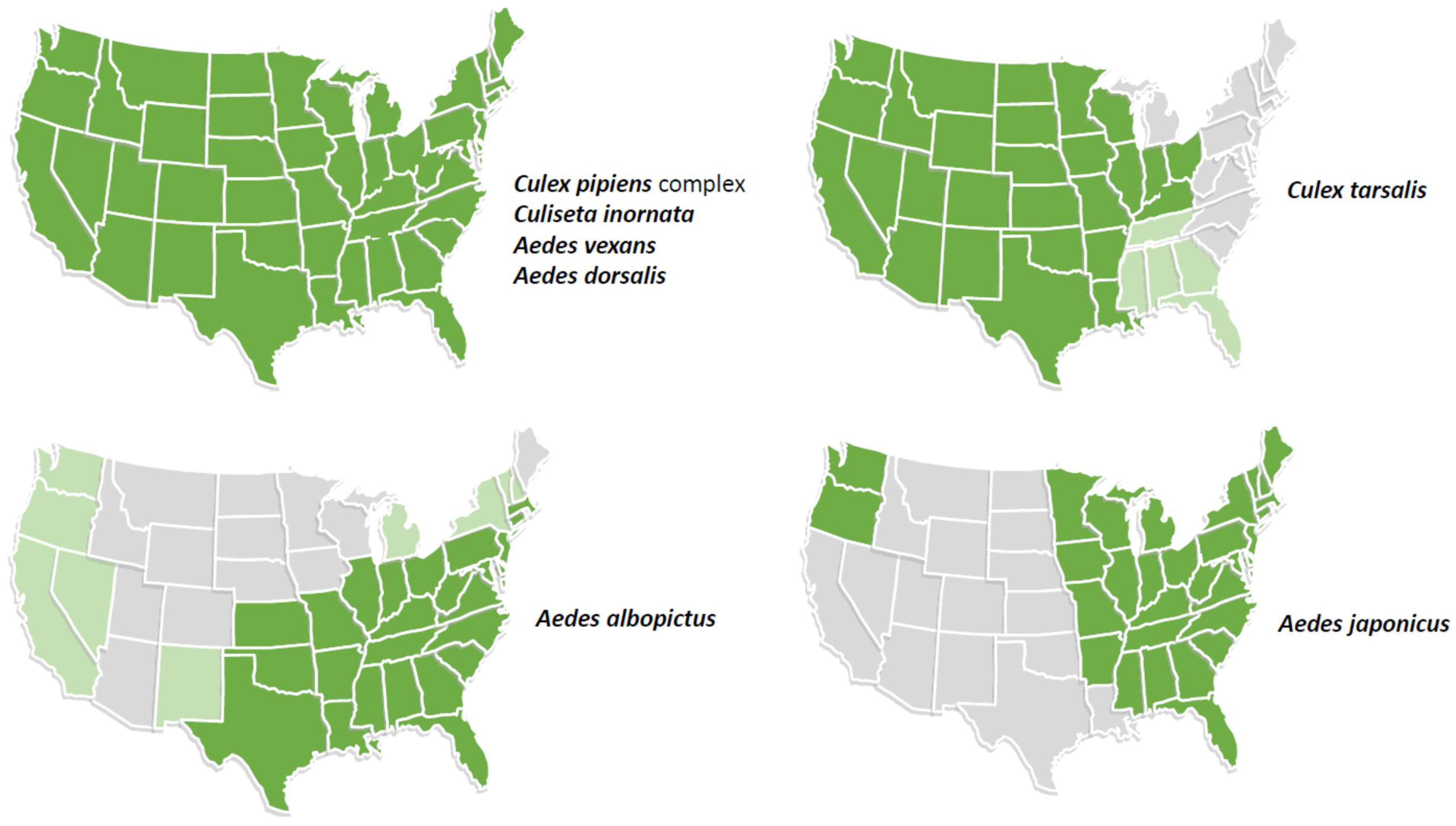
4. Mechanisms of Potential Introduction and Spread of JEV
5. Consequences of and Response to the Introduction of JEV into North America
6. Vaccines as a Countermeasure
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Quan, T.M.; Thao, T.T.N.; Duy, N.M.; Nhat, T.M.; Clapham, H. Estimates of the global burden of Japanese encephalitis and the impact of vac-cination from 2000–2015. eLife 2020, 9, e51027. [Google Scholar] [CrossRef] [PubMed]
- Vannice, K.S.; Hills, S.L.; Schwartz, L.M.; Barrett, A.D.; Heffelfinger, J.; Hombach, J.; Letson, G.W.; Solomon, T.; Marfin, A.A.; Japanese Encephalitis Vaccination Experts Panel. The future of Japanese encephalitis vaccination: Expert recommendations for achieving and maintaining optimal JE control. NPJ Vaccines 2021, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Erlanger, T.E.; Weiss, S.; Keiser, J.; Utzinger, J.; Wiedenmayer, K. Past, present, and future of Japanese encephalitis. Emerg. Infect. Dis. 2009, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, G. Epidemiology of Japanese encephalitis: Past, present, and future prospects. Ther. Clin. Risk Manag. 2015, 11, 435–448. [Google Scholar] [PubMed]
- Auerswald, H.; Maquart, P.O.; Chevalier, V.; Boyer, S. Mosquito Vector Competence for Japanese Encephalitis Virus. Viruses 2021, 13, 1154. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.R.S.; Strathe, E.; Etcheverry, L.; Cohnstaedt, L.W.; McVey, D.S.; Piaggio, J.; Cernicchiaro, N. Assessment of data on vector and host competence for Japanese encephalitis virus: A systematic review of the literature. Prev. Vet. Med. 2018, 154, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.; Bosco-Lauth, A.; Oesterle, P.; Kohler, D.; Bowen, R. North American birds as potential amplifying hosts of Japanese encephali-tis virus. Am. J. Trop. Med. Hyg. 2012, 87, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Rosen, L. The natural history of Japanese encephalitis virus. Annu. Rev. Microbiol. 1986, 40, 395–414. [Google Scholar] [CrossRef]
- Lannes, N.; Summerfield, A.; Filgueira, L. Regulation of inflammation in Japanese encephalitis. J. Neuroinflamm. 2017, 14, 158. [Google Scholar] [CrossRef]
- Myint, K.S.; Gibbons, R.V.; Perng, G.C.; Solomon, T. Unravelling the neuropathogenesis of Japanese encephalitis. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 955–956. [Google Scholar] [CrossRef]
- Chapagain, S.; Pal Singh, P.; Le, K.; Safronetz, D.; Wood, H.; Karniychuk, U. Japanese encephalitis virus persists in the human reproductive epithelium and porcine reproductive tissues. PLoS Negl. Trop. Dis. 2022, 16, e0010656. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.; Shankar, S.K.; Ravi, V.; Chandramuki, A.; Gourie-Devi, M. Japanese encephalitis virus antigen in the human brain and its topo-graphic distribution. Acta Neuropathol. 1995, 89, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P. Japanese encephalitis vaccines: Current vaccines and future prospects. Curr. Top. Microbiol. Immunol. 2002, 267, 105–138. [Google Scholar] [PubMed]
- Hegde, N.R.; Gore, M.M. Japanese encephalitis vaccines: Immunogenicity, protective efficacy, effectiveness, and impact on the burden of disease. Hum. Vaccin. Immunother. 2017, 13, 1320–1337. [Google Scholar] [CrossRef] [PubMed]
- Furuya-Kanamori, L.; Xu, C.; Doi, S.A.R.; Clark, J.; Wangdi, K.; Mills, D.J.; Lau, C.L. Comparison of immunogenicity and safety of licensed Japanese encephalitis vaccines: A systematic review and network meta-analysis. Vaccine 2021, 39, 4429–4436. [Google Scholar] [CrossRef] [PubMed]
- Appaiahgari, M.B.; Vrati, S. Clinical development of IMOJEV ®—A recombinant Japanese encephalitis chimeric vaccine (JE-CV). Expert. Opin. Biol. Ther. 2012, 12, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Jelinek, T. Ixiaro: A new vaccine against Japanese encephalitis. Expert. Rev. Vaccines 2009, 8, 1501–1511. [Google Scholar] [CrossRef] [PubMed]
- Clé, M.; Beck, C.; Salinas, S.; Lecollinet, S.; Gutierrez, S.; Van de Perre, P.; Baldet, T.; Foulongne, V.; Simonin, Y. Usutu virus: A new threat? Epidemiol. Infect. 2019, 147, e232. [Google Scholar] [CrossRef]
- Hanna, J.N.; Ritchie, S.A.; Phillips, D.A.; Lee, J.M.; Hills, S.; van den Hurk, A.F.; Pyke, A.; Johansen, C.A.; Mackenzie, J.S. Japanese encephali-tis in north Queensland, Australia, 1998. Med. J. Aust. 1999, 170, 533–536. [Google Scholar] [CrossRef]
- van den Hurk, A.F.; Skinner, E.; Ritchie, S.A.; Mackenzie, J.S. The Emergence of Japanese Encephalitis Virus in Australia in 2022: Existing Knowledge of Mosquito Vectors. Viruses 2022, 14, 1208. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Williams, D.T.; van den Hurk, A.F.; Smith, D.W.; Currie, B.J. Japanese Encephalitis Virus: The Emergence of Genotype IV in Australia and Its Potential Endemicity. Viruses 2022, 14, 2480. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-J.; Wu, S.-Y. The species of mosquitoes transmitting Japanese B type encephalitis in Fukien. Acta Microbiol. Sin. 1957, 5, 27–32. (In Chinese) [Google Scholar]
- Weng, M.H.; Lien, J.C.; Wang, Y.M.; Lin, C.C.; Lin, H.C.; Chin, C. Isolation of Japanese encephalitis virus from mosquitoes collected in Northern Taiwan between 1995 and 1996. J. Microbiol. Immunol. Infect. 1999, 32, 9–13. [Google Scholar] [PubMed]
- Moore, C.G.; Francy, D.B.; Eliason, D.A.; Monath, T.P. Aedes albopictus in the United States: Rapid spread of a potential disease vector. J. Am. Mosq. Control Assoc. 1988, 4, 356–361. [Google Scholar] [PubMed]
- Rosen, L.; Tesh, R.B.; Lien, J.C.; Cross, J.H. Transovarial transmission of Japanese encephalitis virus by mosquitoes. Science 1978, 199, 909–911. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopic-tus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Nanfack-Minkeu, F.; Delong, A.; Luri, M.; Poelstra, J.W. Invasive Aedes japonicus Mosquitoes Dominate the Aedes Fauna Collected with Gravid Traps in Wooster, Northeastern Ohio, USA. Insects 2023, 14, 56. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.R.S.; Piaggio, J.; Cohnstaedt, L.W.; McVey, D.S.; Cernicchiaro, N. Introduction of the Japanese encephalitis virus (JEV) in the United States—A qualitative risk assessment. Transbound. Emerg. Dis. 2019, 66, 1558–1574. [Google Scholar] [CrossRef]
- Nett, R.J.; Campbell, G.L.; Reisen, W.K. Potential for the emergence of Japanese encephalitis virus in California. Vector Borne Zoonotic Dis. 2009, 9, 511–517. [Google Scholar] [CrossRef]
- Available online: https://www.sacbee.com/news/california/article282384908.html (accessed on 13 October 2023).
- Available online: https://jevisn.org/ (accessed on 15 October 2023).
- Available online: https://www.swinehealth.org/potential-for-jev-in-us-leads-to-usda-statement-on-preparedness-and-testing/ (accessed on 15 October 2023).
- Yakob, L.; Hu, W.; Frentiu, F.D.; Gyawali, N.; Hugo, L.E.; Johnson, B.; Lau, C.; Furuya-Kanamori, L.; Magalhaes, R.S.; Devine, G. Japanese Encephali-tis Emergence in Australia: The Potential Population at Risk. Clin. Infect. Dis. 2023, 76, 335–337. [Google Scholar] [CrossRef]
- Kramer, L.D.; Ciota, A.T.; Kilpatrick, A.M. Introduction, Spread, and Establishment of West Nile Virus in the Americas. J. Med. Entomol. 2019, 56, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.; Brito, A.F.; Swetnam, D.M.; Vogels, C.B.F.; Tokarz, R.E.; Andersen, K.G.; Smith, R.C.; Bedford, T.; Grubaugh, N.D. Twenty years of West Nile virus spread and evolution in the Americas visualized by Nextstrain. PLoS Pathog. 2019, 15, e1008042. [Google Scholar] [CrossRef] [PubMed]
- Ronca, S.E.; Ruff, J.C.; Murray, K.O. A 20-year historical review of West Nile virus since its initial emergence in North America: Has West Nile virus become a neglected tropical disease? PLoS Negl. Trop. Dis. 2021, 15, e0009190. [Google Scholar] [CrossRef]
- Solomon, T.; Dung, N.M.; Kneen, R.; Gainsborough, M.; Vaughn, D.; Khanh, V.T. Japanese encephalitis. J. Neurol. Neurosurg. Psychiatry 2000, 68, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Van den Eynde, C.; Sohier, C.; Matthis, S.; de Regge, N. Japanese encephalitis virus interaction with mosquitoes: A review of vector com-petence, vector capacity and mosquito immunity. Pathogens 2022, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Tolsá-García, M.J.; Wehmeyer, M.L.; Lühken, R.; Roiz, D. Worldwide transmission and infection risk of mosquito vectors of West Nile, St. Louis encephalitis, Usutu and Japanese encephalitis viruses: A systematic review. Sci. Rep. 2023, 13, 308. [Google Scholar] [CrossRef] [PubMed]
- Scherer, W.F.; Buescher, E.L. Ecologic studies of Japanese encephalitis in Japan. I. Introduction. Am. J. Trop. Med. Hyg. 1959, 8, 644–650. [Google Scholar] [CrossRef]
- Lord, J.S.; Gurley, E.S.; Pulliam, J.R. Rethinking Japanese Encephalitis Virus Transmission: A Framework for Implicating Host and Vector Species. PLoS Negl. Trop. Dis. 2015, 9, e0004074. [Google Scholar] [CrossRef]
- Maharaj, P.D.; Bosco-Lauth, A.M.; Langevin, S.A.; Anishchenko, M.; Bowen, R.A.; Reisen, W.K.; Brault, A.C. West Nile and St. Louis encephalitis viral genetic determinants of avian host competence. PLoS Negl. Trop. Dis. 2018, 12, e0006302. [Google Scholar] [CrossRef]
- García-Nicolás, O.; Braun, R.O.; Milona, P.; Lewandowska, M.; Dijkman, R.; Alves, M.P.; Summerfield, A. Targeting of the Nasal Mucosa by Jap-anese Encephalitis Virus for Non-Vector-Borne Transmission. J. Virol. 2018, 92, e01091-18. [Google Scholar] [CrossRef]
- Park, S.L.; Huang, Y.S.; Vanlandingham, D.L. Re-Examining the Importance of Pigs in the Transmission of Japanese Encephalitis Virus. Pathogens 2022, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- Lyons, A.C.; Huang, Y.S.; Park, S.L.; Ayers, V.B.; Hettenbach, S.M.; Higgs, S.; McVey, D.S.; Noronha, L.; Hsu, W.W.; Vanlandingham, D.L. Shed-ding of Japanese Encephalitis Virus in Oral Fluid of Infected Swine. Vector Borne Zoonotic Dis. 2018, 18, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, M.E.; García-Nicolás, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Nougairede, A.; Charrel, R.N.; Posthaus, H.; Oevermann, A.; Summerfield, A. Vector-free transmission and persistence of Japanese encephalitis virus in pigs. Nat. Commun. 2016, 7, 10832. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Palinski, R.; Xu, Y.; Wang, Q.; Cao, S.; Geng, Y.; Zhao, Q.; Wen, Y.; Huang, X.; Yan, Q.; et al. Aerosol and Contact Transmission Following Intranasal Infection of Mice with Japanese Encephalitis Virus. Viruses 2019, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental infection of North American birds with the New York 1999 strain of West Nile virus. Emerg Infect Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.nass.usda.gov/Newsroom/2022/03-30-2022.php (accessed on 18 October 2023).
- Available online: https://nppc.org/the-pork-industry/ (accessed on 16 October 2023).
- Pires, A.F.A.; Peterson, A.; Baron, J.N.; Adams, R.; Martínez-López, B.; Moore, D. Small-scale and backyard livestock owners needs assessment in the western United States. PLoS ONE 2019, 14, e0212372. [Google Scholar] [CrossRef] [PubMed]
- Nidaira, M.; Taira, K.; Itokazu, K.; Kudaka, J.; Nakamura, M.; Ohno, A.; Takasaki, T. Survey of the antibody against Japanese encephalitis virus in Ryukyu wild boars (Sus scrofa riukiuanus) in Okinawa, Japan. Jpn. J. Infect. Dis. 2007, 60, 309. [Google Scholar]
- McClure, M.L.; Burdett, C.L.; Farnsworth, M.L.; Lutman, M.W.; Theobald, D.M.; Riggs, P.D.; Grear, D.A.; Miller, R.S. Modeling and mapping the probability of occurrence of invasive wild pigs across the contiguous United States. PLoS ONE 2015, 10, e0133771. [Google Scholar] [CrossRef]
- Available online: https://www.theguardian.com/environment/2021/nov/24/pig-patrol-amsterdam-airports-innovative-approach-to-flight-safety (accessed on 16 October 2023).
- Platonov, A.; Rossi, G.; Karan, L.; Mironov, K.; Busani, L.; Rezza, G. Does the Japanese encephalitis virus (JEV) represent a threat for human health in Europe? Detection of JEV RNA sequences in birds collected in Italy. Eurosurveillance 2012, 17, 20241. [Google Scholar] [CrossRef]
- Ravanini, P.; Huhtamo, E.; Ilaria, V.; Crobu, M.G.; Nicosia, A.M.; Servino, L.; Rivasi, F.; Allegrini, S.; Miglio, U.; Magri, A.; et al. Japanese encephalitis virus RNA detected in Culex pipiens mosquitoes in Italy. Eurosurveillance 2012, 17, 20221. [Google Scholar] [CrossRef]
- Available online: https://www.farmbiosecurity.com.au/wp-content/uploads/2023/05/IntegratedMosquitoManagementPrinciplesforPiggeries_v3.pdf (accessed on 16 October 2023).
- Work, T.H. On the Japanese B--West Nile virus complex or an arbovirus problem of six continents. Am. J. Trop. Med. Hyg. 1971, 20, 169–186. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.; Mason, G.; Bowen, R. Pathogenesis of Japanese encephalitis virus infection in a golden hamster model and evaluation of flavivirus cross-protective immunity. Am. J. Trop. Med. Hyg. 2011, 84, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Goverdhan, M.K.; Kulkarni, A.B.; Gupta, A.K.; Tupe, C.D.; Rodrigues, J.J. Two-way cross-protection between West Nile and Japanese encepha-litis viruses in bonnet macaques. Acta Virol. 1992, 36, 277–283. [Google Scholar] [PubMed]
- Nemeth, N.M.; Bosco-Lauth, A.M.; Bowen, R.A. Cross-protection between West Nile and Japanese encephalitis viruses in red-winged blackbirds (Agelaius phoeniceus). Avian Dis. 2009, 53, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Lothrop, H.D.; Wheeler, S.S.; Kennsington, M.; Gutierrez, A.; Fang, Y.; Garcia, S.; Lothrop, B. Persistent West Nile virus transmis-sion and the apparent displacement St. Louis encephalitis virus in southeastern California, 2003–2006. J. Med. Entomol. 2008, 45, 494–508. [Google Scholar]
- Bae, W.; Kim, J.H.; Kim, J.; Lee, J.; Hwang, E.S. Changes of epidemiological characteristics of Japanese encephalitis viral infection and birds as a potential viral transmitter in Korea. J. Korean Med. Sci. 2018, 33, e70. [Google Scholar] [CrossRef]
- Diptyanusa, A.; Herini, E.S.; Indarjulianto, S.; Satoto, T.B.T. Estimation of Japanese encephalitis virus infection prevalence in mosquitoes and bats through nationwide sentinel surveillance in Indonesia. PLoS ONE 2022, 17, e0275647. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Guidelines for surveillance, prevention, and control of West Nile virus infection--United States. MMWR Morb. Mortal. Wkly. Rep. 2000, 49, 25–28. [Google Scholar]
- Lindsey, N.P.; Brown, J.A.; Kightlinger, L.; Rosenberg, L.; Fischer, M.; ArboNET Evaluation Working Group. State health department per-ceived utility of and satisfaction with ArboNET, the U.S. National Arboviral Surveillance System. Public. Health Rep. 2012, 127, 383–390. [Google Scholar] [CrossRef]
- Fan, Y.C.; Chen, Y.Y.; Chen, J.M.; Huang, C.; Huang, M.; Chiou, S.S. Effectiveness of Live-Attenuated Genotype III Japanese Encephalitis Viral Vaccine against Circulating Genotype I Viruses in Swine. Viruses 2022, 14, 114. [Google Scholar] [CrossRef]
- Tajima, S.; Taniguchi, S.; Nakayama, E.; Maeki, T.; Inagaki, T.; Saijo, M.; Lim, C.K. Immunogenicity and Protective Ability of Genotype I-Based Recombinant Japanese Encephalitis Virus (JEV) with Attenuation Mutations in E Protein against Genotype V JEV. Vaccines 2021, 9, 1077. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.T.; Chang, L.C.; Lin, S.Y.; Chuang, T.Y.; Ma, C.H.; Inoue, Y.K.; Okuno, T. The effect of vaccination with a live attenuated strain of Japanese encephalitis virus on stillbirths in swine in Taiwan. Bull. World Health Organ. 1972, 46, 465–471. [Google Scholar] [PubMed]
- Nah, J.J.; Yang, D.K.; Kim, H.H.; Song, J.Y. The present and future of veterinary vaccines for Japanese encephalitis in Korea. Clin. Exp. Vaccine Res. 2015, 4, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Fei-fei, G.; Jian, W.; Feng, X.; Li-ping, S.; Quan-yun, S.; Jin-ping, Z.; Pu-yan, C.; Pei-hong, L. Japanese encephalitis protein vaccine candidates expressing neutralizing epitope and M.T hsp70 induce virus-specific memory B cells and long-lasting antibodies in swine. Vaccine 2008, 26, 5590–5594. [Google Scholar] [CrossRef]
- Sheng, Z.; Gao, N.; Cui, X.; Fan, D.; Chen, H.; Wu, N.; Wei, J.; An, J. Electroporation enhances protective immune response of a DNA vaccine against Japanese encephalitis in mice and pigs. Vaccine 2016, 34, 5751–5757. [Google Scholar] [CrossRef]
- García-Nicolás, O.; Ricklin, M.E.; Liniger, M.; Vielle, N.J.; Python, S.; Souque, P.; Charneau, P.; Summerfield, A. A Japanese Encephalitis Virus Vaccine Inducing Antibodies Strongly Enhancing In Vitro Infection Is Protective in Pigs. Viruses 2017, 9, 124. [Google Scholar] [CrossRef]
- Anwar, M.N.; Jiang, C.; Di, D.; Zhang, J.; Guo, S.; Wang, X.; Hameed, M.; Wahaab, A.; Shao, D.; Li, Z.; et al. A Novel Re-combinant Virus-Like Particles Displaying B and T Cell Epitopes of Japanese Encephalitis Virus Offers Protective Immunity in Mice and Guinea Pigs. Vaccines 2021, 9, 980. [Google Scholar] [CrossRef]
- Chen, T.; Zhu, S.; Wei, N.; Zhao, Z.; Niu, J.; Si, Y.; Cao, S.; Ye, J. Protective Immune Responses Induced by an mRNA-LNP Vaccine Encoding prM-E Proteins against Japanese Encephalitis Virus Infection. Viruses 2022, 14, 1121. [Google Scholar] [CrossRef]
- Li, C.; Zhang, L.; Chen, X.; Jiang, D.; Hu, J.; Guo, J.; Ding, J.; Jiao, X.; Bao, W.; Li, Y. Evaluation of the genotype I Japanese encephalitis virus as a stable viral vector for foreign gene expression. Antivir. Res. 2023, 216, 105652. [Google Scholar] [CrossRef]
- Bandrick, M.; Ariza-Nieto, C.; Baidoo, S.K.; Molitor, T.W. Colostral antibody-mediated and cell-mediated immunity contributes to innate and antigen-specific immunity in piglets. Dev. Comp. Immunol. 2014, 43, 114–120. [Google Scholar] [CrossRef]
- Markoff, L. Points to consider in the development of a surrogate for efficacy of novel Japanese encephalitis virus vaccines. Vaccine 2000, 18 (Suppl. S2), 26–32. [Google Scholar] [CrossRef] [PubMed]
- Van Gessel, Y.; Klade, C.S.; Putnak, R.; Formica, A.; Krasaesub, S.; Spruth, M.; Cena, B.; Tungtaeng, A.; Gettayacamin, M.; Dewasthaly, S. Correla-tion of protection against Japanese encephalitis virus and JE vaccine (IXIARO®) induced neutralizing antibody titers. Vaccine 2011, 29, 5925–5931. [Google Scholar] [CrossRef] [PubMed]
- Hombach, J.; Solomon, T.; Kurane, I.; Jacobson, J.; Wood, D. Report on a WHO consultation on immunological endpoints for evaluation of new Japanese encephalitis vaccines, WHO, Geneva 2–3 September 2004. Vaccine 2005, 23, 5205–5211. [Google Scholar] [CrossRef] [PubMed]
- Food & Drug Administration. Summary Basis of Regulatory Action; IXIARO; Food & Drug Administration: Montgomery/Prince Georges, MA, USA, 2018. [Google Scholar]
- Fischer, M.; Lindsey, N.; Staples, J.E.; Hills, S. Japanese encephalitis vaccines: Recommendations of the Advisory Committee on Immuniza-tion Practices (ACIP). MMWR Recomm. Rep. 2010, 59. [Google Scholar]
- CDC. Use of Japanese encephalitis vaccine in children: Recommendations of the Advisory Committee on Immunization Practices, 2013. MMWR Morb. Mortal Wkly. Rep. 2013, 62, 898–900. [Google Scholar]
- Putri, W.C.W.S.; Sawitri, A.A.S.; Yuliyatni, P.C.D.; Ariawan, I.M.D.; Meyta, H.; Labiba, S.U.; Suwarba, I.G.N.M.; Sutarsa, I.N. Cost-effectiveness analysis of Japanese Encephalitis (JE) vaccination program in Bali Province, Indonesia. Vaccine 2023, 41, 6930–6940. [Google Scholar] [CrossRef] [PubMed]
- Carias, C.; Hills, S.L.; Kahn, E.B.; Adhikari, B.B.; Fischer, M.; Meltzer, M.I. Comparative economic analysis of strategies for Japanese encephali-tis vaccination of U.S. travelers. Vaccine 2020, 38, 3351–3357. [Google Scholar] [CrossRef]
- Amat, C.; Bellanger, A.P.; Bozon, F.; Léger, R.; Gbaguidi-Haore, H.; Marguet, P. Current practice of French health professionals regarding Jap-anese encephalitis vaccination. Med. Mal. Infect. 2019, 49, 602–606. [Google Scholar] [CrossRef]
- Mills, D.J.; Lau, C.L.; Furuya-Kanamori, L. Low uptake of Japanese encephalitis vaccination among Australian travellers. J. Travel. Med. 2021, 28, taaa232. [Google Scholar] [CrossRef]
- Hills, S.L.; Walter, E.B.; Atmar, R.L.; Fischer, M. Japanese Encephalitis Vaccine: Recommendations of the Advisory Committee on Immun-ization Practices. MMWR Recomm. Rep. 2019, 68, 1–33. [Google Scholar] [CrossRef]
- Yu, Y. Phenotypic and genotypic characteristics of Japanese encephalitis attenuated live vaccine virus SA14-14-2 and their stabilities. Vaccine 2010, 28, 3635–3641. [Google Scholar] [CrossRef] [PubMed]
- Package-Insert-and-Patient-Information-IXIARO_0PDF. Available online: www.fda.gov (accessed on 11 October 2023).
- Jones, T. ChimeriVax-JE. Acambis. Curr. Opin. Investig. Drugs 2003, 4, 1019–1022. [Google Scholar] [PubMed]
- Chokephaibulkit, K.; Houillon, G.; Feroldi, E.; Bouckenooghe, A. Safety and immunogenicity of a live attenuated Japanese encephalitis chimeric virus vaccine (IMOJEV®) in children. Expert. Rev. Vaccines 2016, 15, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.substipharm.com/biologics-vaccins/ (accessed on 16 October 2023).
- van den Elsen, K.; Chew, B.L.A.; Ho, J.S.; Luo, D. Flavivirus nonstructural proteins and replication complexes as antiviral drug targets. Curr. Opin. Virol. 2023, 59, 101305. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monath, T.P. Japanese Encephalitis: Risk of Emergence in the United States and the Resulting Impact. Viruses 2024, 16, 54. https://doi.org/10.3390/v16010054
Monath TP. Japanese Encephalitis: Risk of Emergence in the United States and the Resulting Impact. Viruses. 2024; 16(1):54. https://doi.org/10.3390/v16010054
Chicago/Turabian StyleMonath, Thomas P. 2024. "Japanese Encephalitis: Risk of Emergence in the United States and the Resulting Impact" Viruses 16, no. 1: 54. https://doi.org/10.3390/v16010054
APA StyleMonath, T. P. (2024). Japanese Encephalitis: Risk of Emergence in the United States and the Resulting Impact. Viruses, 16(1), 54. https://doi.org/10.3390/v16010054