
Fast and Ultrasensitive Detection of Monkeypox by a Pyrococcus furiosus Argonaute System Coupled with a Short Amplification


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Expression and Purification of PfAgo
2.3. Confirmation of the Cleavage Activity of the Recombinant PfAgo
2.4. PCR of the 79 bp Amplicon
2.5. The Procedure of the PfAgo-MPXV System
2.6. Determination of the Copy Number of Extracted MPXV DNA by ddPCR
2.7. The Detection of MPXV in Simulated Samples
2.8. Statistics and Reproducibility
3. Results
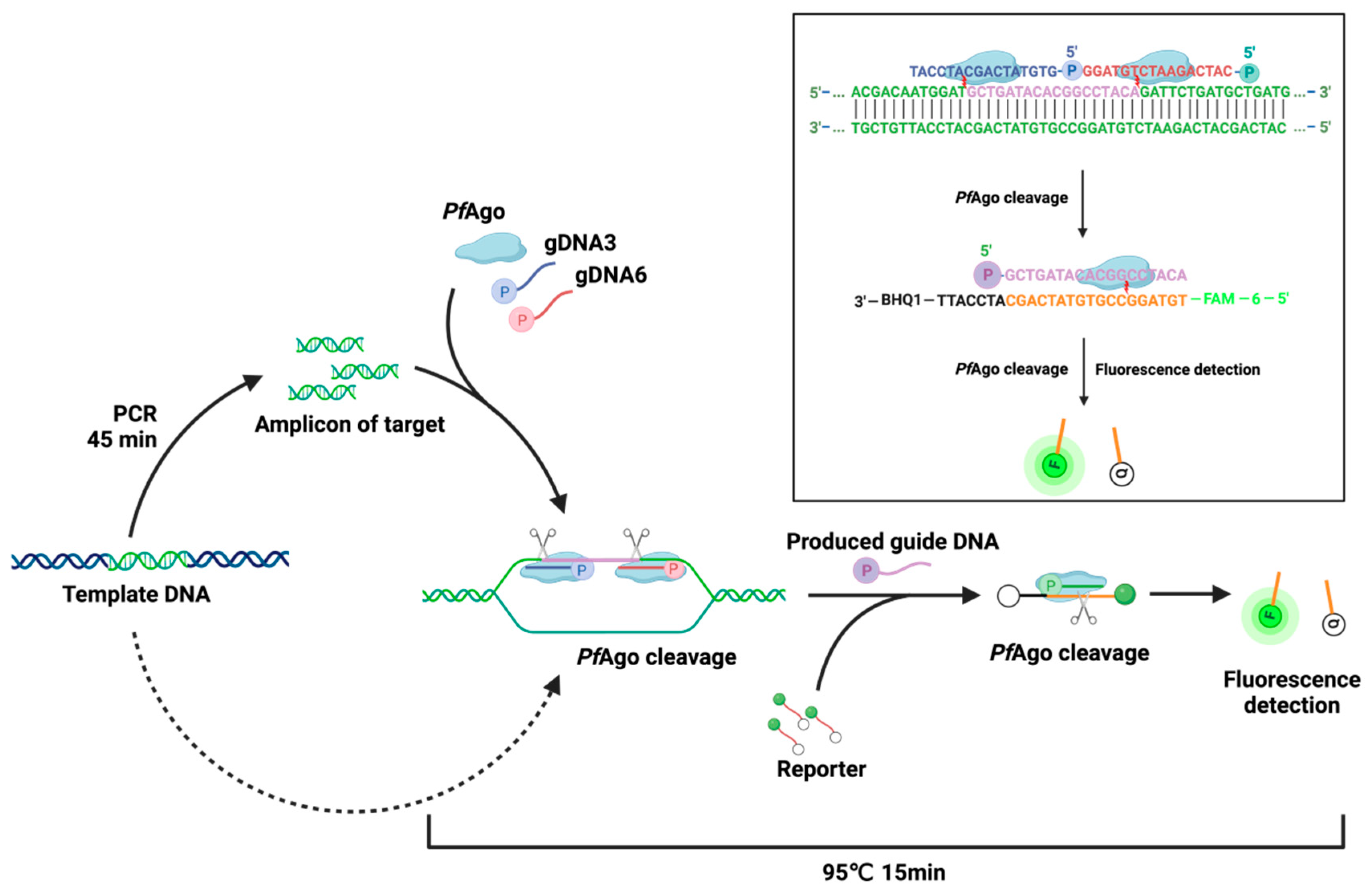
3.1. Design of the PfAgo-MPXV System for the Detection of MPXV
3.2. Evaluation of the PfAgo-MPXV Platform
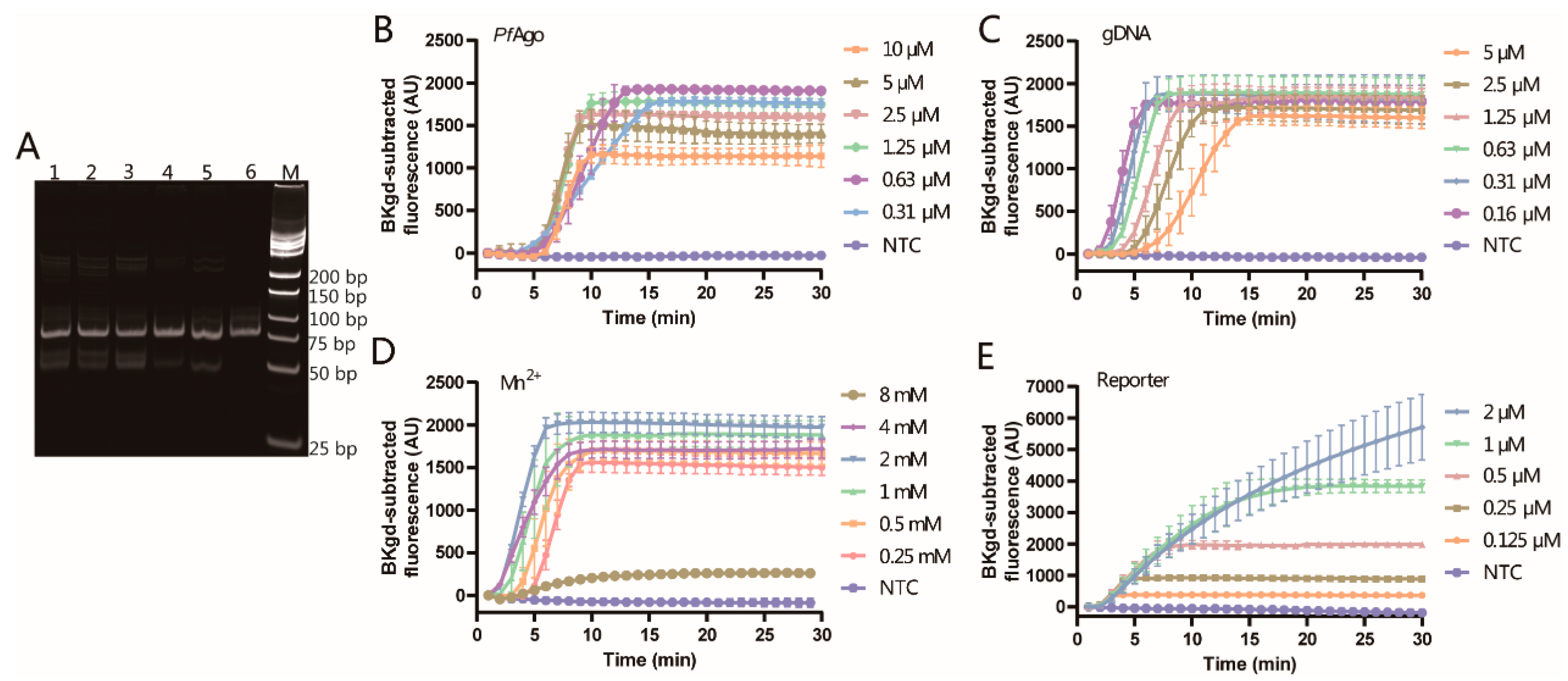
3.3. Optimization of the PfAgo-MPXV System
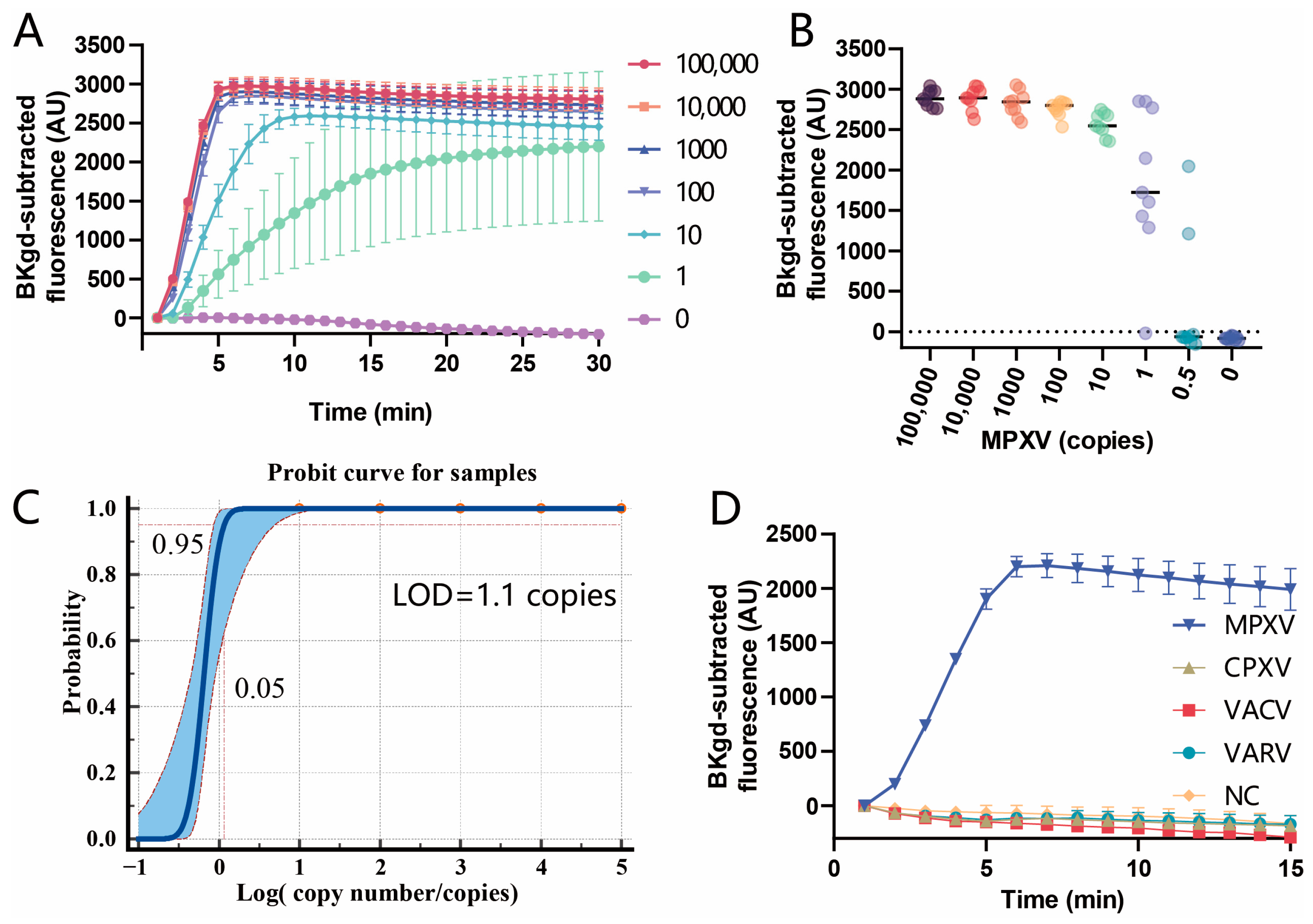
3.4. Sensitivity and Specificity of the PfAgo-MPXV System
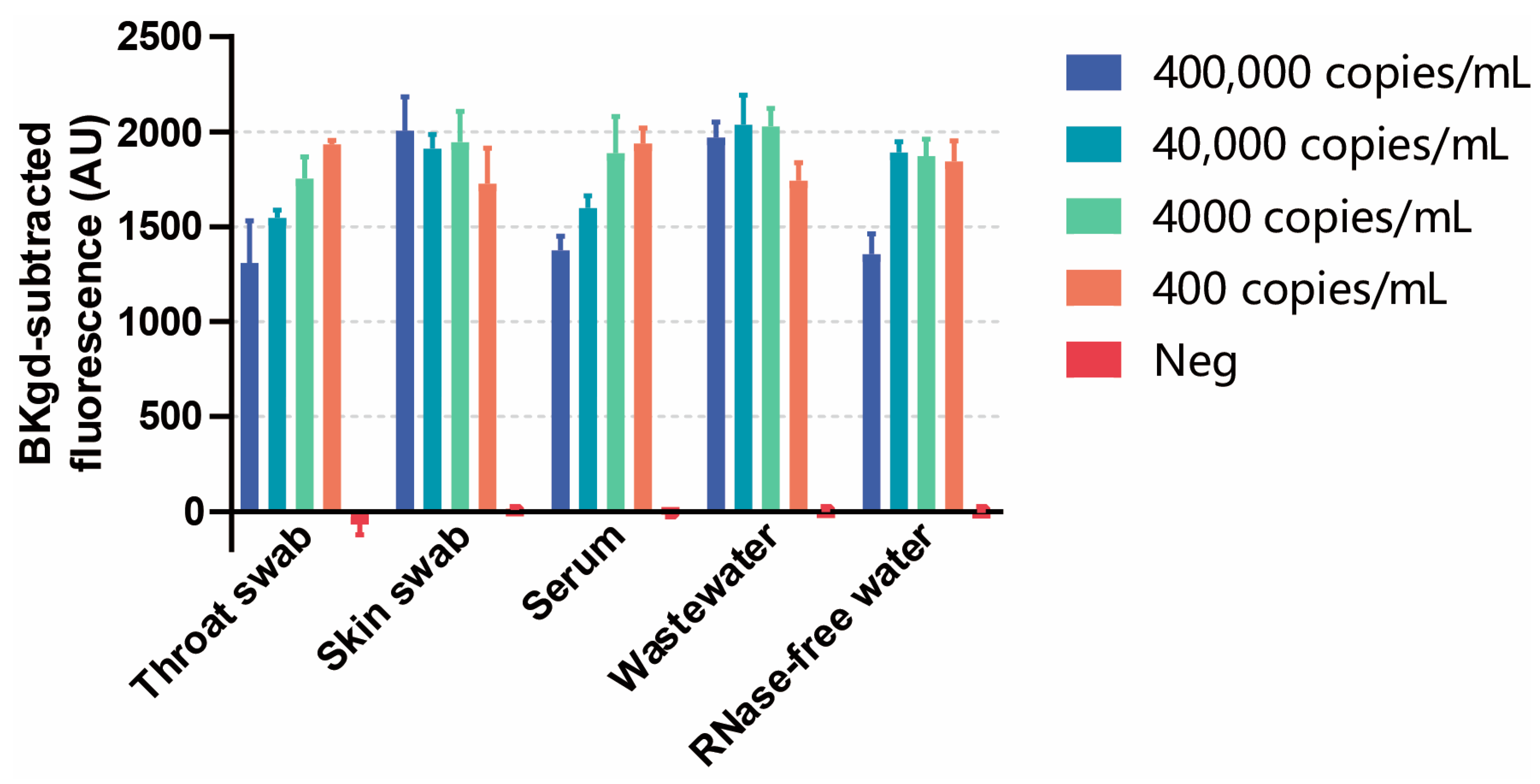
3.5. Detection of MPXV in Simulated Samples
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beer, E.M.; Rao, V.B. A systematic review of the epidemiology of human monkeypox outbreaks and implications for outbreak strategy. PLoS Neglected Trop. Dis. 2019, 13, e0007791. [Google Scholar] [CrossRef]
- World Health Organization. Multi-Country Outbreak of Mpox, External Situation Report 31. Available online: https://www.who.int/publications/m/item/multi-country-outbreak-of-mpox--external-situation-report-31 (accessed on 22 December 2023).
- Zhao, H.; Wang, W.; Zhao, L.; Ye, S.; Song, J.; Lu, R.; Zong, H.; Wu, C.; Huang, W.; Huang, B.; et al. The First Imported Case of Monkeypox in the Mainland of China—Chongqing Municipality, China, September 16, 2022. China CDC Wkly. 2022, 4, 853–854. [Google Scholar] [CrossRef] [PubMed]
- Sharif, N.; Sharif, N.; Alzahrani, K.J.; Halawani, I.F.; Alzahrani, F.M.; Diez, I.T.; Lipari, V.; Flores, M.A.L.; Parvez, A.K.; Dey, S.K. Molecular epidemiology, transmission and clinical features of 2022-mpox outbreak: A systematic review. Health Sci. Rep. 2023, 6, e1603. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, R.C.; Wang, C.; Hatcher, E.L.; Lefkowitz, E.J. Orthopoxvirus genome evolution: The role of gene loss. Viruses 2010, 2, 1933–1967. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, Z. Mpox: A review of laboratory detection techniques. Arch. Virol. 2023, 168, 221. [Google Scholar] [CrossRef] [PubMed]
- Swarts, D.C.; Hegge, J.W.; Hinojo, I.; Shiimori, M.; Ellis, M.A.; Dumrongkulraksa, J.; Terns, R.M.; Terns, M.P.; van der Oost, J. Argonaute of the archaeon Pyrococcus furiosus is a DNA-guided nuclease that targets cognate DNA. Nucleic Acids Res. 2015, 43, 5120–5129. [Google Scholar] [CrossRef] [PubMed]
- Swarts, D.C.; Jore, M.M.; Westra, E.R.; Zhu, Y.; Janssen, J.H.; Snijders, A.P.; Wang, Y.; Patel, D.J.; Berenguer, J.; Brouns, S.J.J.; et al. DNA-guided DNA interference by a prokaryotic Argonaute. Nature 2014, 507, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Willkomm, S.; Oellig, C.A.; Zander, A.; Restle, T.; Keegan, R.; Grohmann, D.; Schneider, S. Structural and mechanistic insights into an archaeal DNA-guided Argonaute protein. Nat. Microbiol. 2017, 2, 17035. [Google Scholar] [CrossRef]
- Enghiad, B.; Zhao, H. Programmable DNA-Guided Artificial Restriction Enzymes. ACS Synth. Biol. 2017, 6, 752–757. [Google Scholar] [CrossRef]
- Wang, L.; He, R.; Lv, B.; Yu, X.; Liu, Y.; Yang, J.; Li, W.; Wang, Y.; Zhang, H.; Yan, G.; et al. Pyrococcus furiosus Argonaute coupled with modified ligase chain reaction for detection of SARS-CoV-2 and HPV. Talanta 2021, 227, 122154. [Google Scholar] [CrossRef]
- Wang, F.; Yang, J.; He, R.; Yu, X.; Chen, S.; Liu, Y.; Wang, L.; Li, A.; Liu, L.; Zhai, C.; et al. PfAgo-based detection of SARS-CoV-2. Biosens. Bioelectron. 2021, 177, 112932. [Google Scholar] [CrossRef]
- Liu, Q.; Guo, X.; Xun, G.; Li, Z.; Chong, Y.; Yang, L.; Wang, H.; Zhang, F.; Luo, S.; Cui, L.; et al. Argonaute integrated single-tube PCR system enables supersensitive detection of rare mutations. Nucleic Acids Res. 2021, 49, e75. [Google Scholar] [CrossRef]
- Ye, X.; Zhou, H.; Guo, X.; Liu, D.; Li, Z.; Sun, J.; Huang, J.; Liu, T.; Zhao, P.; Xu, H.; et al. Argonaute-integrated isothermal amplification for rapid, portable, multiplex detection of SARS-CoV-2 and influenza viruses. Biosens Bioelectron 2022, 207, 114169. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Qiu, L.; Luo, T.; Lu, Z.; Wang, X.; Hong, Q.; Luo, J.; Ma, L.; Wang, Y.; Dong, Y. Novel Nucleic Acid Detection for Human Parvovirus B19 Based on Pyrococcus furiosus Argonaute Protein. Viruses 2023, 15, 595. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xia, W.; Zhao, W.; Hao, P.; Wang, Z.; Yu, X.; Shentu, X.; Sun, K. RT-RPA-PfAgo System: A Rapid, Sensitive, and Specific Multiplex Detection Method for Rice-Infecting Viruses. Biosensors 2023, 13, 941. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.; Xun, G.; Lane, S.T.; Petrov, V.A.; Zhao, H.; Nguyen, T.H. Portable, single nucleotide polymorphism-specific duplex assay for virus surveillance in wastewater. Sci. Total Environ. 2023, 912, 168701. [Google Scholar] [CrossRef]
- He, R.; Wang, L.; Wang, F.; Li, W.; Liu, Y.; Li, A.; Wang, Y.; Mao, W.; Zhai, C.; Ma, L. Pyrococcus furiosus Argonaute-mediated nucleic acid detection. Chem. Commun. 2019, 55, 13219–13222. [Google Scholar] [CrossRef]
- He, R.; Wang, L.; Wang, F.; Yang, J.; Yu, X.; Wang, Y.; Liu, Z.; Li, C.; Ma, L. Combination of ultrashort PCR and Pyrococcus furiosus Argonaute for DNA detection. Analyst 2021, 147, 35–39. [Google Scholar] [CrossRef]
- Jang, H.; Song, J.; Kim, S.; Byun, J.H.; Lee, K.G.; Park, K.H.; Woo, E.; Lim, E.K.; Jung, J.; Kang, T. ANCA: Artificial nucleic acid circuit with argonaute protein for one-step isothermal detection of antibiotic-resistant bacteria. Nat. Commun. 2023, 14, 8033. [Google Scholar] [CrossRef]
- Shchelkunov, S.N.; Shcherbakov, D.N.; Maksyutov, R.A.; Gavrilova, E.V. Species-specific identification of variola, monkeypox, cowpox, and vaccinia viruses by multiplex real-time PCR assay. J. Virol. Methods 2011, 175, 163–169. [Google Scholar] [CrossRef]
- Stokdyk, J.P.; Firnstahl, A.D.; Spencer, S.K.; Burch, T.R.; Borchardt, M.A. Determining the 95% limit of detection for waterborne pathogen analyses from primary concentration to qPCR. Water Res. 2016, 96, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Lum, F.M.; Torres-Ruesta, A.; Tay, M.Z.; Lin, R.T.P.; Lye, D.C.; Renia, L.; Ng, L.F.P. Monkeypox: Disease epidemiology, host immunity and clinical interventions. Nat. Rev. Immunol. 2022, 22, 597–613. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Guo, B.; Wang, Y.; Zhao, C.; Zhang, X.; Wang, Y.; Tang, Y.; Shen, H.; Wang, P.; Gao, S. Pyrococcus furiosus Argonaute Combined with Recombinase Polymerase Amplification for Rapid and Sensitive Detection of Enterocytozoon hepatopenaei. J. Agric. Food Chem. 2023, 71, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Seo, M.R.; Chung, Y.J. Development of reverse-transcription loop-mediated isothermal amplification assays for point-of-care testing of human influenza virus subtypes H1N1 and H3N2. Genom. Inf. 2022, 20, e46. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, P.; Zhou, W.; Wei, H.; Yu, J. Fast and Ultrasensitive Detection of Monkeypox by a Pyrococcus furiosus Argonaute System Coupled with a Short Amplification. Viruses 2024, 16, 382. https://doi.org/10.3390/v16030382
He P, Zhou W, Wei H, Yu J. Fast and Ultrasensitive Detection of Monkeypox by a Pyrococcus furiosus Argonaute System Coupled with a Short Amplification. Viruses. 2024; 16(3):382. https://doi.org/10.3390/v16030382
Chicago/Turabian StyleHe, Ping, Wenhao Zhou, Hongping Wei, and Junping Yu. 2024. "Fast and Ultrasensitive Detection of Monkeypox by a Pyrococcus furiosus Argonaute System Coupled with a Short Amplification" Viruses 16, no. 3: 382. https://doi.org/10.3390/v16030382
APA StyleHe, P., Zhou, W., Wei, H., & Yu, J. (2024). Fast and Ultrasensitive Detection of Monkeypox by a Pyrococcus furiosus Argonaute System Coupled with a Short Amplification. Viruses, 16(3), 382. https://doi.org/10.3390/v16030382