Poxvirus Exploitation of the Ubiquitin-Proteasome System

Abstract
:1. Introduction
2. Poxvirus Encoded Ubiquitin
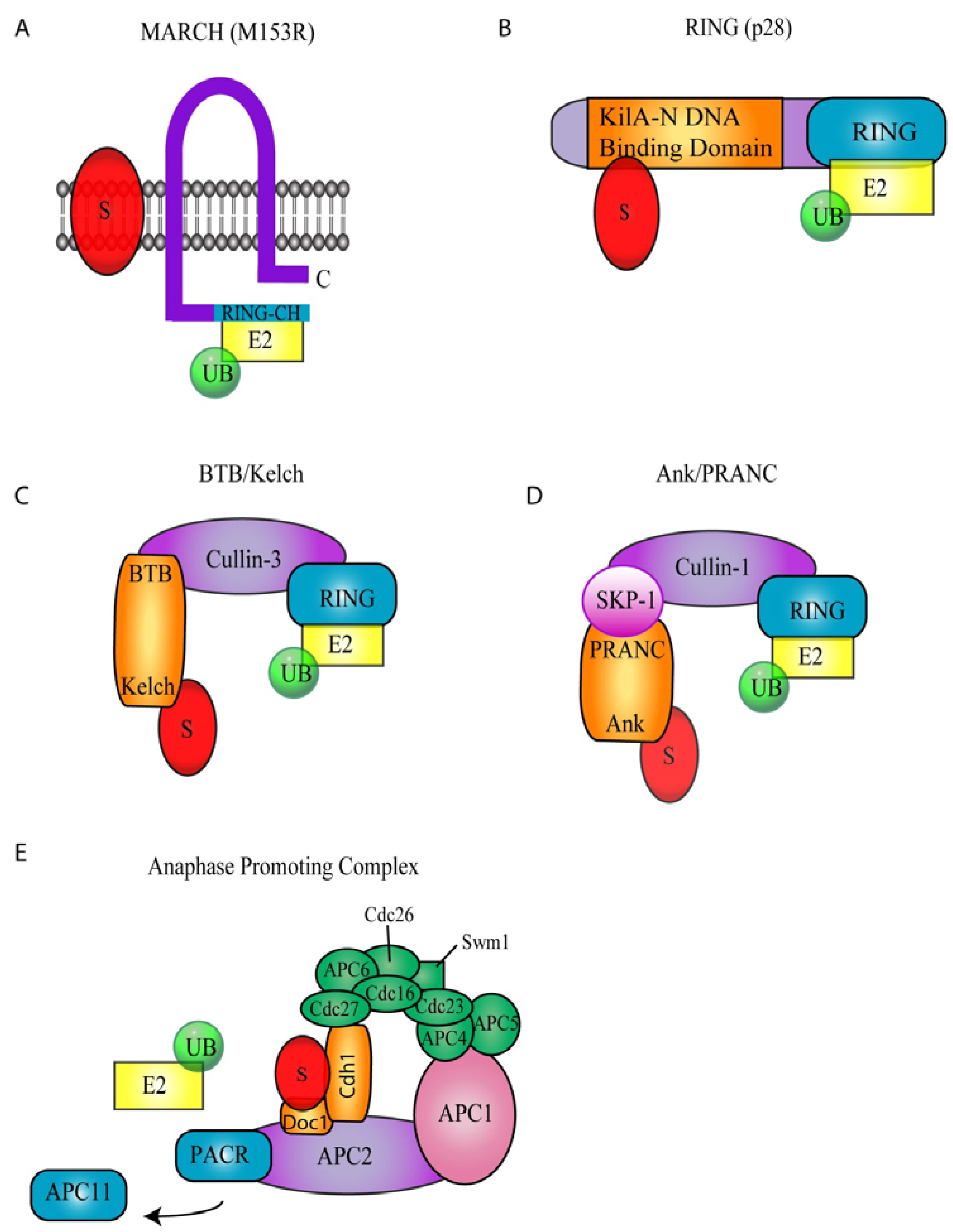
3. Poxvirus Encoded Ubiquitin Ligases
4. A Family of Poxvirus Encoded BTB/Kelch Proteins
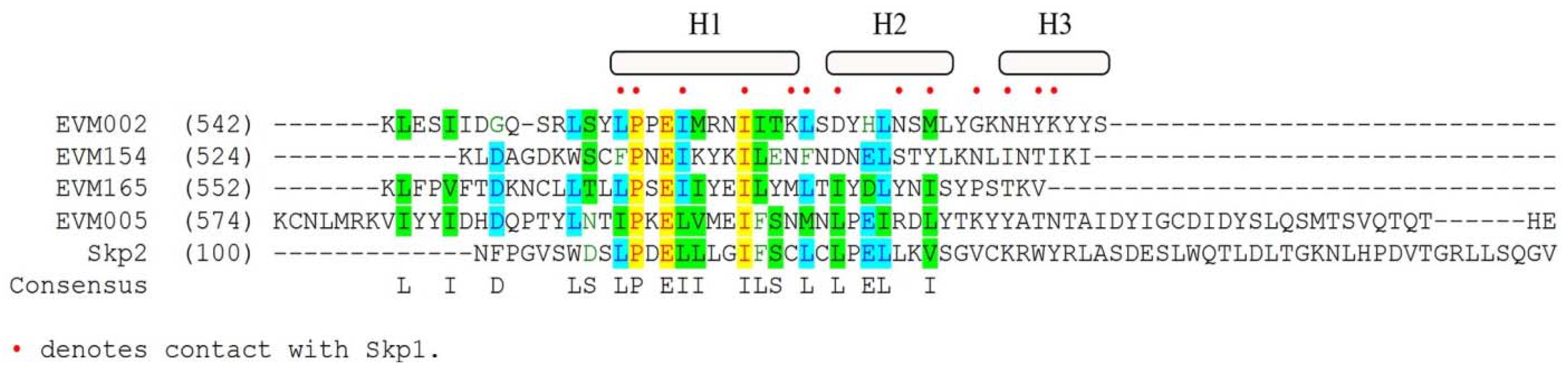
5. Poxvirus Encoded Ankyrin/PRANC Proteins
6. Regulation of the APC/C by Poxviruses
7. Role of the Ubiquitin-Proteasome System During Poxvirus Infection
8. Conclusions
References and Notes
- Pickart, C.M.; Fushman, D. Polyubiquitin chains: Polymeric protein signals. Curr. Opin. Chem. Biol. 2004, 8, 610–616. [Google Scholar] [CrossRef]
- Chen, Z.J.; Sun, L.J. Nonproteolytic functions of ubiquitin in cell signaling. Mol. Cell 2009, 33, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Giles, J. Chemistry Nobel for trio who revealed molecular death-tag. Nature 2004, 431, 729. [Google Scholar] [CrossRef] [PubMed]
- Komander, D. The emerging complexity of protein ubiquitination. Biochem. Soc. Trans. 2009, 37, 937–953. [Google Scholar] [CrossRef] [PubMed]
- Cadwell, K.; Coscoy, L. Ubiquitination on nonlysine residues by a viral E3 ubiquitin ligase. Science 2005, 309, 127–130. [Google Scholar] [CrossRef]
- Iwai, K.; Tokunaga, F. Linear polyubiquitination: A new regulator of NF-kappaB activation. EMBO Rep. 2009, 10, 706–713. [Google Scholar] [CrossRef]
- Moss, B. Poxviridae: The viruses and their replication. In Fields Virology; Fields, B.N., Knipe, D.M., Howley, P.M., Eds.; Lippincott-Raven Publishers: Philadelphia, PA, USA, 1996; pp. 2637–2671. [Google Scholar]
- Wehrle, P.F. A reality in our time—Certification of the global eradication of smallpox. J. Infect. Dis. 1980, 142, 636–638. [Google Scholar] [CrossRef]
- Jacobs, B.L.; Langland, J.O.; Kibler, K.V.; Denzler, K.L.; White, S.D.; Holechek, S.A.; Wong, S.; Huynh, T.; Baskin, C.R. Vaccinia virus vaccines: Past, present and future. Antivir. Res. 2009, 84, 1–13. [Google Scholar] [CrossRef]
- Johnston, J.B.; McFadden, G. Technical knockout: Understanding poxvirus pathogenesis by selectively deleting viral immunomodulatory genes. Cell. Microbiol. 2004, 6, 695–705. [Google Scholar] [CrossRef]
- Seet, B.T.; Johnston, J.B.; Brunetti, C.R.; Barrett, J.W.; Everett, H.; Cameron, C.; Sypula, J.; Nazarian, S.H.; Lucas, A.; McFadden, G. Poxviruses and immune evasion. Annu. Rev. Immunol. 2003, 21, 377–423. [Google Scholar] [CrossRef]
- Barry, M.; Fruh, K. Viral modulators of cullin RING ubiquitin ligases: Culling the host defense. Sci. STKE 2006, 2006, pe21. [Google Scholar] [CrossRef]
- Isaacson, M.K.; Ploegh, H.L. Ubiquitination, ubiquitin-like modifiers, and deubiquitination in viral infection. Cell Host Microbe 2009, 5, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Randow, F.; Lehner, P.J. Viral avoidance and exploitation of the ubiquitin system. Nat. Cell Biol. 2009, 11, 527–534. [Google Scholar] [CrossRef]
- Shchelkunov, S.N. Interaction of orthopoxviruses with the cellular ubiquitin-ligase system. Virus Genes 2010. [Google Scholar] [CrossRef]
- Zhang, L.; Villa, N.Y.; McFadden, G. Interplay between poxviruses and the cellular ubiquitin/ubiquitin-like pathways. FEBS Lett. 2009, 583, 607–614. [Google Scholar] [CrossRef]
- Weissman, A.M. Themes and variations on ubiquitylation. Nat. Rev. Mol. Cell. Biol. 2001, 2, 169–178. [Google Scholar] [CrossRef]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef]
- Schlesinger, M.J.; Bond, U. Ubiquitin genes. Oxf. Surv. Eukaryot. Gene. 1987, 4, 77–91. [Google Scholar]
- Hicke, L.; Schubert, H.L.; Hill, C.P. Ubiquitin-binding domains. Nat. Rev. Mol. Cell. Biol. 2005, 6, 610–621. [Google Scholar] [CrossRef]
- Afonso, C.L.; Tulman, E.R.; Lu, Z.; Oma, E.; Kutish, G.F.; Rock, D.L. The genome of Melanoplus sanguinipes entomopoxvirus. J. Virol. 1999, 73, 533–552. [Google Scholar] [CrossRef]
- Bawden, A.L.; Glassberg, K.J.; Diggans, J.; Shaw, R.; Farmerie, W.; Moyer, R.W. Complete genomic sequence of the Amsacta moorei entomopoxvirus: Analysis and comparison with other poxviruses. Virology 2000, 274, 120–139. [Google Scholar] [CrossRef] [PubMed]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. The genome of canarypox virus. J. Virol. 2004, 78, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.L.; Tulman, E.R.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. The genome of fowlpox virus. J. Virol. 2000, 74, 3815–3831. [Google Scholar] [CrossRef] [PubMed]
- Reilly, L.M.; Guarino, L.A. The viral ubiquitin gene of Autographa californica nuclear polyhedrosis virus is not essential for viral replication. Virology 1996, 218, 243–247. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl. Acid. Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Lefkowitz, E.J.; Upton, C.; Changayil, S.S.; Buck, C.; Traktman, P.; Buller, R.M. Poxvirus Bioinformatics Resource Center: A comprehensive Poxviridae informational and analytical resource. Nucl. Acid. Res. 2005, 33, D311–D316. [Google Scholar] [CrossRef]
- Chung, C.S.; Chen, C.H.; Ho, M.Y.; Huang, C.Y.; Liao, C.L.; Chang, W. Vaccinia virus proteome: Identification of proteins in vaccinia virus intracellular mature virion particles. J. Virol. 2006, 80, 2127–2140. [Google Scholar] [CrossRef]
- Guarino, L.A.; Smith, G.; Dong, W. Ubiquitin is attached to membranes of baculovirus particles by a novel type of phospholipid anchor. Cell 1995, 80, 301–309. [Google Scholar] [CrossRef]
- Webb, J.H.; Mayer, R.J.; Dixon, L.K. A lipid modified ubiquitin is packaged into particles of several enveloped viruses. FEBS Lett. 1999, 444, 136–139. [Google Scholar] [CrossRef]
- Teale, A.; Campbell, S.; Van Buuren, N.; Magee, W.C.; Watmough, K.; Couturier, B.; Shipclark, R.; Barry, M. Orthopoxviruses require a functional ubiquitin-proteasome system for productive replication. J. Virol. 2009, 83, 2099–2108. [Google Scholar] [CrossRef]
- Satheshkumar, P.S.; Anton, L.C.; Sanz, P.; Moss, B. Inhibition of the ubiquitin-proteasome system prevents vaccinia virus DNA replication and expression of intermediate and late genes. J. Virol. 2009, 83, 2469–2479. [Google Scholar] [CrossRef]
- Haas, A.L.; Katzung, D.J.; Reback, P.M.; Guarino, L.A. Functional characterization of the ubiquitin variant encoded by the baculovirus Autographa californica. Biochemistry 1996, 35, 5385–5394. [Google Scholar] [CrossRef]
- Guerin, J.L.; Gelfi, J.; Boullier, S.; Delverdier, M.; Bellanger, F.A.; Bertagnoli, S.; Drexler, I.; Sutter, G.; Messud-Petit, F. Myxoma virus leukemia-associated protein is responsible for major histocompatibility complex class I and Fas-CD95 down-regulation and defines scrapins, a new group of surface cellular receptor abductor proteins. J. Virol. 2002, 76, 2912–2923. [Google Scholar] [CrossRef]
- Huang, J.; Huang, Q.; Zhou, X.; Shen, M.M.; Yen, A.; Yu, S.X.; Dong, G.; Qu, K.; Huang, P.; Anderson, E.M.; Daniel-Issakani, S.; Buller, R.M.; Payan, D.G.; Lu, H.H. The poxvirus p28 virulence factor is an E3 ubiquitin ligase. J. Biol. Chem. 2004, 279, 54110–54116. [Google Scholar] [CrossRef]
- Mansouri, M.; Bartee, E.; Gouveia, K.; Hovey Nerenberg, B.T.; Barrett, J.; Thomas, L.; Thomas, G.; McFadden, G.; Fruh, K. The PHD/LAP-domain protein M153R of myxomavirus is a ubiquitin ligase that induces the rapid internalization and lysosomal destruction of CD4. J. Virol. 2003, 77, 1427–1440. [Google Scholar] [CrossRef]
- Nerenberg, B.T.; Taylor, J.; Bartee, E.; Gouveia, K.; Barry, M.; Fruh, K. The poxviral RING protein p28 is a ubiquitin ligase that targets ubiquitin to viral replication factories. J. Virol. 2005, 79, 597–601. [Google Scholar] [CrossRef]
- Nathan, J.A.; Lehner, P.J. The trafficking and regulation of membrane receptors by the RING-CH ubiquitin E3 ligases. Exp. Cell Res. 2009, 315, 1593–1600. [Google Scholar] [CrossRef]
- Fruh, K.; Bartee, E.; Gouveia, K.; Mansouri, M. Immune evasion by a novel family of viral PHD/LAP-finger proteins of gamma-2 herpesviruses and poxviruses. Virus Res. 2002, 88, 55–69. [Google Scholar] [CrossRef]
- Bartee, E.; Mansouri, M.; Hovey Nerenberg, B.T.; Gouveia, K.; Fruh, K. Downregulation of major histocompatibility complex class I by human ubiquitin ligases related to viral immune evasion proteins. J. Virol. 2004, 78, 1109–1120. [Google Scholar] [CrossRef]
- Boshkov, L.K.; Macen, J.L.; McFadden, G. Virus-induced loss of class I MHC antigens from the surface of cells infected with myxoma virus and malignant rabbit fibroma virus. J. Immunol. 1992, 148, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, M.C.; Wang, H.; Barry, M.; McFadden, G. Endosomal/lysosomal retention and degradation of major histocompatibility complex class I molecules is induced by myxoma virus. Virology 1999, 261, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Senkevich, T.G.; Koonin, E.V.; Buller, R.M. A poxvirus protein with a RING zinc finger motif is of crucial importance for virulence. Virology 1994, 198, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Senkevich, T.G.; Wolffe, E.J.; Buller, R.M. Ectromelia virus RING finger protein is localized in virus factories and is required for virus replication in macrophages. J. Virol. 1995, 69, 4103–4111. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Koonin, E.V.; Aravind, L. Extensive domain shuffling in transcription regulators of DNA viruses and implications for the origin of fungal APSES transcription factors. Genome Biol. 2002, 3, RESEARCH0012. [Google Scholar] [CrossRef]
- Brick, D.J.; Burke, R.D.; Schiff, L.; Upton, C. Shope fibroma virus RING finger protein N1R binds DNA and inhibits apoptosis. Virology 1998, 249, 42–51. [Google Scholar] [CrossRef]
- Mottet, K. University of Alberta, Edmonton, Canada. Unpublished work. 2010. [Google Scholar]
- Nicholls, R.D.; Gray, T.A. Cellular source of the poxviral N1R/p28 gene family. Virus Genes 2004, 29, 359–364. [Google Scholar] [CrossRef]
- Brick, D.J.; Burke, R.D.; Minkley, A.A.; Upton, C. Ectromelia virus virulence factor p28 acts upstream of caspase-3 in response to UV light-induced apoptosis. J. Gen. Virol. 2000, 81, 1087–1097. [Google Scholar] [CrossRef]
- Bardwell, V.J.; Treisman, R. The POZ domain: a conserved protein-protein interaction motif. Genes Dev. 1994, 8, 1664–1677. [Google Scholar] [CrossRef]
- Melnick, A.; Ahmad, K.F.; Arai, S.; Polinger, A.; Ball, H.; Borden, K.L.; Carlile, G.W.; Prive, G.G.; Licht, J.D. In-depth mutational analysis of the promyelocytic leukemia zinc finger BTB/POZ domain reveals motifs and residues required for biological and transcriptional functions. Mol. Cell. Biol. 2000, 20, 6550–6567. [Google Scholar] [CrossRef]
- Kang, M.I.; Kobayashi, A.; Wakabayashi, N.; Kim, S.G.; Yamamoto, M. Scaffolding of Keap1 to the actin cytoskeleton controls the function of Nrf2 as key regulator of cytoprotective phase 2 genes. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 2046–2051. [Google Scholar] [CrossRef]
- Furukawa, M.; He, Y.J.; Borchers, C.; Xiong, Y. Targeting of protein ubiquitination by BTB-Cullin 3-Roc1 ubiquitin ligases. Nat. Cell Biol. 2003, 5, 1001–1007. [Google Scholar] [CrossRef]
- Geyer, R.; Wee, S.; Anderson, S.; Yates, J.; Wolf, D.A. BTB/POZ domain proteins are putative substrate adaptors for cullin 3 ubiquitin ligases. Mol. Cell 2003, 12, 783–790. [Google Scholar] [CrossRef]
- Pintard, L.; Willis, J.H.; Willems, A.; Johnson, J.L.; Srayko, M.; Kurz, T.; Glaser, S.; Mains, P.E.; Tyers, M.; Bowerman, B.; Peter, M. The BTB protein MEL-26 is a substrate-specific adaptor of the CUL-3 ubiquitin-ligase. Nature 2003, 425, 311–316. [Google Scholar] [CrossRef]
- Xu, L.; Wei, Y.; Reboul, J.; Vaglio, P.; Shin, T.H.; Vidal, M.; Elledge, S.J.; Harper, J.W. BTB proteins are substrate-specific adaptors in an SCF-like modular ubiquitin ligase containing CUL-3. Nature 2003, 425, 316–321. [Google Scholar] [CrossRef]
- Aravind, L.; Koonin, E.V. Fold prediction and evolutionary analysis of the POZ domain: Structural and evolutionary relationship with the potassium channel tetramerization domain. J. Mol. Biol. 1999, 285, 1353–1361. [Google Scholar] [CrossRef]
- Schulman, B.A.; Carrano, A.C.; Jeffrey, P.D.; Bowen, Z.; Kinnucan, E.R.; Finnin, M.S.; Elledge, S.J.; Harper, J.W.; Pagano, M.; Pavletich, N.P. Insights into SCF ubiquitin ligases from the structure of the Skp1-Skp2 complex. Nature 2000, 408, 381–386. [Google Scholar] [CrossRef]
- Adams, J.; Kelso, R.; Cooley, L. The kelch repeat superfamily of proteins: Propellers of cell function. Trends Cell Biol. 2000, 10, 17–24. [Google Scholar] [CrossRef]
- Shchelkunov, S.; Totmenin, A.; Kolosova, I. Species-specific differences in organization of orthopoxvirus kelch-like proteins. Virus Genes 2002, 24, 157–162. [Google Scholar] [CrossRef]
- Kotwal, G.J.; Moss, B. Analysis of a large cluster of nonessential genes deleted from a vaccinia virus terminal transposition mutant. Virology 1988, 167, 524–537. [Google Scholar] [CrossRef]
- Shchelkunov, S.N.; Safronov, P.F.; Totmenin, A.V.; Petrov, N.A.; Ryazankina, O.I.; Gutorov, V.V.; Kotwal, G.J. The genomic sequence analysis of the left and right species-specific terminal region of a cowpox virus strain reveals unique sequences and a cluster of intact ORFs for immunomodulatory and host range proteins. Virology 1998, 243, 432–460. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Danila, M.I.; Feng, Z.; Buller, R.M.; Wang, C.; Han, X.; Lefkowitz, E.J.; Upton, C. The genomic sequence of ectromelia virus, the causative agent of mousepox. Virology 2003, 317, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Shchelkunov, S.N.; Totmenin, A.V.; Safronov, P.F.; Mikheev, M.V.; Gutorov, V.V.; Ryazankina, O.I.; Petrov, N.A.; Babkin, I.V.; Uvarova, E.A.; Sandakhchiev, L.S.; Sisler, J.R.; Esposito, J.J.; Damon, I.K.; Jahrling, P.B.; Moss, B. Analysis of the monkeypox virus genome. Virology 2002, 297, 172–194. [Google Scholar] [CrossRef] [PubMed]
- Wilton, B.A.; Campbell, S.; Van Buuren, N.; Garneau, R.; Furukawa, M.; Xiong, Y.; Barry, M. Ectromelia virus BTB/kelch proteins, EVM150 and EVM167, interact with cullin-3-based ubiquitin ligases. Virology 2008, 374, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Wilton, B. University of Alberta, Edmonton, Canada. Unpublished work. 2009. [Google Scholar]
- Pires de Miranda, M.; Reading, P.C.; Tscharke, D.C.; Murphy, B.J.; Smith, G.L. The vaccinia virus kelch-like protein C2L affects calcium-independent adhesion to the extracellular matrix and inflammation in a murine intradermal model. J. Gen. Virol. 2003, 84, 2459–2471. [Google Scholar] [CrossRef]
- Beard, P.M.; Froggatt, G.C.; Smith, G.L. Vaccinia virus kelch protein A55 is a 64 kDa intracellular factor that affects virus-induced cytopathic effect and the outcome of infection in a murine intradermal model. J. Gen. Virol. 2006, 87, 1521–1529. [Google Scholar] [CrossRef]
- Froggatt, G.C.; Smith, G.L.; Beard, P.M. Vaccinia virus gene F3L encodes an intracellular protein that affects the innate immune response. J. Gen. Virol. 2007, 88, 1917–1921. [Google Scholar] [CrossRef]
- Kochneva, G.; Kolosova, I.; Maksyutova, T.; Ryabchikova, E.; Shchelkunov, S. Effects of deletions of kelch-like genes on cowpox virus biological properties. Arch. Virol. 2005, 150, 1857–1870. [Google Scholar] [CrossRef]
- Balinsky, C.A.; Delhon, G.; Afonso, C.L.; Risatti, G.R.; Borca, M.V.; French, R.A.; Tulman, E.R.; Geary, S.J.; Rock, D.L. Sheeppox virus kelch-like gene SPPV-019 affects virus virulence. J. Virol. 2007, 81, 11392–11401. [Google Scholar] [CrossRef]
- Cullinan, S.B.; Gordan, J.D.; Jin, J.; Harper, J.W.; Diehl, J.A. The Keap1-BTB protein is an adaptor that bridges Nrf2 to a Cul3-based E3 ligase: Oxidative stress sensing by a Cul3-Keap1 ligase. Mol. Cell. Biol. 2004, 24, 8477–8486. [Google Scholar] [CrossRef]
- Furukawa, M.; Xiong, Y. BTB protein Keap1 targets antioxidant transcription factor Nrf2 for ubiquitination by the Cullin 3-Roc1 ligase. Mol. Cell. Biol. 2005, 25, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.F.; Kuo, H.P.; Liu, M.; Chou, C.K.; Xia, W.; Du, Y.; Shen, J.; Chen, C.T.; Huo, L.; Hsu, M.C.; Li, C.W.; Ding, Q.; Liao, T.L.; Lai, C.C.; Lin, A.C.; Chang, Y.H.; Tsai, S.F.; Li, L.Y.; Hung, M.C. KEAP1 E3 ligase-mediated downregulation of NF-kappaB signaling by targeting IKKbeta. Mol. Cell 2009, 36, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Maerki, S.; Olma, M.H.; Staubli, T.; Steigemann, P.; Gerlich, D.W.; Quadroni, M.; Sumara, I.; Peter, M. The Cul3-KLHL21 E3 ubiquitin ligase targets aurora B to midzone microtubules in anaphase and is required for cytokinesis. J. Cell Biol. 2009, 187, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Villa, N.Y.; Rahman, M.M.; Smallwood, S.; Shattuck, D.; Neff, C.; Dufford, M.; Lanchbury, J.S.; Labaer, J.; McFadden, G. Analysis of vaccinia virus-host protein-protein interactions: Validations of yeast two-hybrid screenings. J. Proteome Res. 2009, 8, 4311–4318. [Google Scholar] [CrossRef] [PubMed]
- Al-Khodor, S.; Price, C.T.; Kalia, A.; Abu Kwaik, Y. Functional diversity of ankyrin repeats in microbial proteins. Trends Microbiol. 2010, 18, 132–139. [Google Scholar] [CrossRef]
- Mosavi, L.K.; Cammett, T.J.; Desrosiers, D.C.; Peng, Z.Y. The ankyrin repeat as molecular architecture for protein recognition. Protein Sci. 2004, 13, 1435–1448. [Google Scholar] [CrossRef]
- Sedgwick, S.G.; Smerdon, S.J. The ankyrin repeat: A diversity of interactions on a common structural framework. Trends Biochem. Sci. 1999, 24, 311–316. [Google Scholar] [CrossRef]
- Lux, S.E.; John, K.M.; Bennett, V. Analysis of cDNA for human erythrocyte ankyrin indicates a repeated structure with homology to tissue-differentiation and cell-cycle control proteins. Nature 1990, 344, 36–42. [Google Scholar] [CrossRef]
- Mercer, A.A.; Fleming, S.B.; Ueda, N. F-box-like domains are present in most poxvirus ankyrin repeat proteins. Virus Genes 2005, 31, 127–133. [Google Scholar] [CrossRef]
- Skowyra, D.; Craig, K.L.; Tyers, M.; Elledge, S.J.; Harper, J.W. F-box proteins are receptors that recruit phosphorylated substrates to the SCF ubiquitin-ligase complex. Cell 1997, 91, 209–219. [Google Scholar] [CrossRef]
- Zheng, N.; Schulman, B.A.; Song, L.; Miller, J.J.; Jeffrey, P.D.; Wang, P.; Chu, C.; Koepp, D.M.; Elledge, S.J.; Pagano, M.; Conaway, R.C.; Conaway, J.W.; Harper, J.W.; Pavletich, N.P. Structure of the Cul1-Rbx1-Skp1-F boxSkp2 SCF ubiquitin ligase complex. Nature 2002, 416, 703–709. [Google Scholar] [CrossRef]
- Cardozo, T.; Pagano, M. The SCF ubiquitin ligase: Insights into a molecular machine. Nat. Rev. Mol. Cell. Biol. 2004, 5, 739–751. [Google Scholar] [CrossRef]
- Petroski, M.D.; Deshaies, R.J. Function and regulation of cullin-RING ubiquitin ligases. Nat. Rev. Mol. Cell. Biol. 2005, 6, 9–20. [Google Scholar] [CrossRef]
- Werren, J.H.; Richards, S.; Desjardins, C.A.; Niehuis, O.; Gadau, J.; Colbourne, J.K.; Beukeboom, L.W.; Desplan, C.; Elsik, C.G.; Grimmelikhuijzen, C.J.; et al. Functional and evolutionary insights from the genomes of three parasitoid Nasonia species. Science 2010, 327, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Sonnberg, S.; Fleming, S.B.; Mercer, A.A. A truncated two-alpha-helix F-box present in poxvirus ankyrin-repeat proteins is sufficient for binding the SCF1 ubiquitin ligase complex. J. Gen. Virol. 2009, 90, 1224–1228. [Google Scholar] [CrossRef]
- Van Buuren, N.; Couturier, B.; Xiong, Y.; Barry, M. Ectromelia virus encodes a novel family of F-box proteins that interact with the SCF complex. J. Virol. 2008, 82, 9917–9927. [Google Scholar] [CrossRef]
- Chang, S.J.; Hsiao, J.C.; Sonnberg, S.; Chiang, C.T.; Yang, M.H.; Tzou, D.L.; Mercer, A.A.; Chang, W. Poxvirus host range protein CP77 contains an F-box-like domain that is necessary to suppress NF-kappaB activation by tumor necrosis factor alpha but is independent of its host range function. J. Virol. 2009, 83, 4140–4152. [Google Scholar] [CrossRef]
- Piessevaux, J.; Lavens, D.; Peelman, F.; Tavernier, J. The many faces of the SOCS box. Cytokine Growth Factor Rev. 2008, 19, 371–381. [Google Scholar] [CrossRef]
- Sonnberg, S.; Seet, B.T.; Pawson, T.; Fleming, S.B.; Mercer, A.A. Poxvirus ankyrin repeat proteins are a unique class of F-box proteins that associate with cellular SCF1 ubiquitin ligase complexes. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 10955–10960. [Google Scholar] [CrossRef]
- Meng, X.; Xiang, Y. Vaccinia virus K1L protein supports viral replication in human and rabbit cells through a cell-type-specific set of its ankyrin repeat residues that are distinct from its binding site for ACAP2. Virology 2006, 353, 220–233. [Google Scholar] [CrossRef]
- Shisler, J.L.; Jin, X.L. The vaccinia virus K1L gene product inhibits host NF-kappaB activation by preventing IkappaBalpha degradation. J. Virol. 2004, 78, 3553–3560. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.B.; Wang, G.; Barrett, J.W.; Nazarian, S.H.; Colwill, K.; Moran, M.; McFadden, G. Myxoma virus M-T5 protects infected cells from the stress of cell cycle arrest through its interaction with host cell cullin-1. J. Virol. 2005, 79, 10750–10763. [Google Scholar] [CrossRef] [PubMed]
- Werden, S.J.; Lanchbury, J.; Shattuck, D.; Neff, C.; Dufford, M.; McFadden, G. The myxoma virus m-t5 ankyrin repeat host range protein is a novel adaptor that coordinately links the cellular signaling pathways mediated by Akt and Skp1 in virus-infected cells. J. Virol. 2009, 83, 12068–12083. [Google Scholar] [CrossRef]
- Blanie, S.; Mortier, J.; Delverdier, M.; Bertagnoli, S.; Camus-Bouclainville, C. M148R and M149R are two virulence factors for myxoma virus pathogenesis in the European rabbit. Vet. Res. 2009, 40, 11. [Google Scholar] [CrossRef]
- Mossman, K.; Lee, S.F.; Barry, M.; Boshkov, L.; McFadden, G. Disruption of M-T5, a novel myxoma virus gene member of poxvirus host range superfamily, results in dramatic attenuation of myxomatosis in infected European rabbits. J. Virol. 1996, 70, 4394–4410. [Google Scholar] [CrossRef]
- Camus-Bouclainville, C.; Fiette, L.; Bouchiha, S.; Pignolet, B.; Counor, D.; Filipe, C.; Gelfi, J.; Messud-Petit, F. A virulence factor of myxoma virus colocalizes with NF-kappaB in the nucleus and interferes with inflammation. J. Virol. 2004, 78, 2510–2516. [Google Scholar] [CrossRef]
- Sperling, K.M.; Schwantes, A.; Schnierle, B.S.; Sutter, G. The highly conserved orthopoxvirus 68k ankyrin-like protein is part of a cellular SCF ubiquitin ligase complex. Virology 2008, 374, 234–239. [Google Scholar] [CrossRef]
- Hsiao, J.C.; Chao, C.C.; Young, M.J.; Chang, Y.T.; Cho, E.C.; Chang, W. A poxvirus host range protein, CP77, binds to a cellular protein, HMG20A, and regulates its dissociation from the vaccinia virus genome in CHO-K1 cells. J. Virol. 2006, 80, 7714–7728. [Google Scholar] [CrossRef]
- Mohamed, M.R.; Rahman, M.M.; Lanchbury, J.S.; Shattuck, D.; Neff, C.; Dufford, M.; Van Buuren, N.; Fagan, K.; Barry, M.; Smith, S.; Damon, I.; McFadden, G. Proteomic screening of variola virus reveals a unique NF-kappaB inhibitor that is highly conserved among pathogenic orthopoxviruses. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 9045–9050. [Google Scholar] [CrossRef]
- Mohamed, M.R.; Rahman, M.M.; Rice, A.; Moyer, R.W.; Werden, S.J.; McFadden, G. Cowpox virus expresses a novel ankyrin repeat NF-kappaB inhibitor that controls inflammatory cell influx into virus-infected tissues and is critical for virus pathogenesis. J. Virol. 2009, 83, 9223–9236. [Google Scholar] [CrossRef]
- Peters, J.M. The anaphase promoting complex/cyclosome: A machine designed to destroy. Nat. Rev. Mol. Cell. Biol. 2006, 7, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, B.; Bharadwaj, R.; Zhu, H.; Ozkan, E.; Hakala, K.; Deisenhofer, J.; Yu, H. APC2 Cullin protein and APC11 RING protein comprise the minimal ubiquitin ligase module of the anaphase-promoting complex. Mol. Biol. Cell. 2001, 12, 3839–3851. [Google Scholar] [CrossRef] [PubMed]
- Leverson, J.D.; Joazeiro, C.A.; Page, A.M.; Huang, H.; Hieter, P.; Hunter, T. The APC11 RING-H2 finger mediates E2-dependent ubiquitination. Mol. Biol. Cell. 2000, 11, 2315–2325. [Google Scholar] [CrossRef] [PubMed]
- Glotzer, M.; Murray, A.W.; Kirschner, M.W. Cyclin is degraded by the ubiquitin pathway. Nature 1991, 349, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Pfleger, C.M.; Kirschner, M.W. The KEN box: An APC recognition signal distinct from the D box targeted by Cdh1. Genes Dev. 2000, 14, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Nasmyth, K. Disseminating the genome: Joining, resolving, and separating sister chromatids during mitosis and meiosis. Annu. Rev. Genet. 2001, 35, 673–745. [Google Scholar] [CrossRef]
- Mo, M.; Fleming, S.B.; Mercer, A.A. Cell cycle deregulation by a poxvirus partial mimic of anaphase-promoting complex subunit 11. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 19527–19532. [Google Scholar] [CrossRef]
- Johnston, J.B.; McFadden, G. Poxvirus immunomodulatory strategies: current perspectives. J. Virol. 2003, 77, 6093–6100. [Google Scholar] [CrossRef]
- Mimnaugh, E.G.; Chen, H.Y.; Davie, J.R.; Celis, J.E.; Neckers, L. Rapid deubiquitination of nucleosomal histones in human tumor cells caused by proteasome inhibitors and stress response inducers: effects on replication, transcription, translation, and the cellular stress response. Biochemistry 1997, 36, 14418–14429. [Google Scholar] [CrossRef]
- Lee, D.H.; Goldberg, A.L. Proteasome inhibitors: Valuable new tools for cell biologists. Trends Cell Biol. 1998, 8, 397–403. [Google Scholar] [CrossRef]
- Myung, J.; Kim, K.B.; Crews, C.M. The ubiquitin-proteasome pathway and proteasome inhibitors. Med. Res. Rev. 2001, 21, 245–273. [Google Scholar] [CrossRef]
- Einsele, H. Bortezomib. Recent Results Cancer Res. 2010, 184, 173–187. [Google Scholar]
- De Silva, F.S.; Moss, B. Origin-independent plasmid replication occurs in vaccinia virus cytoplasmic factories and requires all five known poxvirus replication factors. Virol. J. 2005, 2, 23. [Google Scholar] [CrossRef]
- Schubert, U.; Ott, D.E.; Chertova, E.N.; Welker, R.; Tessmer, U.; Princiotta, M.F.; Bennink, J.R.; Krausslich, H.G.; Yewdell, J.W. Proteasome inhibition interferes with gag polyprotein processing, release, and maturation of HIV-1 and HIV-2. Proc. Natl. Acad. Sci. U. S. A. 2000, 97, 13057–13062. [Google Scholar] [CrossRef]
- Dudek, S.E.; Luig, C.; Pauli, E.K.; Schubert, U.; Ludwig, S. The clinically approved proteasome inhibitor PS-341 efficiently blocks influenza A virus and vesicular stomatitis virus propagation by establishing an antiviral state. J. Virol. 2010, 84, 9439–9451. [Google Scholar] [CrossRef]
- Raaben, M.; Posthuma, C.C.; Verheije, M.H.; Te Lintelo, E.G.; Kikkert, M.; Drijfhout, J.W.; Snijder, E.J.; Rottier, P.J.; de Haan, C.A. The ubiquitin-proteasome system plays an important role during various stages of the coronavirus infection cycle. J. Virol. 2010, 84, 7869–7879. [Google Scholar] [CrossRef]
- Prosch, S.; Priemer, C.; Hoflich, C.; Liebenthaf, C.; Babel, N.; Kruger, D.H.; Volk, H.D. Proteasome inhibitors: A novel tool to suppress human cytomegalovirus replication and virus-induced immune modulation. Antivir. Ther. 2003, 8, 555–567. [Google Scholar] [CrossRef]
- Lupfer, C.; Pastey, M.K. Decreased replication of human respiratory syncytial virus treated with the proteasome inhibitor MG-132. Virus Res. 2010, 149, 36–41. [Google Scholar] [CrossRef]
- Delboy, M.G.; Roller, D.G.; Nicola, A.V. Cellular proteasome activity facilitates herpes simplex virus entry at a postpenetration step. J. Virol. 2008, 82, 3381–3390. [Google Scholar] [CrossRef]
- Bandi, P.; Garcia, M.L.; Booth, C.J.; Chisari, F.V.; Robek, M.D. Bortezomib inhibits hepatitis B virus replication in transgenic mice. Antimicrob. Agents Chemother. 2010, 54, 749–756. [Google Scholar] [CrossRef]
- Basler, M.; Lauer, C.; Beck, U.; Groettrup, M. The proteasome inhibitor bortezomib enhances the susceptibility to viral infection. J. Immunol. 2009, 183, 6145–6150. [Google Scholar] [CrossRef] [PubMed]
- Fagan-Garcia, K. University of Alberta, Edmonton, Canada. Unpublished work. 2010. [Google Scholar]
- Han, X.; Aslanian, A.; Yates, J.R., 3rd. Mass spectrometry for proteomics. Curr. Opin. Chem. Biol. 2008, 12, 483–490. [Google Scholar] [CrossRef]
- Kirkpatrick, D.S.; Denison, C.; Gygi, S.P. Weighing in on ubiquitin: The expanding role of mass-spectrometry-based proteomics. Nat. Cell Biol. 2005, 7, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Rigbolt, K.T.; Blagoev, B. Proteome-wide quantitation by SILAC. Methods Mol. Biol. 2010, 658, 187–204. [Google Scholar]
- Yates, J.R.; Ruse, C.I.; Nakorchevsky, A. Proteomics by mass spectrometry: Approaches, advances, and applications. Annu. Rev. Biomed. Eng. 2009, 11, 49–79. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/). |




{kind=link}
{kind=link}
{kind=link}
| Involvement in Ubiquitination | Genus | Virusa | Gene/Protein | Length, aa | VBRC accessionb |
|---|---|---|---|---|---|
| poxvirus-encoded ubiquitin homologs | Avipoxvirus | CNPV-VR111 | 96 | 85 | VP0043569 |
| Betaentomopoxvirus | AMEV-Moyer | 180 | 81 | VP0037620 | |
| Unclassified Poxviridae | MSEV-Tuc | 144 | 80 | VP0038302 | |
| MARCH Poxviral E3 Ubiquitin Ligase | Capripoxvirus | GTPV-Pellor | 8 | 162 | VP0044818 |
| LSDV-Nee | 10 | 162 | VP0040213 | ||
| SPPV-A | 8 | 162 | VP0044517 | ||
| Leporipoxvirus | MYXV-Lau | M153 | 206 | VP0038581 | |
| RFV-Kas | gp153R | 201 | VP0038747 | ||
| Suipoxvirus | SWPV-Neb | 9 | 155 | VP0040564 | |
| Yatapoxvirus | TANV-COD | 5 | 156 | VP0067544 | |
| YLDV-Davis | 5 | 156 | VP0040054 | ||
| YMTV-Amano | 4 | 156 | VP0043053 | ||
| p28 Poxviral E3 Ubiquitin Ligase | Avipoxvirus | CNPV-VR111 | 205 | 318 | VP0043678 |
| 197 | 275 | VP0043670 | |||
| FWPV-Iowa | 157 | 311 | VP0037889 | ||
| 150 | 276 | VP0037882 | |||
| Capripoxvirus | LSDV-Nee | 140 | 240 | VP0040345 | |
| SPPV-A | 136 | 240 | VP0044645 | ||
| GTPV-Pellor | 127 | 240 | VP0044947 | ||
| Leporipoxvirus | MYXV-Lau | M143 | 234 | VP0038572 | |
| RFV-Kas | gp143R (N1R) | 234 | VP0038740 | ||
| Orthopoxvirus | CMLV-CMS | 14R | 242 | VP0041112 | |
| CPXV-GRI | C7R | 242 | VP0042678 | ||
| ECTV-Mos | 12 | 241 | VP0040932 | ||
| MPXV-ZAR | D5R | 242 | VP0040369 | ||
| VACV IHD-W | p28 | 243 | c | ||
| VARV-BGD75maj | D6Rd | 242 | VP0038767 | ||
| RPXV-Utr | 8 | 242 | VP0041370 | ||
| Suipoxvirus | SWPV-Neb | 138 | 246 | VP0040694 | |
| Yatapoxvirus | TANV-COD | 143R | 234 | VP0067759 | |
| YMTV-Amano | 143R | 236 | VP0043181 | ||
| Unclassified Poxviridae | DPV-W1170_84 | 154 | 245 | VP0045437 | |
| BTB/KELCH proteins associated with cullin-3-based E3 ubiquitin ligase | Capripoxvirus | GTPV-Pellor | 16 | 562 | VP0044826 |
| 141 | 547 | VP0044951 | |||
| 148 | 552 | VP0044958 | |||
| LSDV-NEE | 19 | 569 | VP0040222 | ||
| 144 | 547 | VP0040349 | |||
| 151 | 550 | VP0040356 | |||
| SPPV-A | 16 | 569 | VP0044525 | ||
| 140 | 547 | VP0044649 | |||
| 147 | 552 | VP0044656 | |||
| Leporipoxvirus | MYXV-Lau | M014L | 517 | VP0038442 | |
| M140R | 553 | VP0038569 | |||
| RFV-Kas | gp013L | 516 | VP0038613 | ||
| gp0140R | 553 | VP0038737 | |||
| Orthopoxvirus | CMLV-CMS | 21L | 200 | VP0041119 | |
| 24L | 512 | VP0041122 | |||
| 38L | 480 | VP0041137 | |||
| 172R | 564 | VP0041317 | |||
| 186R | 501 | VP0041335 | |||
| CPXV-GRI | D11L | 521 | VP0042668 | ||
| C18L | 512 | VP0042689 | |||
| G3L | 485 | VP0042703 | |||
| A54R | 564 | VP0042838 | |||
| B9R | 501 | VP0042849 | |||
| B19R | 557 | VP0042686 | |||
| ECTV-Mos | 18 | 512 | VP0040938 | ||
| 27 | 482 | VP0040947 | |||
| 150 | 563 | VP0041074 | |||
| 165 | 594 | VP0041089 | |||
| MPXV-ZAI | D12L | 206 | VP0040376 | ||
| D19L | 107 | VP0040382 | |||
| C9L | 487 | VP0040396 | |||
| TATV-DAH68 | 24 | 150 | VP0052942 | ||
| 43 | 480 | VP0052961 | |||
| 181 | 219 | VP0053099 | |||
| 196 | 209 | VP0053114 | |||
| VACV-COP | C2L | 512 | VP0039555 | ||
| C5L | 615 | VP0039551 | |||
| F3L | 480 | VP0039572 | |||
| A55R | 564 | VP0039751 | |||
| RPXV-Utr | 15 | 204 | VP0041377 | ||
| 18 | 512 | VP0041380 | |||
| 31 | 480 | VP0041393 | |||
| 162 | 564 | VP0041526 | |||
| Suipoxvirus | SWPV-Neb | 6 | 530 | VP0040561 | |
| 15 | 534 | VP0040570 | |||
| 136 | 574 | VP0040692 | |||
| Yatapoxvirus | YLDV-Davis | 19L | 522 | VP0040068 | |
| 140R | 570 | VP0040192 | |||
| YMTV-Amano | 19L | 524 | VP0043062 | ||
| Unclassified Poxviridae | DPV-W1170_84 | 25 | 529 | VP0045308 | |
| 159 | 546 | VP0045442 | |||
| ankyrin/PRANC proteins associated with cullin-1-based E3 ubiquitin ligase | Avipoxvirus | FWPV-Iowa | 12 | 331 | VP0037744 |
| 14 | 437 | VP0037746 | |||
| 18 | 700 | VP0037750 | |||
| 22 | 578 | VP0037754 | |||
| 26 | 436 | VP0037758 | |||
| 31 | 341 | VP0037763 | |||
| 162 | 603 | VP0037894 | |||
| 218 | 461 | VP0037952 | |||
| 219 | 434 | VP0037953 | |||
| 222 | 747 | VP0037956 | |||
| 227 | 361 | VP0037961 | |||
| 228 | 525 | VP0037962 | |||
| 231 | 256 | VP0037965 | |||
| 232 | 482 | VP0037966 | |||
| 233 | 512 | VP0037967 | |||
| 234 | 428 | VP0037968 | |||
| 240 | 410 | VP0037974 | |||
| 243 | 262 | VP0037977 | |||
| 244 | 668 | VP0037978 | |||
| 246 | 592 | VP0037980 | |||
| Capripoxvirus | GTPV-Pellor | 142 | 634 | VP0044952 | |
| 144 | 498 | VP0044954 | |||
| 145 | 447 | VP0044955 | |||
| 149 | 453 | VP0044959 | |||
| LSDV-Nee | 145 | 634 | VP0040350 | ||
| 147 | 498 | VP0040352 | |||
| 148 | 447 | VP0040353 | |||
| 152 | 489 | VP0042090 | |||
| SPPV-A | 141 | 631 | VP0044650 | ||
| 143 | 498 | VP0044652 | |||
| 144 | 447 | VP0044653 | |||
| 148 | 484 | VP0044657 | |||
| Leporipoxvirus | MYXV-Lau | 148R | 675 | VP0038576 | |
| 149R | 490 | VP0038577 | |||
| 150R | 494 | VP0038578 | |||
| 005R (MT-5) | 483 | VP0038588 | |||
| Orthopoxvirus | CMLV-CMS | 3L | 585 | VP0041099 | |
| 4L | 672 | VP0041101 | |||
| 177L | 564 | VP0041325 | |||
| 197R | 783 | VP0041349 | |||
| CPXV-GRI | D3L | 586 | VP0042660 | ||
| D4L | 672 | VP0042661 | |||
| D8L (CP77) | 661 | VP0042665 | |||
| C1L | 437 | VP0042672 | |||
| C11L | 614 | VP0042682 | |||
| B3R | 558 | VP0042843 | |||
| B16R | 574 | VP0042856 | |||
| B18R | 795 | VP0042858 | |||
| K1R | 581 | VP0042863 | |||
| I2R | 672 | VP0042868 | |||
| I3R | 586 | VP0042869 | |||
| ECTV-Mos | 2 | 587 | VP0040921 | ||
| 5 | 650 | VP0040924 | |||
| 154 | 564 | VP0041078 | |||
| 165 | 594 | VP0041089 | |||
| MPXV-ZAR | B5R | 561 | VP0040530 | ||
| J1R | 587 | VP0040553 | |||
| N4R | 437 | VP0040552 | |||
| B17R | 793 | VP0040542 | |||
| TATV-DAH68 | 220 | 640 | VP0053138 | ||
| 187 | 558 | VP0053105 | |||
| 18 | 661 | VP0052936 | |||
| 6 | 627 | VP0052924 | |||
| VACV-Cop | B18R | 574 | VP0039778 | ||
| C19L | 259 | VP0039532 | |||
| B4R | 558 | VP0039761 | |||
| RPXV-Utr | 180 | 791 | VP0041544 | ||
| 178 | 574 | VP0041542 | |||
| 166 | 558 | VP0041530 | |||
| VARV-BDG75maj | B5R | 558 | VP0038933 | ||
| G1R | 585 | VP0039159 | |||
| B16R | 574 | VP0038944 | |||
| B18R | 787 | VP0038946 | |||
| Parapoxvirus | ORFV-NZ2 | 8 | 516 | VP0047660 | |
| 123 | 525 | VP0047777 | |||
| 126 | 497 | VP0047780 | |||
| 128 | 500 | VP0047782 | |||
| 129 | 520 | VP0047783 | |||
| Suipoxvirus | SWPV-Neb | 141 | 635 | VP0040697 | |
| 142 | 485 | VP0040698 | |||
| 143 | 430 | VP0040699 | |||
| 144 | 493 | VP0040700 | |||
| Yatapoxvirus | YLDV-Davis | 148R | 476 | VP0040200 | |
| 147R | 491 | VP0040199 | |||
| 146R | 473 | VP0040198 | |||
| 11L | 637 | VP0040060 | |||
| YMTV-Amano | 11L | 637 | VP0043056 | ||
| 146R | 356 | VP0043184 | |||
| 147R | 497 | VP0043185 | |||
| 148R | 483 | VP0043186 | |||
| Unclassified Poxviridae | DPV-W1170_84 | 164 | 493 | VP0045447 | |
| 163 | 483 | VP0045446 | |||
| 162 | 501 | VP0045445 | |||
| 160 | 641 | VP0045443 | |||
| 19 | 643 | VP0045302 | |||
| poxvirus APC/cyclosome regulators | Molluscipoxvirus | MOCV-st1 | 026L | 83 | VP0038021 |
| Parapoxvirus | BSPV-AR02 | 13 | 93 | VP0043354 | |
| ORFV-NZ2 | 14 | 93 | VP0047667 | ||
| Unclassified Poxvirdae | CRV-ZWE | 47 | 81 | VP0066074 | |
| SPV | A11L | 86 | DQ377804e |
© 2010 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Barry, M.; Van Buuren, N.; Burles, K.; Mottet, K.; Wang, Q.; Teale, A. Poxvirus Exploitation of the Ubiquitin-Proteasome System. Viruses 2010, 2, 2356-2380. https://doi.org/10.3390/v2102356
Barry M, Van Buuren N, Burles K, Mottet K, Wang Q, Teale A. Poxvirus Exploitation of the Ubiquitin-Proteasome System. Viruses. 2010; 2(10):2356-2380. https://doi.org/10.3390/v2102356
Chicago/Turabian StyleBarry, Michele, Nicholas Van Buuren, Kristin Burles, Kelly Mottet, Qian Wang, and Alastair Teale. 2010. "Poxvirus Exploitation of the Ubiquitin-Proteasome System" Viruses 2, no. 10: 2356-2380. https://doi.org/10.3390/v2102356
APA StyleBarry, M., Van Buuren, N., Burles, K., Mottet, K., Wang, Q., & Teale, A. (2010). Poxvirus Exploitation of the Ubiquitin-Proteasome System. Viruses, 2(10), 2356-2380. https://doi.org/10.3390/v2102356