Antiviral Immunity in Amphibians


Abstract
:1. Introduction
2. Organization of the Ectothermic Vertebrate Immune System Compared to Mammals
3. The Xenopus Immune System
4. Immunity to RVs in Xenopus and Other Anuran Species
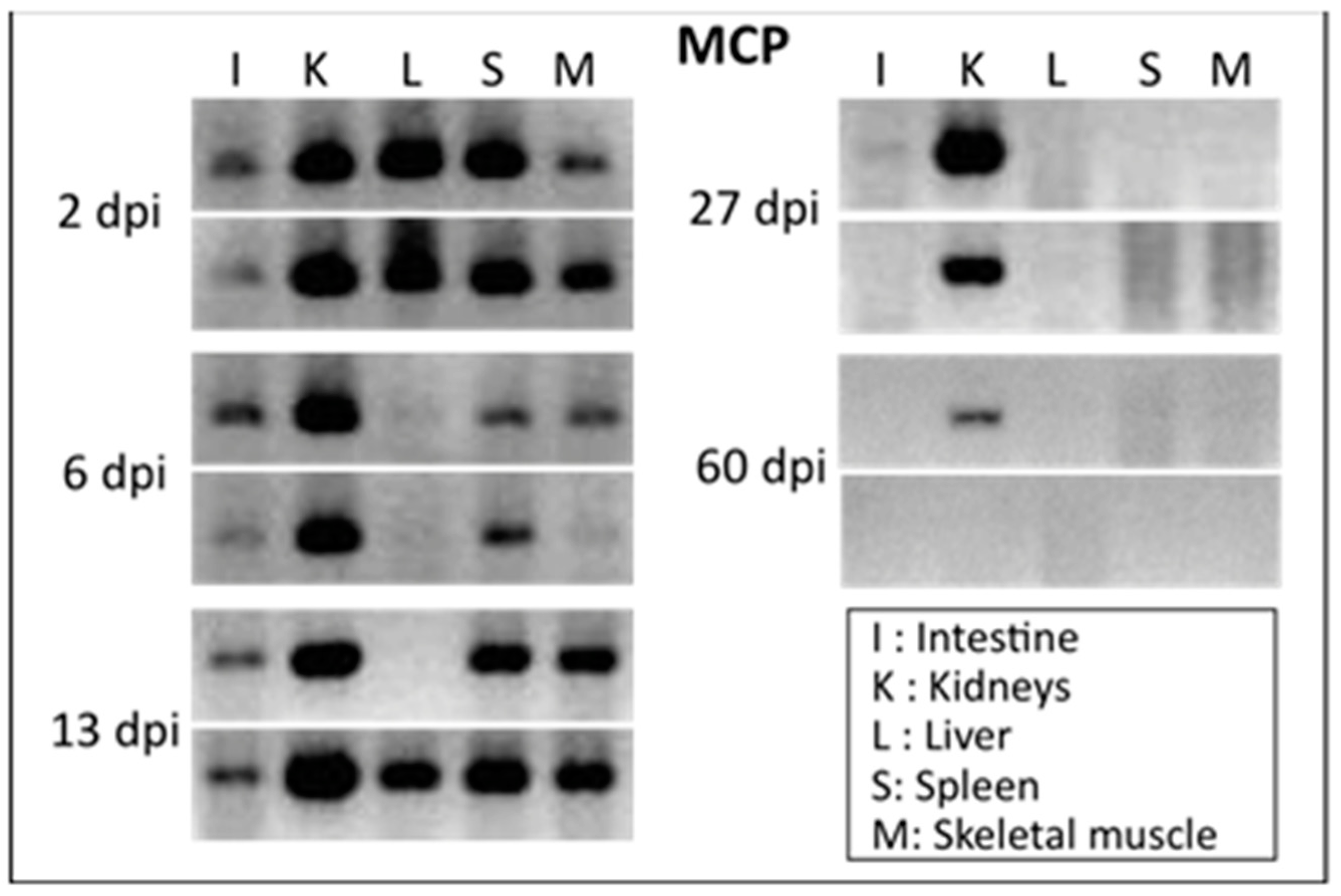
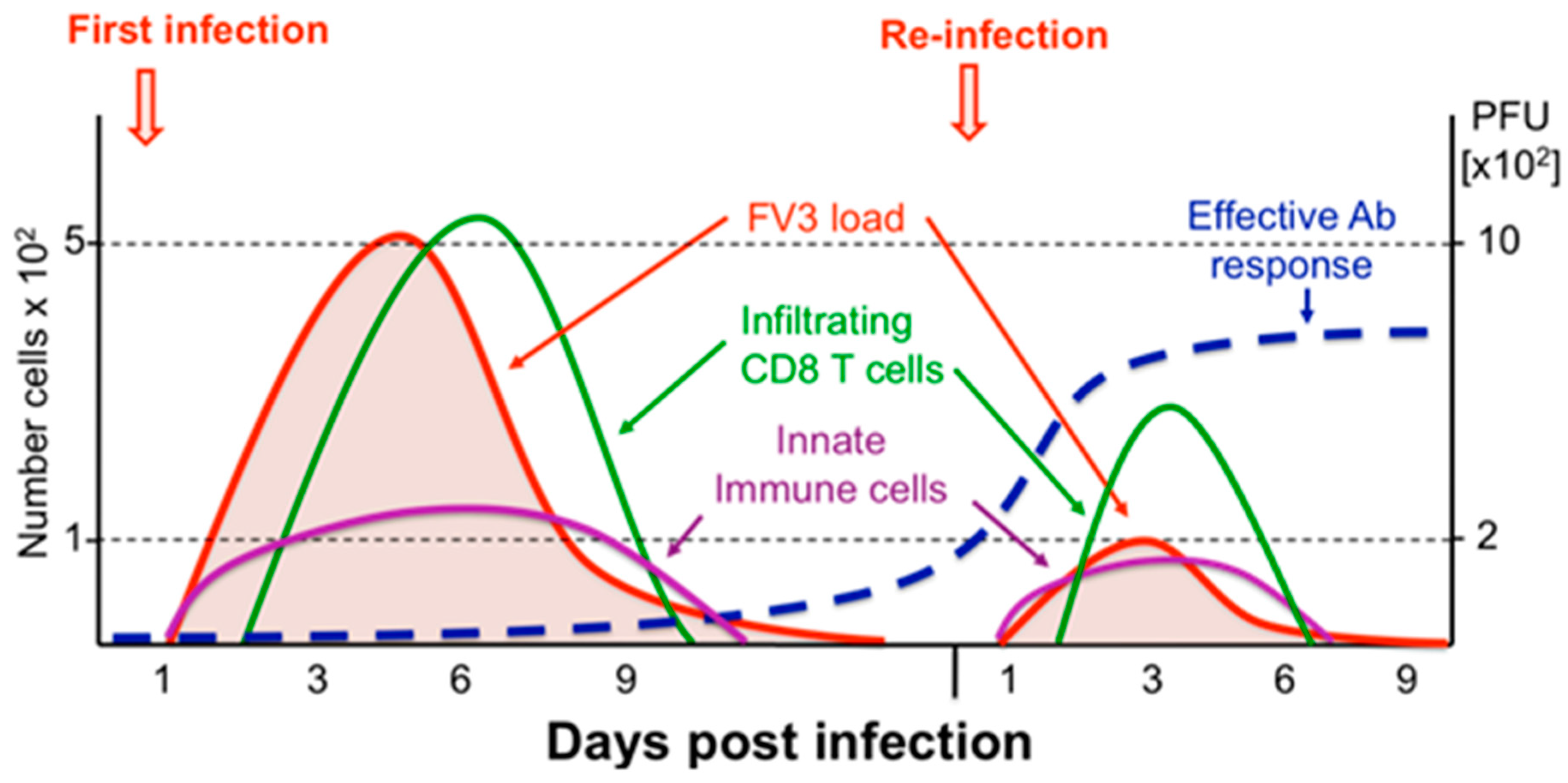
4.1. Adults
4.2. Larvae
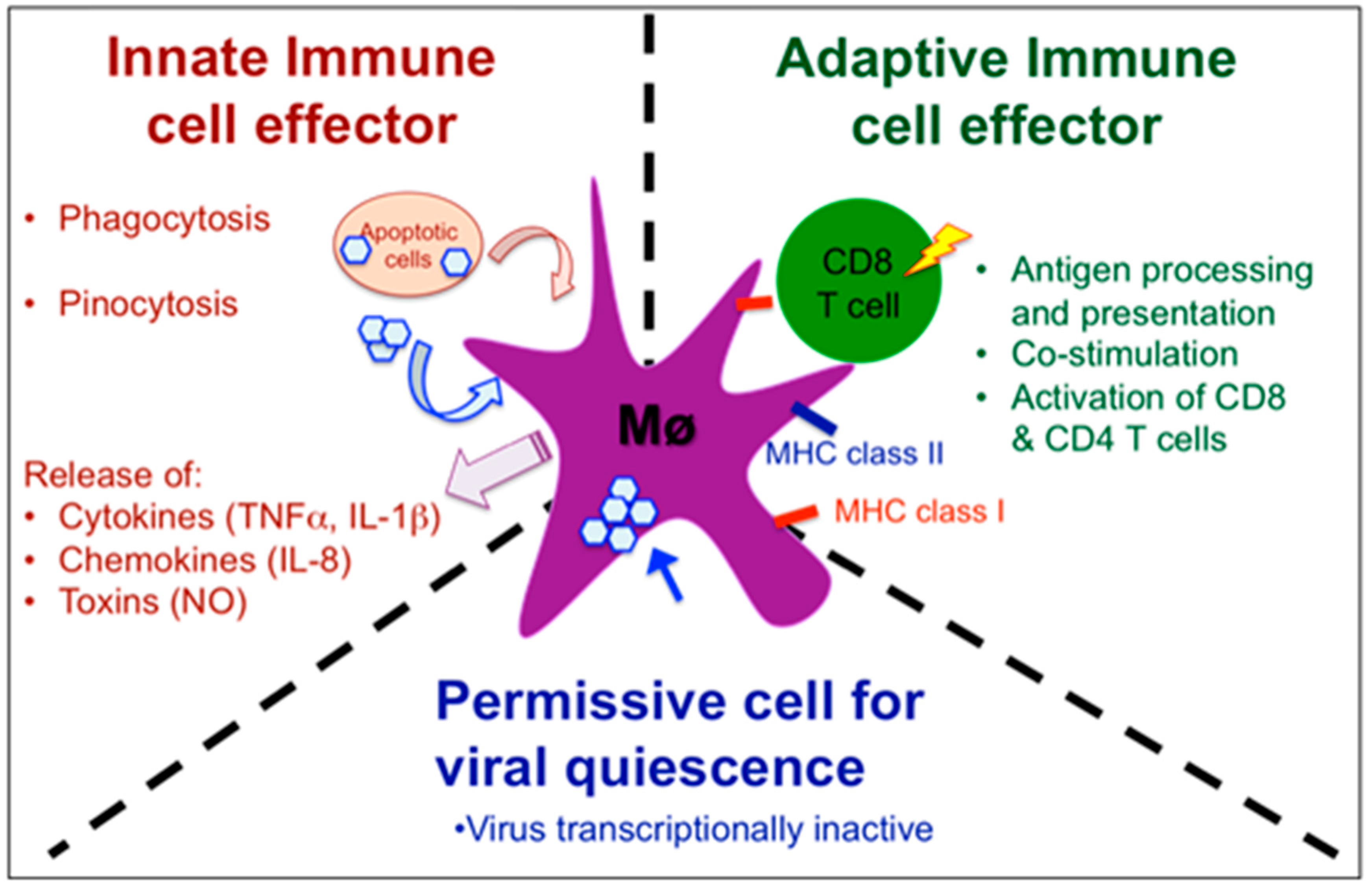
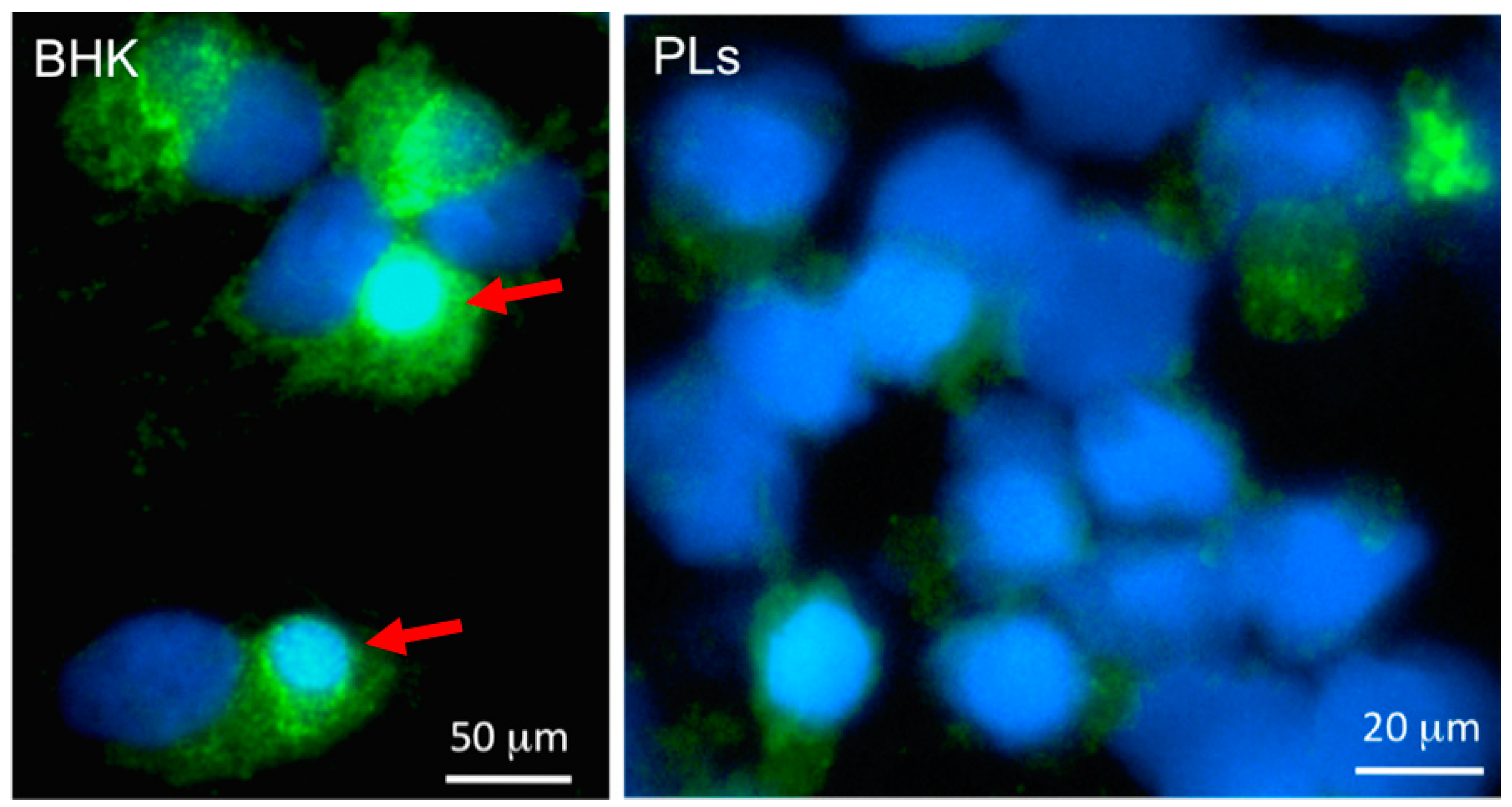
4.3. Complex Role of Xenopus Macrophages in Host Defenses and Viral Persistence
4.4. MHC Genotype and Susceptibility to RVs
5. The Immunity to RVs in Salamanders
6. Viral Immune Evasion and Virulence Proteins
7. Role of Host Immune System in Pathogenesis Transmission and Persistence of RV Infections
8. Concluding Remarks
Acknowledgments
Conflicts of Interest
References and Notes
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- The International Union for Conservation of Nature and Natural Resources (IUCN) Red List of Threatened Species (Amphibians). Available online: http://www.iucnredlist.org/initiatives/amphibians (accessed on 28 October 2011).
- Daszak, P.; Berger, L.; Cunningham, A.A.; Hyatt, A.D.; Green, D.E.; Speare, R. Emerging infectious diseases and amphibian population declines. Emerg. Infect. Dis. 1999, 5, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.P. Amphibian decline and extinction: What we know and what we need to learn. Dis. Aquat. Organ. 2010, 92, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Briggs, C.J.; Daszak, P. The ecology and impact of chytridiomycosis: An emerging disease of amphibians. Trends Ecol. Evol. 2010, 25, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Green, D.E.; Converse, K.A.; Schrader, A.K. Epizootiology of sixty-four amphibian morbidity and mortality events in the USA, 1996–2001. Ann. N. Y. Acad. Sci. 2002, 969, 323–339. [Google Scholar] [CrossRef]
- Robert, J.; Gregory Chinchar, V. "Ranaviruses: An emerging threat to ectothermic vertebrates" report of the first international symposium on ranaviruses, Minneapolis MN July 8, 2011. Dev. Comp. Immunol. 2011. [Google Scholar] [CrossRef]
- Lesbarrères, D.; Balseiro, A.; Brunner, J.; Chinchar, V.G.; Duffus, A.; Kerby, J.; Miller, D.L.; Robert, J.; Schock, D.M.; Waltzek, T.; et al. Symposium report:Ranavirus: Past, present and future. Biol. Lett. 2011, in press. [Google Scholar]
- Kerby, J. Global focus: 1st international symposium on Ranaviruses. FrogLog 2011, 98, 33–35. [Google Scholar]
- 2011 International Ranavirus Symposium. Available online: http://Fwf.Ag.Utk.Edu/Mgray/Ranavirus/2011ranavirus.htm (accessed on 28 October 2011).
- Global Ranavirus Consortium. Available online: http://fwf.Ag.Utk.Edu/mgray/ranavirus/ranavirus.htm (accessed on 28 October 2011).
- The World Organisation for Animal Health. Available online: http://www.Oie.Int/ (accessed on 28 October 2011).
- Schloegel, L.M.; Daszak, P.; Cunningham, A.A.; Speare, R.; Hill, B. Two amphibian diseases, chytridiomycosis and ranaviral disease, are now globally notifiable to the world organization for animal health (oie): An assessment. Dis. Aquat. Organ. 2010, 92, 101–108. [Google Scholar] [CrossRef]
- Gray, M.J.; Miller, D.L.; Hoverman, J.T. Ecology and pathology of amphibian ranaviruses. Dis. Aquat. Organ. 2009, 87, 243–266. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Hyatt, A.; Miyazaki, T.; Williams, T. Family iridoviridae: Poor viral relations no longer. Curr. Top. Microbiol. Immunol. 2009, 328, 123–170. [Google Scholar] [PubMed]
- Chinchar, V.G. Ranaviruses (family iridoviridae): Emerging cold-blooded killers. Arch. Virol. 2002, 147, 447–470. [Google Scholar] [CrossRef]
- Carey, C.; Cohen, N.; Rollins-Smith, L. Amphibian declines: An immunological perspective. Dev. Comp. Immunol. 1999, 23, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Hayes, T.B.; Falso, P.; Gallipeau, S.; Stice, M. The cause of global amphibian declines: A developmental endocrinologist’s perspective. J. Exp. Biol. 2010, 213, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Workenhe, S.T.; Rise, M.L.; Kibenge, M.J.; Kibenge, F.S. The fight between the teleost fish immune response and aquatic viruses. Mol. Immunol. 2010, 47, 2525–2536. [Google Scholar] [CrossRef]
- Robert, J.; Ohta, Y. Comparative and developmental study of the immune system in xenopus. Dev. Dyn. 2009, 238, 1249–1270. [Google Scholar] [CrossRef]
- Goodbourn, S.; Didcock, L.; Randall, R.E. Interferons: Cell signalling, immune modulation, antiviral response and virus countermeasures. J. Gen. Virol. 2000, 81, 2341–2364. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr. The immune system evolved to discriminate infectious nonself from noninfectious self. Immunol. Today 1992, 13, 11–16. [Google Scholar] [CrossRef]
- Lee, S.H.; Miyagi, T.; Biron, C.A. Keeping nk cells in highly regulated antiviral warfare. Trends Immunol. 2007, 28, 252–259. [Google Scholar] [CrossRef]
- Prince, L.R.; Whyte, M.K.; Sabroe, I.; Parker, L.C. The role of tlrs in neutrophil activation. Curr. Opin. Pharmacol. 2011, 11, 397–403. [Google Scholar] [CrossRef]
- Viret, C.; Janeway, C.A., Jr. Mhc and t cell development. Rev. Immunogenet. 1999, 1, 91–104. [Google Scholar] [PubMed]
- Du Pasquier, L.; Schwager, J.; Flajnik, M.F. The immune system of xenopus. Annu. Rev. Immunol. 1989, 7, 251–275. [Google Scholar] [CrossRef]
- Marr, S.; Morales, H.; Bottaro, A.; Cooper, M.; Flajnik, M.; Robert, J. Localization and differential expression of activation-induced cytidine deaminase in the amphibian xenopus upon antigen stimulation and during early development. J. Immunol. 2007, 179, 6783–6789. [Google Scholar] [CrossRef]
- Li, J.; Barreda, D.R.; Zhang, Y.A.; Boshra, H.; Gelman, A.E.; Lapatra, S.; Tort, L.; Sunyer, J.O. B lymphocytes from early vertebrates have potent phagocytic and microbicidal abilities. Nat. Immunol. 2006, 7, 1116–1124. [Google Scholar] [CrossRef] [PubMed]
- Ishii, A.; Kawasaki, M.; Matsumoto, M.; Tochinai, S.; Seya, T. Phylogenetic and expression analysis of amphibian xenopus toll-like receptors. Immunogenetics 2007, 59, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Roach, J.C.; Glusman, G.; Rowen, L.; Kaur, A.; Purcell, M.K.; Smith, K.D.; Hood, L.E.; Aderem, A. The evolution of vertebrate toll-like receptors. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 9577–9582. [Google Scholar] [CrossRef]
- Bleicher, P.A.; Rollins-Smith, L.A.; Jacobs, D.M.; Cohen, N. Mitogenic responses of frog lymphocytes to crude and purified preparations of bacterial lipopolysaccharide (lps). Dev. Comp. Immunol. 1983, 7, 483–496. [Google Scholar] [CrossRef]
- Mescher, A.L.; Wolf, W.L.; Moseman, E.A.; Hartman, B.; Harrison, C.; Nguyen, E.; Neff, A.W. Cells of cutaneous immunity in xenopus: Studies during larval development and limb regeneration. Dev. Comp. Immunol. 2007, 31, 383–393. [Google Scholar] [CrossRef]
- Morales, H.D.; Abramowitz, L.; Gertz, J.; Sowa, J.; Vogel, A.; Robert, J. Innate immune responses and permissiveness to ranavirus infection of peritoneal leukocytes in the frog xenopus laevis. J. Virol. 2010, 84, 4912–4922. [Google Scholar] [CrossRef]
- Du Pasquier, L.; Robert, J.; Courtet, M.; Mussmann, R. B-cell development in the amphibian xenopus. Immunol. Rev. 2000, 175, 201–213. [Google Scholar] [CrossRef]
- Hsu, E. Mutation, selection, and memory in b lymphocytes of exothermic vertebrates. Immunol. Rev. 1998, 162, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.; Hsu, E.; Marcuz, A.; Courtet, M.; Du Pasquier, L.; Steinberg, C. What limits affinity maturation of antibodies in xenopus—The rate of somatic mutation or the ability to select mutants? EMBO J. 1992, 11, 4337–4347. [Google Scholar] [CrossRef]
- Flajnik, M.F.; Du Pasquier, L. Mhc class i antigens as surface markers of adult erythrocytes during the metamorphosis of xenopus. Dev. Biol. 1988, 128, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Horton, T.L.; Stewart, R.; Cohen, N.; Rau, L.; Ritchie, P.; Watson, M.D.; Robert, J.; Horton, J.D. Ontogeny of xenopus nk cells in the absence of mhc class i antigens. Dev. Comp. Immunol. 2003, 27, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Gantress, J.; Maniero, G.D.; Cohen, N.; Robert, J. Development and characterization of a model system to study amphibian immune responses to iridoviruses. Virology 2003, 311, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Robert, J. Comparative study of infectivity of frog virus 3, rana catesbeiana virus z, and ambystoma tigrinum virus in xenopus laevis and xenopus tropicalis. University of Rochester Medical Center, Rocherster, NY, USA. Unpublished work, 2011.
- Robert, J.; Morales, H.; Buck, W.; Cohen, N.; Marr, S.; Gantress, J. Adaptive immunity and histopathology in frog virus 3-infected xenopus. Virology 2005, 332, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Morales, H.D.; Robert, J. Characterization of primary and memory cd8 t-cell responses against ranavirus (fv3) in xenopus laevis. J. Virol. 2007, 81, 2240–2248. [Google Scholar] [CrossRef]
- Panchanathan, V.; Chaudhri, G.; Karupiah, G. Antiviral protection following immunization correlates with humoral but not cell-mediated immunity. Immunol. Cell Biol. 2010, 88, 461–467. [Google Scholar] [CrossRef]
- Maniero, G.D.; Morales, H.; Gantress, J.; Robert, J. Generation of a long-lasting, protective, and neutralizing antibody response to the ranavirus fv3 by the frog xenopus. Dev. Comp. Immunol. 2006, 30, 649–657. [Google Scholar] [CrossRef]
- Zupanovic, Z.; Lopez, G.; Hyatt, A.D.; Green, B.; Bartran, G.; Parkes, H.; Whittington, R.J.; Speare, R. Giant toads bufo marinus in australia and venezuela have antibodies against ’ranaviruses’. Dis. Aquat. Organ. 1998, 32, 1–8. [Google Scholar] [CrossRef]
- Majji, S.; LaPatra, S.; Long, S.M.; Sample, R.; Bryan, L.; Sinning, A.; Chinchar, V.G. Rana catesbeiana virus z (rcv-z): A novel pathogenic ranavirus. Dis. Aquat. Organ. 2006, 73, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.A.; Tems, C.A.; Russell, P.H. Immunohistochemical demonstration of ranavirus antigen in the tissues of infected frogs (rana temporaria) with systemic haemorrhagic or cutaneous ulcerative disease. J. Comp. Pathol. 2008, 138, 3–11. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Bryan, L.; Silphadaung, U.; Noga, E.; Wade, D.; Rollins-Smith, L. Inactivation of viruses infecting ectothermic animals by amphibian and piscine antimicrobial peptides. Virology 2004, 323, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G.; Wang, J.; Murti, G.; Carey, C.; Rollins-Smith, L. Inactivation of frog virus 3 and channel catfish virus by esculentin-2p and ranatuerin-2p, two antimicrobial peptides isolated from frog skin. Virology 2001, 288, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Tweedell, K.; Granoff, A. Viruses and renal carcinoma of rana pipiens. V. Effect of frog virus 3 on developing frog embryos and larvae. J. Natl. Cancer Inst. 1968, 40, 407–410. [Google Scholar] [PubMed]
- Miller, D.L.; Rajeev, S.; Gray, M.J.; Baldwin, C.A. Frog virus 3 infection, cultured american bullfrogs. Emerg. Infect. Dis. 2007, 13, 342–343. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, R.; de Mesquita, A.J.; Fleury, L.F.; de Brito, W.M.; Nunes, I.A.; Robert, J.; Morales, H.; Coelho, A.S.; Barthasson, D.L.; Galli, L.; et al. Mass mortality associated with a frog virus 3-like ranavirus infection in farmed tadpoles rana catesbeiana from brazil. Dis. Aquat. Organ. 2009, 86, 181–191. [Google Scholar] [CrossRef]
- Chen, G.; Robert, J. Susceptibility of xenopus laevis tadpoles to infection by the ranavirus frog-virus 3 correlates with a weak and delayed innate immune response. Eur. J. Immunol. submitted for publication, 2011.
- Nishikawa, A.; Murata, E.; Akita, M.; Kaneko, K.; Moriya, O.; Tomita, M.; Hayashi, H. Roles of macrophages in programmed cell death and remodeling of tail and body muscle of xenopus laevis during metamorphosis. Histochem. Cell Biol. 1998, 109, 11–17. [Google Scholar] [CrossRef]
- Whitley, D.S.; Yu, K.; Sample, R.C.; Sinning, A.; Henegar, J.; Norcross, E.; Chinchar, V.G. Frog virus 3 orf 53r, a putative myristoylated membrane protein, is essential for virus replication in vitro. Virology 2010, 405, 448–456. [Google Scholar] [CrossRef]
- Robert, J.; George, E.; De Jesus Andino, F.; Chen, G. Waterborne infectivity of the ranavirus frog virus 3 in xenopus laevis. Virology 2011, 417, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Haislip, N.A.; Gray, M.J.; Hoverman, J.T.; Miller, D.L. Development and disease: How susceptibility to an emerging pathogen changes through anuran development. PLoS One 2011, 6, e22307. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.J. Are ranaviruses capable of contributing to amphibian species declines? Available online: http://fwf.ag.utk.edu/mgray/ranavirus/2011Symposium/Gray.pdf (accessed on 28 October 2011).
- Flajnik, M.F.; Du Pasquier, L. The major histocompatibility complex of frogs. Immunol. Rev. 1990, 113, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Teacher, A.G.; Garner, T.W.; Nichols, R.A. Evidence for directional selection at a novel major histocompatibility class i marker in wild common frogs (rana temporaria) exposed to a viral pathogen (ranavirus). PLoS One 2009, 4, e4616. [Google Scholar] [CrossRef] [PubMed]
- Cotter, J.D.; Storfer, A.; Page, R.B.; Beachy, C.K.; Voss, S.R. Transcriptional response of mexican axolotls to ambystoma tigrinum virus (atv) infection. BMC Genomics 2008, 9, 493. [Google Scholar] [CrossRef] [PubMed]
- Manning, M.J. Histological organization of the spleen: Implications for immune functions in amphibians. Res. Immunol. 1991, 142, 355–359. [Google Scholar] [CrossRef]
- Ching, Y.C.; Wedgwood, R.J. Immunologic responses in the axolotl, siredon mexicanum. J. Immunol. 1967, 99, 191–200. [Google Scholar] [CrossRef]
- Charlemagne, J. Thymus independent anti-horse erythrocyte antibody response and suppressor t cells in the mexican axolotl (amphibia, urodela, ambystoma mexicanum). Immunology 1979, 36, 643–648. [Google Scholar]
- Kaufman, J.; Volk, H.; Wallny, H.J. A "minimal essential mhc" and an "unrecognized mhc": Two extremes in selection for polymorphism. Immunol. Rev. 1995, 143, 63–88. [Google Scholar] [CrossRef]
- Tournefier, A.; Laurens, V.; Chapusot, C.; Ducoroy, P.; Padros, M.R.; Salvadori, F.; Sammut, B. Structure of mhc class i and class ii cdnas and possible immunodeficiency linked to class ii expression in the mexican axolotl. Immunol. Rev. 1998, 166, 259–277. [Google Scholar] [CrossRef]
- Cohen, N. Amphibian transplantation reactions. Amer. Zool. 1971, 11, 93–205. [Google Scholar] [CrossRef]
- Rojas, S.; Richards, K.; Jancovich, J.K.; Davidson, E.W. Influence of temperature on ranavirus infection in larval salamanders ambystoma tigrinum. Dis. Aquat. Organ. 2005, 63, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Sheafor, B.; Davidson, E.W.; Parr, L.; Rollins-Smith, L. Antimicrobial peptide defenses in the salamander, ambystoma tigrinum, against emerging amphibian pathogens. J. Wildl. Dis. 2008, 44, 226–236. [Google Scholar] [CrossRef]
- Whitley, D.S.; Sample, R.C.; Sinning, A.R.; Henegar, J.; Chinchar, V.G. Antisense approaches for elucidating ranavirus gene function in an infected fish cell line. Dev. Comp. Immunol. 2011, 35, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ward, B.M.; Yu, E.K.; Chinchar, V.G.; Robert, J. Improved knockout methodology reveals that frog virus 3 mutants lacking either the 18k immediate-early gene or the truncated vif2-{alpha} gene are defective for replication and growth in vivo. J. Virol. 2011, 85, 11131–11138. [Google Scholar] [CrossRef]
- Langland, J.O.; Cameron, J.M.; Heck, M.C.; Jancovich, J.K.; Jacobs, B.L. Inhibition of pkr by rna and DNA viruses. Virus Res. 2006, 119, 100–110. [Google Scholar] [CrossRef]
- Jancovich, J.K.; Jacobs, B.L. Innate immune evasion mediated by the ambystoma tigrinum virus eukaryotic translation initiation factor 2alpha homologue. J. Virol. 2011, 85, 5061–5069. [Google Scholar] [CrossRef] [PubMed]
- Rothenburg, S.; Chinchar, V.G.; Dever, T.E. Characterization of a ranavirus inhibitor of the antiviral protein kinase pkr. BMC Microbiol. 2011, 11, 56. [Google Scholar] [CrossRef]
- Maniero, G.D.; Carey, C. Changes in selected aspects of immune function in the leopard frog, rana pipiens, associated with exposure to cold. J. Comp. Physiol. B 1997, 167, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Kiesecker, J.M.; Blaustein, A.R.; Belden, L.K. Complex causes of amphibian population declines. Nature 2001, 410, 681–684. [Google Scholar] [CrossRef]
- Bancroft, B.A.; Baker, N.J.; Blaustein, A.R. A meta-analysis of the effects of ultraviolet b radiation and its synergistic interactions with ph, contaminants, and disease on amphibian survival. Conserv. Biol. 2008, 22, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Langerveld, A.J.; Celestine, R.; Zaya, R.; Mihalko, D.; Ide, C.F. Chronic exposure to high levels of atrazine alters expression of genes that regulate immune and growth-related functions in developing xenopus laevis tadpoles. Environ. Res. 2009, 109, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Christin, M.S.; Menard, L.; Gendron, A.D.; Ruby, S.; Cyr, D.; Marcogliese, D.J.; Rollins-Smith, L.; Fournier, M. Effects of agricultural pesticides on the immune system of xenopus laevis and rana pipiens. Aquat. Toxicol. 2004, 67, 33–43. [Google Scholar] [CrossRef]
- Kerby, J.L.; Hart, A.J.; Storfer, A. Combined effects of virus, pesticide, and predator cue on the larval tiger salamander (ambystoma tigrinum). Ecohealth 2011. [Google Scholar] [CrossRef]
- Williams, T.; Barbosa-Solomieu, V.; Chinchar, V.G. A decade of advances in iridovirus research. Adv. Virus Res. 2005, 65, 173–248. [Google Scholar] [PubMed]
- Forson, D.D.; Storfer, A. Atrazine increases ranavirus susceptibility in the tiger salamander, ambystoma tigrinum. Ecol. Appl. 2006, 16, 2325–2332. [Google Scholar] [CrossRef] [PubMed]
- Greer, A.L.; Brunner, J.L.; Collins, J.P. Spatial and temporal patterns of ambystoma tigrinum virus (atv) prevalence in tiger salamanders ambystoma tigrinum nebulosum. Dis. Aquat. Organ. 2009, 85, 1–6. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adults | |||
|---|---|---|---|
| Primary | Secondary | Larvae | |
| Symptoms | 2–3 weeks | 3–5 days | Long lasting, >80% death |
| Virus Clearance | 1 month | 1 week | Ineffective |
| Innate Immunity | |||
| Cells | 1 dpi: Activated Mø | Same + Mø as APC Similar to primary | Mø less resistant to FV3 No NK |
| Induced genes | 3 dpi: NK cells 1 dpi: TNF-α, IL-1β, IFN-γ, Mx1 | Similar to primary | Delayed (3 dpi) and weaker |
| Adaptive T cell immunity | |||
| Splenic CD8 T cell | Peak at 6 dpi | Peak at 3 dpi but lower expansion | ? |
| CD8 T cell in kidneys | At 6 dpi | At 3 dpi but fewer | ? |
| T cell memory | - | yes | ? |
| Adaptive B cell immunity | |||
| Anti-FV3 antibodies | Not detected | IgY from 10 dpi | Not detected |
| More IgY mRNAs | 6 dpi | 3dpi | 6–7 dpi |
| AID up-regulation | 9 dpi | 3 dpi | 6–7 dpi |
| B cell memory | - | Yes | Possibly |
© 2011 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, G.; Robert, J. Antiviral Immunity in Amphibians. Viruses 2011, 3, 2065-2086. https://doi.org/10.3390/v3112065
Chen G, Robert J. Antiviral Immunity in Amphibians. Viruses. 2011; 3(11):2065-2086. https://doi.org/10.3390/v3112065
Chicago/Turabian StyleChen, Guangchun, and Jacques Robert. 2011. "Antiviral Immunity in Amphibians" Viruses 3, no. 11: 2065-2086. https://doi.org/10.3390/v3112065
APA StyleChen, G., & Robert, J. (2011). Antiviral Immunity in Amphibians. Viruses, 3(11), 2065-2086. https://doi.org/10.3390/v3112065