Baculovirus Induced Transcripts in Hemocytes from the Larvae of Heliothis virescens

Abstract
:1. Introduction
2. Results
2.1. Transcriptome Generation and Assembly
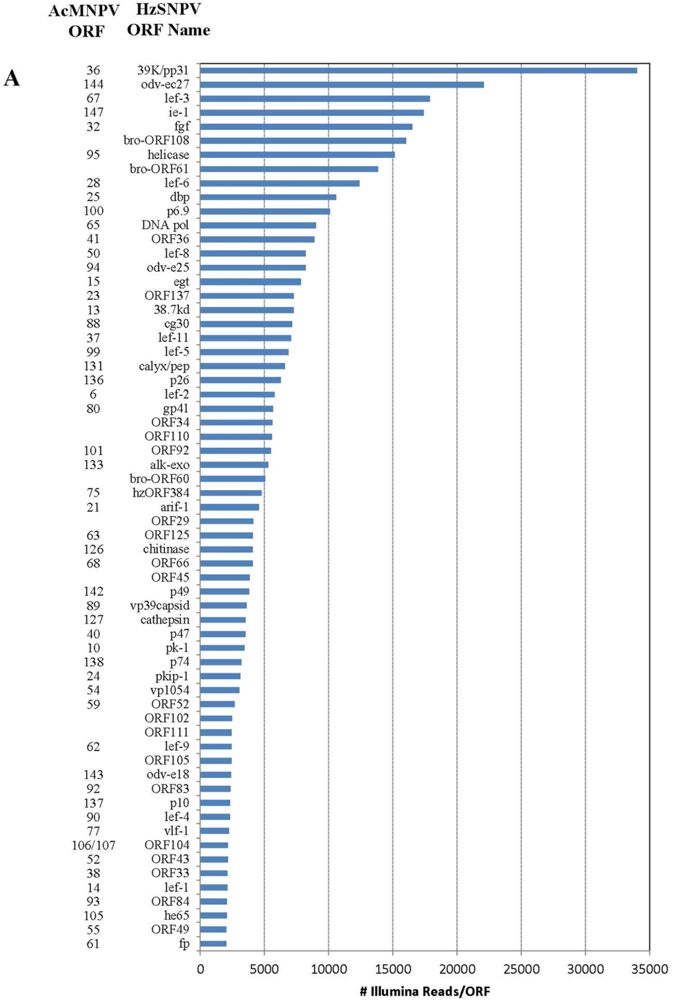
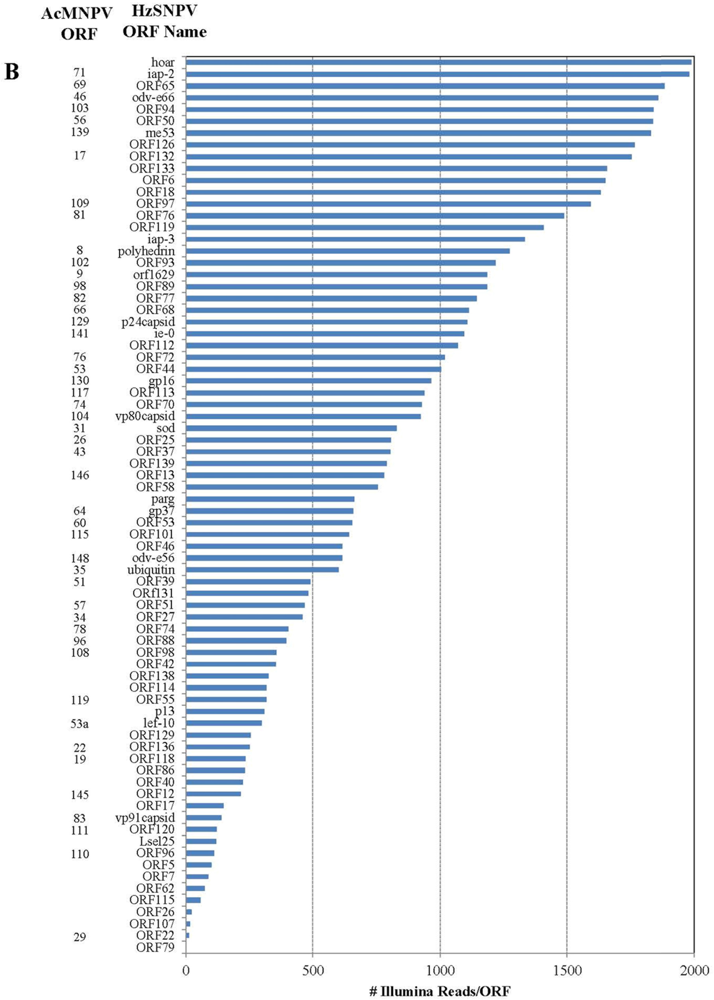
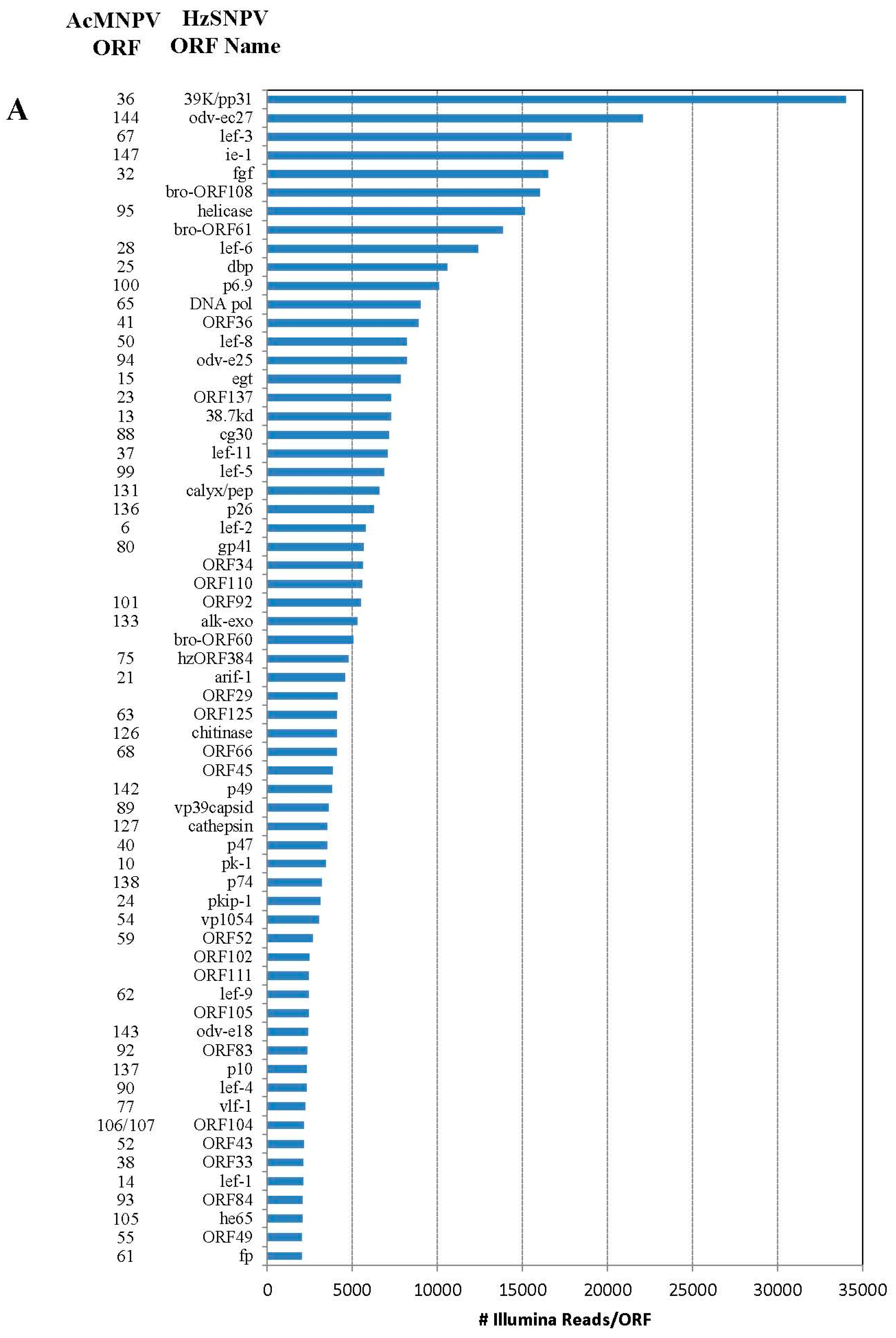
2.2. Illumina Reads Matching the HzSNPV Genome
2.3. HSP70/Small HSPs
2.4. Cellular Adhesion/Immunity Genes
2.5. Endocrine Transcripts
2.6. Metabolic Regulation
2.7. Errantivirus
3. Experimental Section
3.1. Insects, Infection, and RNA Isolation
3.2. Sequence Generation, Assembly and Annotation
3.3. Alignment of Sequence Tags with HzSNPV Reference Genome
3.4. Identification of Differentially Regulated Genes
4. Discussion and Conclusions
Acknowledgements
Conflicts of Interest
References and Notes
- Grabherr, R.; Ernst, W. Baculovirus for eukaryotic protein display. Curr. Gene Ther. 2010, 10, 195–200. [Google Scholar] [CrossRef]
- Liu, C.Y.; Chen, H.Z.; Chao, Y.C. Maximizing baculovirus-mediated foreign proteins expression in mammalian cells. Curr. Gene Ther. 2010, 10, 232–241. [Google Scholar]
- Passarelli, A.L.; Guarino, L.A. Baculovirus late and very late gene regulation. Curr. Drug Targets 2007, 8, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, B.; Hoyos-Carvajal, L.; Paluszek, M.; Skrzecz, I.; Lobo de, S.M. Baculoviruses—Re-emerging biopesticides. Biotechnol. Adv. 2006, 24, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Granados, R.R.; Lawler, K.A.; Burand, J.P. Replication of Heliothis zea baculovirus in an insect cell line. Intervirology 1981, 16, 71–79. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, A.H.; Ignoffo, C.M.; Andrews, P.L. In vitro host range of five baculoviruses in lepidopteran cell lines. Intervirology 1985, 23, 150–156. [Google Scholar] [CrossRef]
- Washburn, J.O.; Wong, J.F.; Volkman, L.E. Comparative pathogenesis of Helicoverpa zea S nucleopolyhedrovirus in noctuid larvae. J. Gen. Virol. 2001, 82, 1777–1784. [Google Scholar] [CrossRef]
- Werner, T. Next generation sequencing allows deeper analysis and understanding of genomes and transcriptomes including aspects to fertility. Reprod. Fertil. Dev. 2011, 23, 75–80. [Google Scholar] [CrossRef]
- Alkan, C.; Coe, B.P.; Eichler, E.E. Genome structural variation discovery and genotyping. Nat. Rev. Genet. 2011, 12, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.W.; Lau, K.C.; Tong, S.F. Microarrays for personalized genomic medicine. Adv. Clin. Chem. 2010, 52, 1–18. [Google Scholar] [PubMed]
- Yao, Q.; Li, M.W.; Wang, Y.; Wang, W.B.; Lu, J.; Dong, Y.; Chen, K.P. Screening of molecular markers for NPV resistance in Bombyx mori L. (Lep., Bombycidae). J. Appl.Entomol. 2003, 127, 134–136. [Google Scholar] [CrossRef]
- Kidokoro, K.; Ito, K.; Ogoyi, D.O.; Abe, H.; Mita, K. Non-susceptibility genes to Bombyx densovirus type 1, Nid-1 and nsd-1, affect distinct steps of the viral infection pathway. J. Invertebr. Pathol. 2010, 103, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Asser-Kaiser, S.; Radtke, P.; El-Salamouny, S.; Winstanley, D.; Jehle, J.A. Baculovirus resistance in codling moth (Cydia pomonella L.) caused by early block of virus replication. Virology 2011, 410, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Sessions, O.M.; Barrows, N.J.; Souza-Neto, J.A.; Robinson, T.J.; Hershey, C.L.; Rodgers, M.A.; Ramirez, J.L.; Dimoupoulos, G.; Yang, P.L.; Pearson, J.L.; Garcia-Blanco, M.A. Discovery of insect and human dengue virus host factors. Nature 2009, 458, 1047–1050. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Vallene, S.; Ramet, M. Drosophila as a model for antiviral immunity. World J. Biol. Chem. 2010, 1, 151–159. [Google Scholar] [CrossRef]
- Sabin, L.R.; Hanna, S.L.; Cherry, S. Innate antiviral immunity in Drosophila. Curr. Opin. Immunol. 2010, 22, 4–9. [Google Scholar] [CrossRef]
- Hao, L.; Sakurai, A.; Watanabe, T.; Sorensen, E.; Nidom, C.A.; Newton, M.A.; Ahlquist, P.; Kawaoka, Y. Drosophila RNAi screen identifies host genes important for influenza virus replication. Nature 2008, 454, 890–893. [Google Scholar] [CrossRef] [PubMed]
- Steinert, S.; Levashina, A.A. Intracellular immune responses of dipteran insects. Immunol. Revs. 2011, 240, 129–140. [Google Scholar] [CrossRef]
- Girard, Y.A.; Mayhew, G.F.; Fuchs, J.F.; Li, H.; Schneider, B.S.; McGee, C.E.; Rocheleau, T.A.; Helmy, H.; Christensen, B.M.; Higgs, S.; Bartholomay, L.C. Transcriptome changes in Culex quinquefasciatus (Diptera: Culicidae) salivary glands during West Nile virus infection. J. Med. Entomol. 2010, 47, 421–435. [Google Scholar] [CrossRef]
- Blair, C.D. Mosquito RNAi is the major innate immune pathway controlling arbovirus infection and transmission. Future Microbiol. 2011, 6, 265–277. [Google Scholar] [CrossRef]
- Yang, Z.; Bruno, D.P.; Martens, C.A.; Porcella, S.F.; Moss, B. Simultaneous high-resolution analysis of vaccinia virus and host cell transcriptomes by deep RNA sequencing. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 11513–11518. [Google Scholar] [CrossRef]
- Bengali, Z.; Satheshkumar, P.S.; Yang, Z.; Weisberg, A.S.; Paran, N.; Moss, B. Drosophila S2 cells are non-permissive for vaccinia virus DNA replication following entry via low pH-dependent endocytosis and early transcription. PLoS One 2011, 6, e17248. [Google Scholar] [CrossRef]
- Clem, R.J.; Popham, H.J.R.; Shelby, K.S. Antiviral respnse in insects: Apoptosis and humoral responses. In Insect Virology, 1st ed.; Asgari, S., Johnson, K., Eds.; Caister Academic Press: St. Lucia, Queensland, Australia, 2011; pp. 389–410. [Google Scholar]
- Whitfield, A.E.; Rotenberg, D.; Aritua, V.; Hogenhout, S.A. Analysis of expressed sequence tags from Maize mosaic rhabdovirus-infected gut tissues of Peregrinus maidis reveals the presence of key components of insect innate immunity. Insect Mol. Biol. 2011, 20, 225–242. [Google Scholar] [CrossRef]
- Luan, J.-B.; Li, J.-M.; Varela, N.; Wang, Y.-L.; Li, F.-F.; Bao, Y.-Y.; Zhang, C.-X.; Liu, S.-S.; Wang, X.-W. Global analysis of the transcriptional response of the whitefly to Tomato Yellow Leaf Curl China Virus reveals the realtionship of coevolved adaptations. J. Virol. 2011, 85, 3330–3340. [Google Scholar] [CrossRef]
- Popham, H.J.R.; Grasela, J.J.; Goodman, C.L.; McIntosh, A.H. Baculovirus infection influences host protein expression in two established insect cell lines. J. Insect Physiol. 2010, 56, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, W.J.; Wong, J.; Chun, G.; Lu, A.; McCutchen, B.F.; Presnail, J.K.; Herrmann, R.; Dolan, M.; Tingey, S.; et al. Comparative analysis of the complete genome sequences of Helicoverpa zea and Helicoverpa armigera single-nucleocapsid nucleopolyhedroviruses. J. Gen. Virol. 2002, 83, 673–684. [Google Scholar] [CrossRef]
- Ayres, M.; Howard, S.C.; Kuzio, J.; Lopez-Ferber, M.; Possee, R.D. The complete DNA sequence of Autographa californica nuclear polyhedrosis virus. Virology 1994, 202, 586–605. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Valente, V.L.; Loreto, E.L. Multiple invasions of Errantivirus in the genus Drosophila. Insect Mol. Biol. 2008, 17, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Popham, H.J.R.; Shelby, K.S.; Popham, T.W. Effect of dietary selenium supplementation on resistance to baculovirus infection. Biol. Contr. 2005, 32, 419–426. [Google Scholar] [CrossRef]
- Terenius, O.; Popham, H.J.R.; Shelby, K.S. Bacterial, but not baculoviral infections stimulate Hemolin expression in noctuid moths. Dev. Comp. Immunol. 2009, 33, 1176–1185. [Google Scholar] [CrossRef]
- Shelby, K.S.; Popham, H.J.R. Plasma phenoloxidase of larval Heliothis virescens is virucidal. J. Insect Sci. 2006, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shelby, K.S.; Popham, H.J.R. Analysis of ESTs generated from immune-stimulated hemocytes of larval Heliothis virescens. J. Invertebr. Pathol. 2009, 101, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Marioni, J.C.; Mason, C.E.; Mane, S.M.; Stephens, M.; Gilad, Y. RNA-seq: An assessment of technical reproducibility and comparison with gene expression arrays. Genome Res. 2008, 18, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- FASTX-Tool Kit FASTQ/A Short-Reads Pre-Processing Tools. Available online: hannonlab.cshl.edu/fastx_toolkit/ (accessed on 10 February 2011).
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Shulz, M.; Zerbino, D. Oases De novo assembler for very short reads. Available online: http://www.ebi.ac.uk/~zerbino/oases/j/ (accessed on 10 February 2011).
- Kurtz, S. The Vmatch large scale sequence analysis software. Available online: http://www.vmatch.de/ (accessed on 12 February 2011).
- Jain, E.; Bairoch, A.; Duvaud, S.; Phan, I.; Redaschi, N.; Suzek, B.E.; Martin, M.J.; McGarvey, P.; Gasteiger, E. Infrastructure for the life sciences: Design and implementation of the UniProt website. BMC Bioinformatics 2009, 10, 136–144. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Heliothis virescens Transcriptome Project. Available online: http://www.ncbi.nlm.nih.gov/bioproject/49697/ (accessed on 20 July 2011).
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef]
- Washburn, J.O.; Wong, J.F. , Volkman, L.E. Comparative pathogenesis of Helicoverpa zea S nucleopolyhedrovirus in noctuid larvae. J. Gen. Virol. 2001, 82, 1777–1784. [Google Scholar] [CrossRef]
- Washburn, J.O.; Trudeau, D.; Wong, J.F. , Volkman, L.E. Early pathogenesis of Autographa californica multiple nucleopolyhedrovirus and Helicoverpa zea single nucleopolyhedrovirus in Heliothis virescens: A comparison of the ‘M’ and ‘S’ strategies for establishing fatal infection. J. Gen. Virol. 2003, 84, 343–351. [Google Scholar] [CrossRef]
- Breitenbach, J.E.; Popham, H.J.R. Baculovirus infection induces heat shock response in vitro and in vivo. USDA, ARS, BCIRL, Columbia, MO, USA. To be submitted for publication. 2011. [Google Scholar]
- Lyupina, Y.V.; Dmitrieva, S.B.; Timokhova, A.V.; Beljelarskaya, S.N.; Zatsepina, O.G.; Evgen’ev, M.B.; Mikhailov, V.S. An important role of the heat shock response in infected cells for replication of baculoviruses. Virology 2010, 406, 336–341. [Google Scholar] [CrossRef]
- Dustin, M. Viral spread through protoplasmic kiss. Nat. Cell Biol. 2003, 5, 271–272. [Google Scholar] [CrossRef] [PubMed]
- Hutt-Fletcher, L.M.; Chesnokova, L.S. Integrins as triggers of Epstein-Barr virus fusion and epithelial cell infection. Virulence 2010, 1, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, A.R.; Nansen, A.; Madsen, A.N.; Bartholdy, C.; Christensen, J.P. Regulation of T cell migration during viral infection: role of adhesion molecules and chemokines. Immunol. Lett. 2003, 85, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Nardi, J.B.; Bee, C.M.; Miller, L.A.; Mathur, D.; Ohlstein, B. Cell renewal in adjoining intestinal and tracheal epithelia of Manduca. J. Insect Physiol. 2011, 57, 487–493. [Google Scholar] [CrossRef]
- Hebbar, S.; Fernandes, J.J. Glial remodeling during metamorphosis influences the stabilization of motor neuron branches in Drosophila. Dev. Biol. 2010, 340, 344–354. [Google Scholar] [CrossRef]
- Mao, Y.; Freeman, M. Fasciclin 2, the Drosophila orthologue of neural cell-adhesion molecule, inhibits EGF receptor signalling. Development 2009, 136, 473–481. [Google Scholar] [CrossRef]
- Nassif, C.; Noveen, A.; Hartenstein, V. Early development of the Drosophila brain: III. The pattern of neuropile founder tracts during the larval period. J. Comp Neurol. 2003, 455, 417–434. [Google Scholar] [CrossRef]
- Lemaitre, B. The road to toll. Nat. Rev. Immunol. 2004, 4, 521–527. [Google Scholar] [CrossRef]
- Dimopoulos, G.; Muller, H.M.; Levashina, E.A.; Kafatos, F.C. Innate immune defense against malaria infection in the mosquito. Curr. Opin. Immunol. 2001, 13, 79–88. [Google Scholar] [CrossRef]
- Ueno, K.; Suzuki, Y. p260/270 expressed in embryonic abdominal leg cells of Bombyx mori can transfer palmitate to peptides. J. Biol. Chem. 1997, 272, 13519–13526. [Google Scholar] [CrossRef]
- Lourenco, A.P.; Martins, J.R.; Bitondi, M.M.; Simoes, Z.L. Trade-off between immune stimulation and expression of storage protein genes. Arch. Insect Biochem. Physiol. 2009, 71, 70–87. [Google Scholar] [CrossRef]
- Shelby, K.S.; Webb, B.A. Polydnavirus infection inhibits synthesis of an insect plasma protein, arylphorin. J. Gen. Virol. 1994, 75, 2285–2292. [Google Scholar] [CrossRef]
- O’Reilly, D.R. Baculovirus-encoded ecdysteroid UDP-glucosyltransferases. Insect Biochem. Mol. Biol. 1995, 25, 541–550. [Google Scholar] [CrossRef]
- Bonning, B.C.; Ward, V.K.; van Meer, M.M.; Booth, T.F.; Hammock, B.D. Disruption of lysosomal targeting is associated with insecticidal potency of juvenile hormone esterase. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 6007–6012. [Google Scholar] [CrossRef]
- Wroblewski, V.J.; Harshman, L.G.; Hanzlik, T.N.; Hammock, B.D. Regulation of juvenile hormone esterase gene expression in the tobacco budworm (Heliothis virescens). Arch. Biochem. Biophys. 1990, 278, 461–466. [Google Scholar] [CrossRef]
- Menzel, T.; Rohrmann, G.F. Diversity of errantivirus (retrovirus) sequences in two cell lines used for baculovirus expression, Spodoptera frugiperda and Trichoplusia ni. Virus Genes 2008, 36, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Rohrmann, G.F. Envelope gene capture and insect retrovirus evolution: the relationship between errantivirus and baculovirus envelope proteins. Virus Res. 2006, 118, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.W.; Miller, L.K. A virus mutant with an insertion of a copia-like transposable element. Nature 1982, 299, 562–564. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Total Seqs | After Cleaning | After Decontamination | Aligned | Uniquely Aligned |
|---|---|---|---|---|---|
| Control | 50396515 | 48734654 | 48646727 | 31308740 | 19809154 |
| Bacterial | 53593477 | 51046869 | 51000867 | 32021434 | 21516663 |
| Fungal | 44558357 | 42149252 | 42139873 | 25948555 | 17600091 |
| Viral | 53944832 | 50600536 | 50575131 | 32107380 | 20716956 |
| HzSNPV ORF | HzSNPV ORF Name | Corresponding AcMNPV ORF | ORF Start | ORF End | # of Hits Unique to ORF | # of Hits ORF | Difference in Hits |
|---|---|---|---|---|---|---|---|
| 26 | ORF26 | 23950 | 24102 | 22 | 29 | 7 | |
| 27 | ORF27 | 34 | 24045 | 24812 | 459 | 1256 | 797 |
| 28 | ubiquitin | 35 | 24652 | 24903 | 600 | 1390 | 790 |
| 46 | ORF46 | 39876 | 40103 | 616 | 618 | 2 | |
| 47 | lef-10 | 53a | 40063 | 40278 | 298 | 676 | 378 |
| 48 | vp1054 | 54 | 40151 | 41206 | 3032 | 3408 | 376 |
| 75 | gp41 | 80 | 65470 | 66438 | 5648 | 5791 | 143 |
| 76 | ORF76 | 81 | 66368 | 67093 | 1488 | 2135 | 647 |
| 77 | ORF77 | 82 | 66966 | 67643 | 1146 | 1669 | 523 |
| 78 | vp91capsid | 83 | 67573 | 70023 | 140 | 159 | 19 |
| 86 | viral_ppase | 98 | 80264 | 81229 | 1185 | 1361 | 176 |
| 87 | lef-5 | 99 | 81125 | 82072 | 6877 | 7053 | 176 |
| 127 | ORF127 | 119843 | 120421 | 481 | 489 | 8 | |
| 128 | BV-ec31 | 17 | 120372 | 121172 | 1754 | 1762 | 8 |
| Function/Gene ID | Control | NPV | Fold Change | Length | %Cov | e-value |
|---|---|---|---|---|---|---|
| Heat shock | ||||||
| Heat shock protein 70 Helicoverpa zea | 24 | 4238 | 176.6 | 1452 | 69 | 0 |
| Heat shock protein 70 Spodoptera exigua | 30 | 2122 | 70.7 | 1763 | 98 | 0 |
| PREDICTED: similar to HSP70 Acyrthosiphon pisum | 1 | 85 | 85.0 | 170 | 100 | 6E-05 |
| Hsp70-interacting protein H. armigera | 23 | 1934 | 84.1 | 599 | 94 | 2E-126 |
| Heat shock protein 68 Drosophila yakuba | 1 | 72 | 72.0 | 150 | 100 | 1E-39 |
| Heat shock protein 19.5 Sesamia nonagrioides | 3 | 64 | 21.3 | 171 | 100 | 7E-13 |
| Heat shock protein hsp20.4 Bombyx mori | 1 | 21 | 21.0 | 200 | 100 | 2E-08 |
| Heat shock protein 19.7 Mamestra brassicae | 1 | 19 | 19.0 | 87 | 100 | 4E-12 |
| Heat shock protein 20.8 Bombyx mori | 5 | 80 | 16.0 | 105 | 100 | 5 E-45 |
| Cell action | ||||||
| EGF-containing fibulin-like ECM prot. 2 Macaca fascicularis | 65 | 462 | 7.1 | 2092 | 100 | 2E-64 |
| Cecropin D Spodoptera litura | 1 | 12 | 12.0 | 263 | 64 | 1E-12 |
| Larval cuticle protein Bombyx mori | 4 | 26 | 6.5 | 163 | 100 | 5E-25 |
| Dystroglycan Bos taurus | 5990 | 1575 | 0.3 | 2924 | 60 | 2E-44 |
| Signal transducing adapter molecule 2 Rattus norvegicus | 420 | 88 | 0.2 | 417 | 100 | 3E-13 |
| Fasciclin-1 Schistocerca americana | 1383 | 23 | 0.02 | 755 | 72 | 1E-103 |
| Function/Gene ID | Control | NPV | Fold Change | Length | %Cov | e-value |
|---|---|---|---|---|---|---|
| Hormone | ||||||
| Probable nuclear hormone receptor HR3 Manduca sexta | 33 | 253 | 7.7 | 99 | 100 | 7E-34 |
| Nuclear hormone receptor FTZ-F1 Bombyx mori | 766 | 3232 | 4.2 | 2867 | 98 | 0 |
| Juvenile hormone epoxide hydrolase Helicoverpa armigera | 25 | 89 | 3.6 | 657 | 100 | 3E-22 |
| Hormone receptor 3C Choristoneura fumiferana | 5 | 15 | 3.0 | 124 | 100 | 3E-13 |
| Basic juvenile hormone-suppressible protein 1 Trichoplusia ni | 2575 | 67 | 0.03 | 2279 | 78 | 0 |
| Juvenile hormone diol kinase Manduca sexta | 58 | 11 | 0.2 | 567 | 100 | 4E-59 |
| Basic juvenile hormone-suppressible protein 2 Trichoplusia ni | 199 | 1 | 0.01 | 2170 | 40 | 0 |
| Lipid Metabolism | ||||||
| P260 Bombyx mori | 15 | 6420 | 428.0 | 4896 | 92 | 0 |
| Stearoyl-CoA desaturase Aedes aegypti | 1 | 237 | 237.0 | 775 | 87 | 1E-74 |
| Fatty-acyl CoA reductase Ostrinia nubilalis | 5 | 1038 | 207.6 | 1520 | 88 | 2E-179 |
| Fatty acid synthase (Fragment) Heliothis virescens | 2 | 149 | 74.5 | 548 | 93 | 8E-100 |
| Acyl-CoA delta-9 desaturase isoform Helicoverpa zea | 19 | 1922 | 101.16 | 1238 | 94 | 7E-131 |
| P270 Bombyx mori | 6 | 306 | 51.0 | 959 | 81 | 2E-133 |
| Fatty acyl-CoA desaturase Glossina morsitans | 1 | 38 | 38.0 | 190 | 100 | 6E-16 |
| Acyl-CoA desaturase (Fragment) Heliothis virescens | 0 | 36 | 36.0 | 215 | 100 | 2E-37 |
| Acyl-CoA-delta-6-desaturase Antheraea pernyi | 0 | 19 | 19.0 | 141 | 100 | 2E-12 |
| Acyl-CoA oxidase (Fragment) H. virescens | 0 | 25 | 25.0 | 295 | 100 | 2E-29 |
| PREDICTED: similar to acyl-CoA oxidase Acyrthosiphon pisum | 0 | 24 | 24.0 | 145 | 61 | 6E-21 |
| Peroxisomal acyl-CoA oxidase, putative Ixodes scapularis | 1 | 29 | 29.0 | 252 | 96 | 4E-24 |
| Acyl-CoA synthetase short-chain family member 3, mitochondrial Camponotus floridanus | 0 | 16 | 16.0 | 159 | 89 | 7E-22 |
| Acyl-CoA desaturase PintNPRD (Fragment) Plodia interpunctella | 0 | 12 | 12.0 | 158 | 56 | 5E-24 |
| Lipid storage droplets surface-binding protein Drosophila melanogaster | 22 | 70 | 3.2 | 494 | 85 | 2E-31 |
| Fatty acid beta-oxidation complex subunit beta Heliothis virescens | 11 | 34 | 3.1 | 204 | 92 | 2E-12 |
| Glycine C-acetyltransferase/2-amino-3-ketobutyrate- CoA ligase Ixodes scapularis | 45 | 11 | 0.2 | 185 | 100 | 2E-21 |
| Fatty acid-binding protein 3 Helicoverpa armigera | 25 | 5 | 0.2 | 65 | 100 | 1E-11 |
| Probable malonyl-CoA-acyl carrier protein transacylase Drosophila melanogaster | 171 | 23 | 0.13 | 272 | 100 | 1E-14 |
| Low-density lipoprotein receptor-related protein 1 Gallus gallus | 45 | 3 | 0.07 | 97 | 100 | 2E-9 |
| Storage | ||||||
| Arylphorin subunit Spodoptera litura | 2646 | 172 | 0.07 | 1324 | 84 | 0 |
| Hexamerine Helicoverpa armigera | 100 | 0 | 0.01 | 210 | 100 | 4E-145 |
| P82 riboflavin-binding hexamer Heliothis virescens | 19 | 1 | 0.05 | 353 | 93 | 2E-13 |
| Gene ID | Control | NPV | Fold Change | Length | %Cov | e-value |
|---|---|---|---|---|---|---|
| Endonuclease-reverse transcriptase Bombyx mori | 4 | 45 | 11.2 | 260 | 100 | 3E-23 |
| RNase H and integrase-like protein (Fragment) Bombyx mori | 6 | 60 | 10.0 | 162 | 100 | 4E-6 |
| PREDICTED: similar to Copia protein (Gag-int-pol protein) Tribolium castaneum | 49 | 402 | 8.2 | 1164 | 97 | 1E-101 |
| PREDICTED: similar to putative gag-pol protein, partial Acyrthosiphon pisum | 6 | 48 | 8.0 | 589 | 100 | 1E-35 |
| RNA-directed DNA polymerase (Reverse transcriptase) Medicago truncatula | 60 | 366 | 6.1 | 980 | 100 | 1E-88 |
| Protease and reverse transcriptase-like Protein Bombyx Mori | 65 | 386 | 5.9 | 928 | 100 | 3E-86 |
| Putative gag-pol polyprotein Aster yellows phytoplasma | 36 | 202 | 5.6 | 939 | 100 | 2E-43 |
| Gag-like protein Bombyx mori | 68 | 368 | 5.4 | 799 | 89 | 2E-61 |
| PREDICTED: similar to protease, reverse transcriptase, ribonuclease H, integrase Tribolium castaneum | 11 | 49 | 4.5 | 113 | 100 | 5E-23 |
| PREDICTED: similar to gag-pol polyprotein Tribolium Castaneum | 75 | 304 | 4.1 | 1025 | 100 | 6E-35 |
| PREDICTED: similar to protease, reverse transcriptase, ribonuclease H, integrase Tribolium castaneum | 27 | 103 | 3.8 | 307 | 78 | 1E-18 |
| Enzymatic polyprotein; Endonuclease; Reverse transcriptase, putative Pediculus humanus corporis | 15 | 55 | 3.7 | 355 | 100 | 1E-19 |
| Putative uncharacterized protein (Gag-pol polyprotein) Drosophila melanogaster | 26 | 87 | 3.4 | 765 | 80 | 6E-52 |
| PREDICTED: similar to Copia protein (Gag-int-pol protein) Tribolium castaneum | 110 | 0 | 0.01 | 448 | 100 | 2E-31 |
© 2011 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Breitenbach, J.E.; Shelby, K.S.; Popham, H.J.R. Baculovirus Induced Transcripts in Hemocytes from the Larvae of Heliothis virescens. Viruses 2011, 3, 2047-2064. https://doi.org/10.3390/v3112047
Breitenbach JE, Shelby KS, Popham HJR. Baculovirus Induced Transcripts in Hemocytes from the Larvae of Heliothis virescens. Viruses. 2011; 3(11):2047-2064. https://doi.org/10.3390/v3112047
Chicago/Turabian StyleBreitenbach, Jonathan E., Kent S. Shelby, and Holly J.R. Popham. 2011. "Baculovirus Induced Transcripts in Hemocytes from the Larvae of Heliothis virescens" Viruses 3, no. 11: 2047-2064. https://doi.org/10.3390/v3112047
APA StyleBreitenbach, J. E., Shelby, K. S., & Popham, H. J. R. (2011). Baculovirus Induced Transcripts in Hemocytes from the Larvae of Heliothis virescens. Viruses, 3(11), 2047-2064. https://doi.org/10.3390/v3112047