Meta-Analysis of High-Throughput Datasets Reveals Cellular Responses Following Hemorrhagic Fever Virus Infection

Abstract
:Acknowledgments
Conflict of Interest
References and Notes
- Calvano, S.E.; Xiao, W.; Richards, D.R.; Felciano, R.M.; Baker, H.V.; Cho, R.J.; Chen, R.O.; Brownstein, B.H.; Cobb, J.P.; Tschoeke, S.K.; et al. A network-based analysis of systemic inflammation in humans. Nature 2005, 437, 1032–1037. [Google Scholar] [CrossRef]
- McLeay, R.C.; Bailey, T.L. Motif Enrichment Analysis: A unified framework and an evaluation on ChIP data. BMC Bioinformatics 2010, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Aerts, S.; Thijs, G.; Coessens, B.; Staes, M.; Moreau, Y.; De Moor, B. Toucan: Deciphering the cis-regulatory logic of coregulated genes. Nucl. Acid. Res. 2003, 31, 1753–1764. [Google Scholar] [CrossRef] [PubMed]
- Fennewald, S.M.; Scott, E.P.; Zhang, L.; Yang, X.; Aronson, J.F.; Gorenstein, D.G.; Luxon, B.A.; Shope, R.E.; Beasley, D.W.; Barrett, A.D.; et al. Thioaptamer decoy targeting of AP-1 proteins influences cytokine expression and the outcome of arenavirus infections. J. Gen. Virol. 2007, 88, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Bowick, G.C.; Fennewald, S.M.; Elsom, B.L.; Aronson, J.F.; Luxon, B.A.; Gorenstein, D.G.; Herzog, N.K. Differential signaling networks induced by mild and lethal hemorrhagic fever virus infections. J. Virol. 2006, 80, 10248–10252. [Google Scholar] [CrossRef] [PubMed]
- Bowick, G.C.; Fennewald, S.M.; Scott, E.P.; Zhang, L.H.; Elsom, B.L.; Aronson, J.F.; Spratt, H.M.; Luxon, B.A.; Gorenstein, D.G.; Herzog, N.K. Identification of differentially activated cell-signaling networks associated with Pichinde virus pathogenesis by using systems kinomics. J. Virol. 2007, 81, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Bowick, G.C.; Fennewald, S.M.; Zhang, L.H.; Yang, X.B.; Aronson, J.F.; Shope, R.E.; Luxon, B.A.; Gorenstein, D.G.; Herzog, N.K. Attenuated and Lethal Variants of Pichinde Virus Induce Differential Patterns of NF-kappa B Activation Suggesting a Potential Target for Novel Therapeutics. Viral Immunol. 2009, 22, 457–462. [Google Scholar] [CrossRef]
- Bowick, G.C.; Spratt, H.M.; Hogg, A.E.; Endsley, J.J.; Wiktorowicz, J.E.; Kurosky, A.; Luxon, B.A.; Gorenstein, D.G.; Herzog, N.K. Analysis of the differential host cell nuclear proteome induced by attenuated and virulent hemorrhagic arenavirus infection. J. Virol. 2009, 83, 687–700. [Google Scholar] [CrossRef]
- Hartman, A.L.; Ling, L.; Nichol, S.T.; Hibberd, M.L. Whole-genome expression profiling reveals that inhibition of host innate immune response pathways by Ebola virus can be reversed by a single amino acid change in the VP35 protein. J. Virol. 2008, 82, 5348–5358. [Google Scholar] [CrossRef]
- Djavani, M.; Crasta, O.R.; Zhang, Y.; Zapata, J.C.; Sobral, B.; Lechner, M.G.; Bryant, J.; Davis, H.; Salvato, M.S. Gene expression in primate liver during viral hemorrhagic fever. Virol. J. 2009, 6, 20. [Google Scholar] [CrossRef]
- Djavani, M.M.; Crasta, O.R.; Zapata, J.C.; Fei, Z.; Folkerts, O.; Sobral, B.; Swindells, M.; Bryant, J.; Davis, H.; Pauza, C.D.; et al. Early blood profiles of virus infection in a monkey model for lassa fever. J. Virol. 2007, 81, 7960–7973. [Google Scholar] [CrossRef] [PubMed]
- do Valle, T.Z.; Billecocq, A.; Guillemot, L.; Alberts, R.; Gommet, C.; Geffers, R.; Calabrese, K.; Schughart, K.; Bouloy, M.; Montagutelli, X.; et al. A new mouse model reveals a critical role for host innate immunity in resistance to Rift Valley fever. J. Immunol. 2010, 185, 6146–6156. [Google Scholar] [CrossRef] [PubMed]
- Ingenuity Pathway Analysis application (IPA), Version 9.0, Content Version 3206; Ingenuity Systems: Redwood City, CA, USA, 2011.
- Al-Hazmi, M.; Ayoola, E.A.; Abdurahman, M.; Banzal, S.; Ashraf, J.; El-Bushra, A.; Hazmi, A.; Abdullah, M.; Abbo, H.; Elamin, A.; et al. Epidemic Rift Valley fever in Saudi Arabia: A clinical study of severe illness in humans. Clin. Infect. Dis. 2003, 36, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Fisher-Hoch, S.P.; Platt, G.S.; Neild, G.H.; Southee, T.; Baskerville, A.; Raymond, R.T.; Lloyd, G.; Simpson, D.I. Pathophysiology of shock and hemorrhage in a fulminating viral infection (Ebola). J. Infect. Dis. 1985, 152, 887–894. [Google Scholar] [CrossRef]
- Zambelli, F.; Pesole, G.; Pavesi, G. Pscan: Finding over-represented transcription factor binding site motifs in sequences from co-regulated or co-expressed genes. Nucl. Acid. Res. 2009, 37, W247–W252. [Google Scholar] [CrossRef]
- Matys, V.; Kel-Margoulis, O.V.; Fricke, E.; Liebich, I.; Land, S.; Barre-Dirrie, A.; Reuter, I.; Chekmenev, D.; Krull, M.; Hornischer, K.; et al. TRANSFAC and its module TRANSCompel: transcriptional gene regulation in eukaryotes. Nucl. Acid. Res. 2006, 34, D108–D110. [Google Scholar] [CrossRef]
- Vela, E.M.; Bowick, G.C.; Herzog, N.K.; Aronson, J.F. Genistein treatment of cells inhibits arenavirus infection. Antivir. Res. 2008, 77, 153–156. [Google Scholar] [CrossRef]
- Vela, E.M.; Bowick, G.C.; Herzog, N.K.; Aronson, J.F. Exploring kinase inhibitors as therapies for human arenavirus infections. Future Virol. 2008, 3, 243–251. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Young, H.A.; Jahrling, P.B.; Davis, K.J.; Larsen, T.; Kagan, E.; Hensley, L.E. Pathogenesis of Ebola hemorrhagic fever in primate models—Evidence that hemorrhage is not a direct effect of virus-induced cytolysis of endothelial cells. Amer. J. Pathol. 2003, 163, 2371–2382. [Google Scholar] [CrossRef]
- Wu, W.L.; Ho, L.J.; Chang, D.M.; Chen, C.H.; Lai, J.H. Triggering of DC migration by dengue virus stimulation of COX-2-dependent signaling cascades in vitro highlights the significance of these cascades beyond inflammation. Eur. J. Immunol. 2009, 39, 3413–3422. [Google Scholar] [CrossRef]
- Bowick, G.C.; Barrett, A.D. Comparative pathogenesis and systems biology for biodefense virus vaccine development. J. Biomed. Biotechnol. 2010, 2010, 236528. [Google Scholar] [CrossRef] [PubMed]



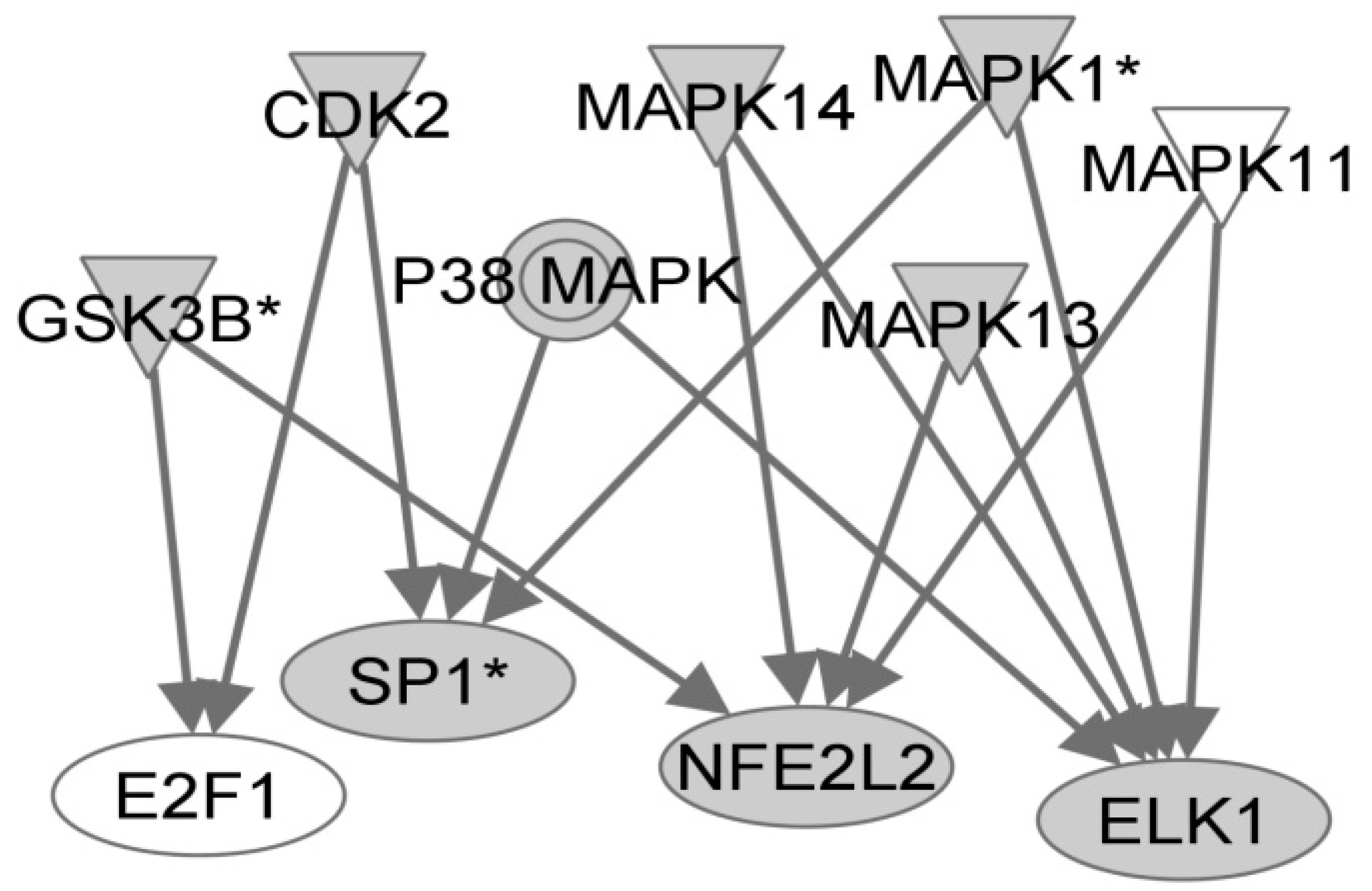{kind=link}
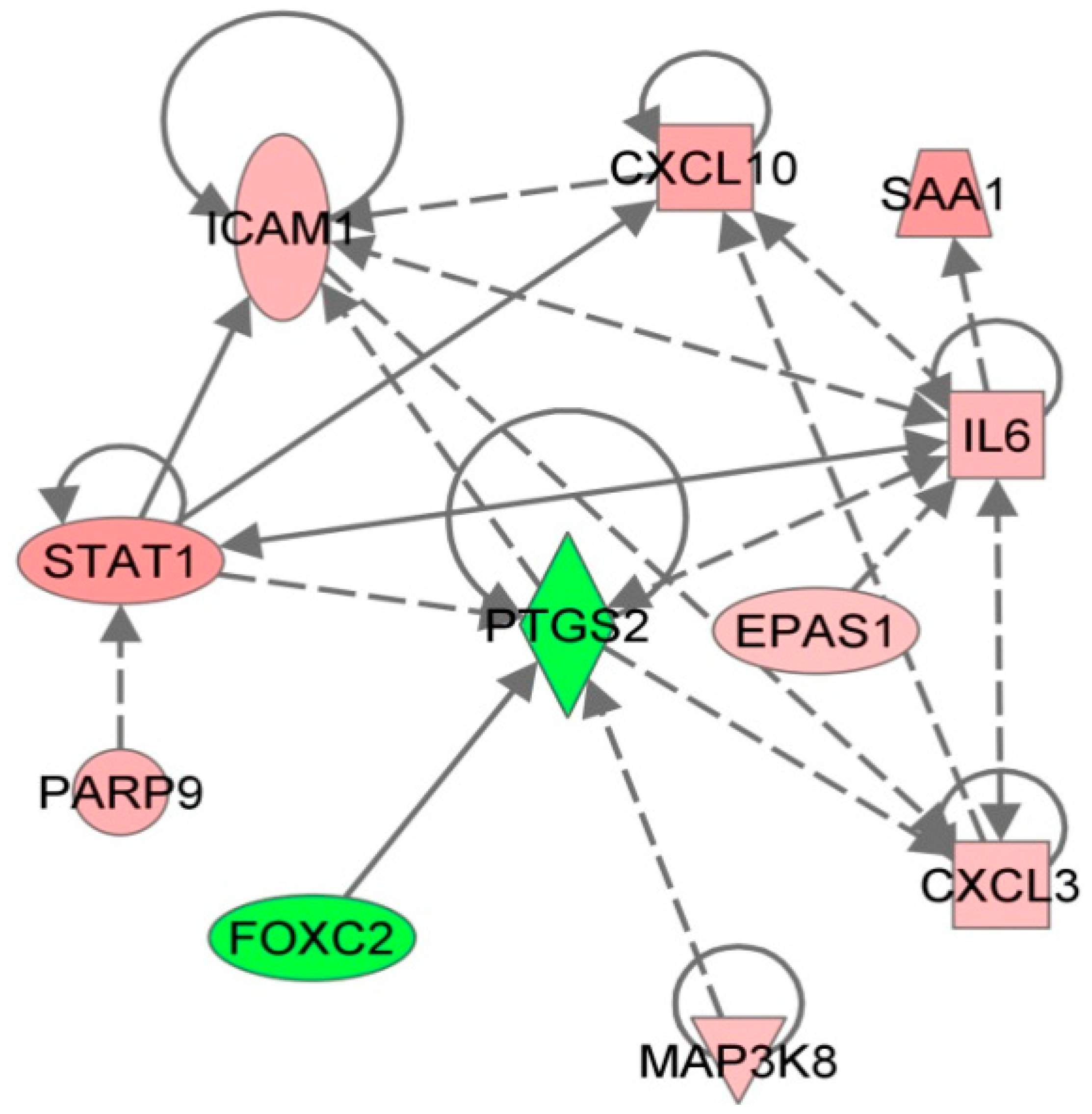{kind=link}
| EBOV | RVFV | LCMV (PBMC) | LCMV (LIVER) |
|---|---|---|---|
| FAC1 (0.017) | ISRE1 (1.2 × 10−15) | SP1 (1.2 × 10−22) | E2F (2.8 × 10−18) |
| NFY (0.018) | IRF7 (2.4 × 10−10) | E2F (3.9 × 10−16) | ELK1 (5.6 × 10−17) |
| ATF6 (0.021) | IRF2 (3.6 × 10−6) | ELK1 (2 × 10−14) | SP1 (3.7 × 10−15) |
| FOXO4 (0.021) | AP1 (1.3 × 10−5) | EGR3 (8.7 × 10−12) | ATF (3.5 × 10−11) |
| RFX1 (0.022) | NF-κB (3 × 10−4) | NRF2 (9.4 × 10−11) | AP2 (6.3 × 10−11) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2011 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowick, G.C.; McAuley, A.J. Meta-Analysis of High-Throughput Datasets Reveals Cellular Responses Following Hemorrhagic Fever Virus Infection. Viruses 2011, 3, 613-619. https://doi.org/10.3390/v3050613
Bowick GC, McAuley AJ. Meta-Analysis of High-Throughput Datasets Reveals Cellular Responses Following Hemorrhagic Fever Virus Infection. Viruses. 2011; 3(5):613-619. https://doi.org/10.3390/v3050613
Chicago/Turabian StyleBowick, Gavin C., and Alexander J. McAuley. 2011. "Meta-Analysis of High-Throughput Datasets Reveals Cellular Responses Following Hemorrhagic Fever Virus Infection" Viruses 3, no. 5: 613-619. https://doi.org/10.3390/v3050613
APA StyleBowick, G. C., & McAuley, A. J. (2011). Meta-Analysis of High-Throughput Datasets Reveals Cellular Responses Following Hemorrhagic Fever Virus Infection. Viruses, 3(5), 613-619. https://doi.org/10.3390/v3050613