The Prevalence and Significance of HTLV-I/II Seroindeterminate Western Blot Patterns

Abstract
:1. Background
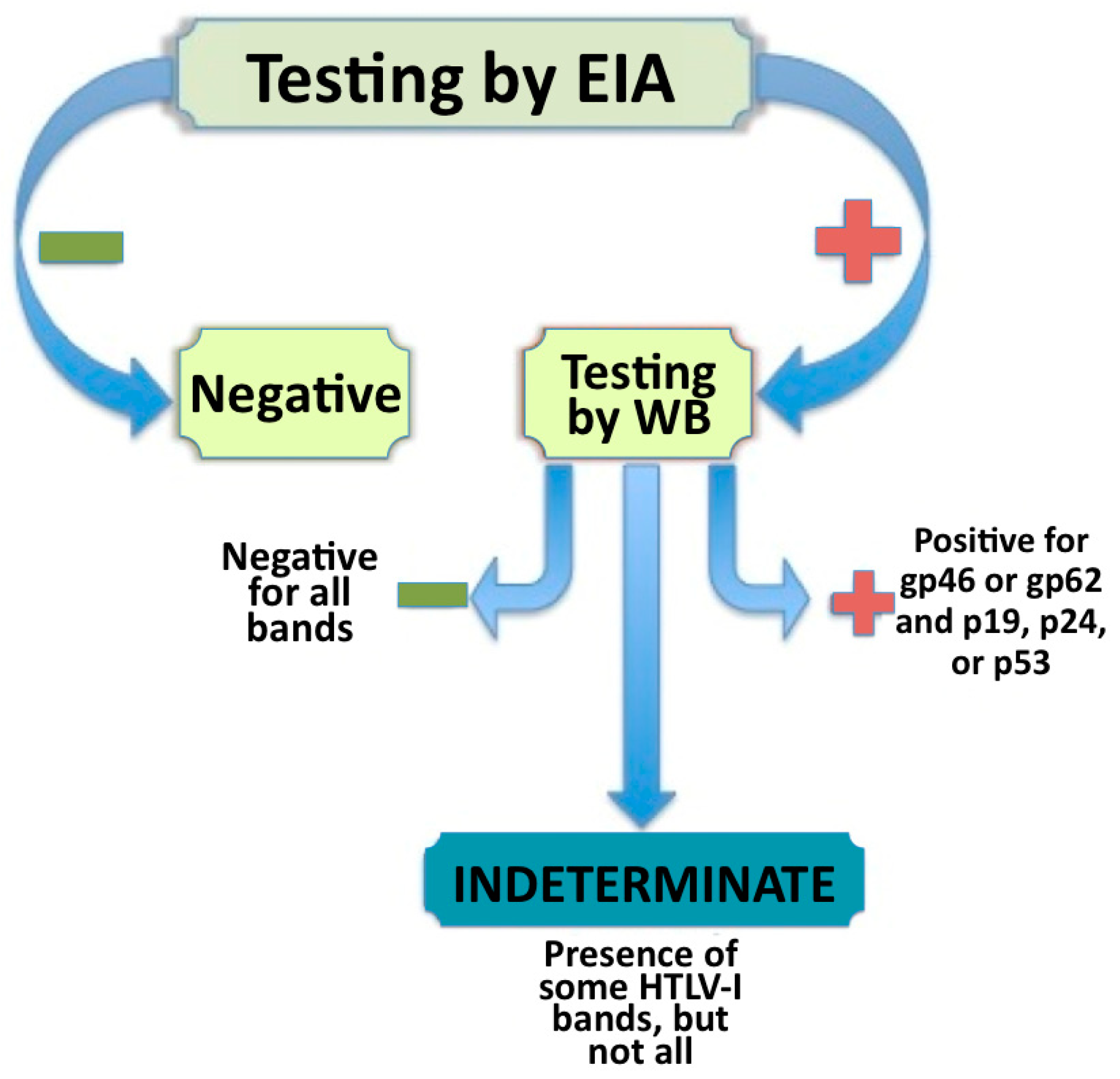
2. Defining and Identifying Seroindeterminates
3. Possible Explanations for HTLV-I/II Seroindeterminates
3.1. Malaria and Severe Acute Respiratory Syndrome (SARS)
3.2. Cross-Reactivities to Other Retrovirus
3.3. Low Copy Number of Prototype HTLV-I
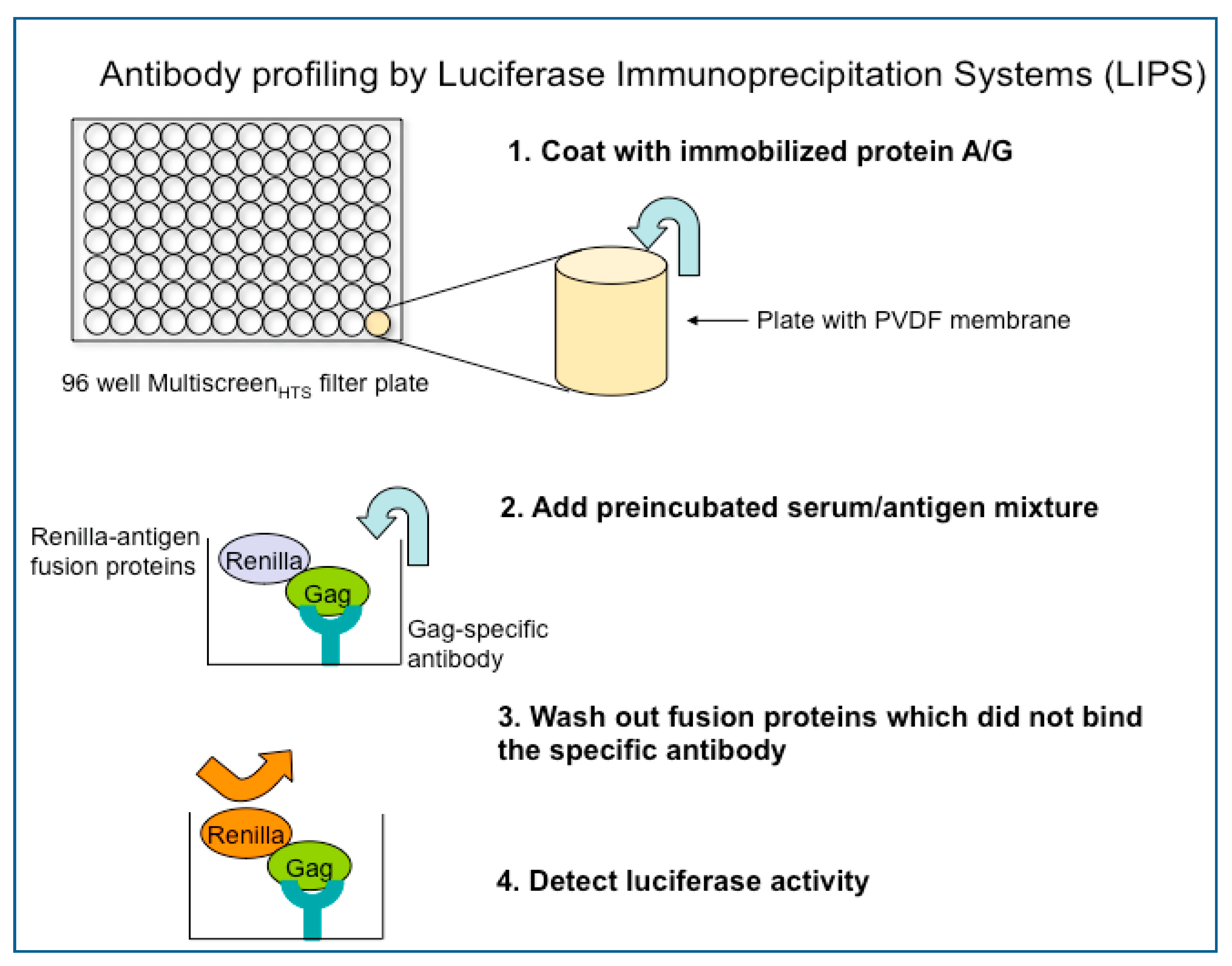
4. Current Research and Future Directions

Acknowledgements
References and Notes
- Gallo, R.C. The discovery of the first human retrovirus: HTLV-1 and HTLV-2. Retrovirology 2005, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Proietti, F.A.; Carneiro-Proietti, A.B.; Catalan-Soares, B.C.; Murphy, E.L. Global epidemiology of HTLV-I infection and associated diseases. Oncogene 2005, 24, 6058–6068. [Google Scholar] [CrossRef] [PubMed]
- Takatsuki, K. Discovery of adult T-cell leukemia. Retrovirology 2005, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Coligan, J.E.; Essex, M. Human T-cell leukemia virus specific antigens. Princess Takamatsu Symp. 1984, 15, 197–203. [Google Scholar]
- Merl, S.; Kloster, B.; Moore, J.; Hubbell, C.; Tomar, R.; Davey, F.; Kalinowski, D.; Planas, A.; Ehrlich, G.; Clark, D.; et al. Efficient transformation of previously activated and dividing T lymphocytes by human T cell leukemia-lymphoma virus. Blood 1984, 64, 967–974. [Google Scholar] [CrossRef]
- Ehrlich, G.D.; Poiesz, B.J. Clinical and molecular parameters of HTLV-I infection. Clin. Lab. Med. 1988, 8, 65–84. [Google Scholar] [CrossRef]
- Geroldi, D.; Arico, M.; Plebani, A.; Maccario, R.; Notarangelo, L.D.; Duse, M.; Rey, F.; Barre Sinoussi, F.; Chermann, J.C.; Burgio, G.R. Western blot technique in the serological evaluation of three LAV/HTLV III-infected Italian families. Infection 1986, 14, 60–63. [Google Scholar] [CrossRef]
- Stroun, J.; Graf, I.; Frei, P.C. Detection of anti-HTLV-III/LAV antibody by enzyme-linked immunosorbent assay in high-risk individuals in Switzerland 1974–1985. Eur. J. Clin. Microbiol. 1985, 4, 583–586. [Google Scholar] [CrossRef]
- Burbelo, P.D.; Meoli, E.; Leahy, H.P.; Graham, J.; Yao, K.; Oh, U.; Janik, J.E.; Mahieux, R.; Kashanchi, F.; Iadarola, M.J.; et al. Anti-HTLV antibody profiling reveals an antibody signature for HTLV-I-associated myelopathy/tropical spastic paraparesis (HAM/TSP). Retrovirology 2008, 5, 96. [Google Scholar] [CrossRef]
- Osame, M.; Izumo, S.; Igata, A.; Matsumoto, M.; Matsumoto, T.; Sonoda, S.; Tara, M.; Shibata, Y. Blood transfusion and HTLV-I associated myelopathy. Lancet 1986, 2, 104–105. [Google Scholar] [CrossRef]
- Osame, M.; Janssen, R.; Kubota, H.; Nishitani, H.; Igata, A.; Nagataki, S.; Mori, M.; Goto, I.; Shimabukuro, H.; Khabbaz, R.; et al. Nationwide survey of HTLV-I-associated myelopathy in Japan: association with blood transfusion. Ann. Neurol. 1990, 28, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Soldan, S.S.; Graf, M.D.; Waziri, A.; Flerlage, A.N.; Robinson, S.M.; Kawanishi, T.; Leist, T.P.; Lehky, T.J.; Levin, M.C.; Jacobson, S. HTLV-I/II seroindeterminate Western blot reactivity in a cohort of patients with neurological disease. J. Infect. Dis. 1999, 180, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Varma, M.; Rudolph, D.L.; Knuchel, M.; Switzer, W.M.; Hadlock, K.G.; Velligan, M.; Chan, L.; Foung, S.K.; Lal, R.B. Enhanced specificity of truncated transmembrane protein for serologic confirmation of human T-cell lymphotropic virus type 1 (HTLV-1) and HTLV-2 infections by western blot (immunoblot) assay containing recombinant envelope glycoproteins. J. Clin. Microbiol. 1995, 33, 3239–3244. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Learn, G.H.; Rudicell, R.S.; Robertson, J.D.; Keele, B.F.; Ndjango, J.B.; Sanz, C.M.; Morgan, D.B.; Locatelli, S.; et al. Origin of the human malaria parasite Plasmodium falciparum in gorillas. Nature 2010, 467, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Rouet, F.; Meertens, L.; Courouble, G.; Herrmann-Storck, C.; Pabingui, R.; Chancerel, B.; Abid, A.; Strobel, M.; Mauclere, P.; Gessain, A. Serological, epidemiological, and molecular differences between human T-cell lymphotropic virus Type 1 (HTLV-1)-seropositive healthy carriers and persons with HTLV-I Gag indeterminate Western blot patterns from the Caribbean. J. Clin. Microbiol. 2001, 39, 1247–1253. [Google Scholar] [CrossRef]
- Khabbaz, R.F.; Heneine, W.; Grindon, A.; Hartley, T.M.; Shulman, G.; Kaplan, J. Indeterminate HTLV serologic results in U.S. blood donors: Are they due to HTLV-I or HTLV-II? J. Acquir. Immune Defic. Syndr. 1992, 5, 400–404. [Google Scholar] [PubMed]
- Zanjani, D.S.; Shahabi, M.; Talaei, N.; Afzalaghaee, M.; Tehranian, F.; Bazargani, R. Molecular analysis of human T cell lymphotropic virus type 1 and 2 (HTLV-1/2) seroindeterminate blood donors from Northeast Iran: Evidence of proviral tax, env, and gag sequences. AIDS Res. Hum. Retroviruses 2011, 27, 131–135. [Google Scholar] [CrossRef]
- Lu, S.C.; Chen, B.H. Seroindeterminate HTLV-1 prevalence and characteristics in blood donors in Taiwan. Int. J. Hematol. 2003, 77, 412–413. [Google Scholar] [CrossRef]
- Costa, J.M.; Segurado, A.C. Molecular evidence of human T-cell lymphotropic virus types 1 and 2 (HTLV-1 and HTLV-2) infections in HTLV seroindeterminate individuals from Sao Paulo, Brazil. J. Clin. Virol. 2009, 44, 185–189. [Google Scholar] [CrossRef]
- Busch, M.P.; Switzer, W.M.; Murphy, E.L.; Thomson, R.; Heneine, W. Absence of evidence of infection with divergent primate T-lymphotropic viruses in United States blood donors who have seroindeterminate HTLV test results. Transfusion 2000, 40, 443–449. [Google Scholar] [CrossRef]
- Garin, B.; Gosselin, S.; de The, G.; Gessain, A. HTLV-I/II infection in a high viral endemic area of Zaire, Central Africa: Comparative evaluation of serology, PCR, and significance of indeterminate western blot pattern. J. Med. Virol. 1994, 44, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Waziri, A.; Soldan, S.S.; Graf, M.D.; Nagle, J.; Jacobson, S. Characterization and sequencing of prototypic human T-lymphotropic virus type 1 (HTLV-1) from an HTLV-1/2 seroindeterminate patient. J. Virol. 2000, 74, 2178–2185. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Santos-Fortuna, E.; Azevedo, R.S.; Caterino-de-Araujo, A. Serological patterns and temporal trends of HTLV-1/2 infection in high-risk populations attending Public Health Units in Sao Paulo, Brazil. J. Clin. Virol. 2008, 42, 149–155. [Google Scholar] [CrossRef]
- Mangano, A.M.; Remesar, M.; del Pozo, A.; Sen, L. Human T lymphotropic virus types I and II proviral sequences in Argentinian blood donors with indeterminate Western blot patterns. J. Med. Virol. 2004, 74, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Hayes, C.G.; Burans, J.P.; Oberst, R.B. Antibodies to human T lymphotropic virus type I in a population from the Philippines: Evidence for cross-reactivity with Plasmodium falciparum. J. Infect. Dis. 1991, 163, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Lal, R.B.; Rudolph, D.; Alpers, M.P.; Sulzer, A.J.; Shi, Y.P.; Lal, A.A. Immunologic cross-reactivity between structural proteins of human T-cell lymphotropic virus type I and the blood stage of Plasmodium falciparum. Clin. Diagn. Lab. Immunol. 1994, 1, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Tsao, K.C.; Chen, G.W.; Huang, C.G.; Huang, Y.L.; Lin, J.Y.; Mok, C.K.; Sun, C.F.; Shih, S.R. False positive antibody results against human T-cell lymphotropic virus in patients with severe acute respiratory syndrome. J. Med. Virol. 2005, 77, 331–336. [Google Scholar] [CrossRef]
- Vandamme, A.M.; Van Laethem, K.; Liu, H.F.; Van Brussel, M.; Delaporte, E.; de Castro Costa, C.M.; Fleischer, C.; Taylor, G.; Bertazzoni, U.; Desmyter, J.; et al. Use of a generic polymerase chain reaction assay detecting human T-lymphotropic virus (HTLV) types I, II and divergent simian strains in the evaluation of individuals with indeterminate HTLV serology. J. Med. Virol. 1997, 52, 1–7. [Google Scholar] [CrossRef]
- Mauclere, P.; Le Hesran, J.Y.; Mahieux, R.; Salla, R.; Mfoupouendoun, J.; Abada, E.T.; Millan, J.; de The, G.; Gessain, A. Demographic, ethnic, and geographic differences between human T cell lymphotropic virus (HTLV) type I-seropositive carriers and persons with HTLV-I Gag-indeterminate Western blots in Central Africa. J. Infect. Dis. 1997, 176, 505–509. [Google Scholar] [CrossRef]
- Banki, K.; Maceda, J.; Hurley, E.; Ablonczy, E.; Mattson, D.H.; Szegedy, L.; Hung, C.; Perl, A. Human T-cell lymphotropic virus (HTLV)-related endogenous sequence, HRES-1, encodes a 28-kDa protein: A possible autoantigen for HTLV-I gag-reactive autoantibodies. Proc. Natl. Acad. Sci. U. S. A. 1992, 89, 1939–1943. [Google Scholar] [CrossRef]
- Bagossi, P.; Bander, P.; Bozoki, B.; Tozser, J. Discovery and significance of new human T-lymphotropic viruses: HTLV-3 and HTLV-4. Expert Rev. Anti-Infect. Ther. 2009, 7, 1235–1249. [Google Scholar] [CrossRef] [PubMed]
- Mahieux, R.; Gessain, A. The human HTLV-3 and HTLV-4 retroviruses: New members of the HTLV family. Pathol. Biol. (Paris) 2009, 57, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Hisada, M.; Maloney, E.; Yamano, Y.; Hanchard, B.; Wilks, R.; Rios, M.; Jacobson, S. Human T lymphotropic virus types I and II western blot seroindeterminate status and its association with exposure to prototype HTLV-I. J. Infect. Dis. 2006, 193, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Abbaszadegan, M.R.; Gholamin, M.; Tabatabaee, A.; Farid, R.; Houshmand, M.; Abbaszadegan, M. Prevalence of human T-lymphotropic virus type 1 among blood donors from Mashhad, Iran. J. Clin. Microbiol. 2003, 41, 2593–2595. [Google Scholar] [CrossRef]
- Lu, S.C.; Kao, C.L.; Chin, L.T.; Chen, J.W.; Yang, C.M.; Chang, J.H.; Hsu, S.C.; Chang, A.C.; Chen, B.H. Seroprevalence and demographic characteristics of HTLV-I among blood donors in Taiwan: 1996–1999. Int. J. Hematol. 2001, 74, 333–337. [Google Scholar] [CrossRef]
- Lal, R.B.; Rudolph, D.L.; Coligan, J.E.; Brodine, S.K.; Roberts, C.R. Failure to detect evidence of human T-lymphotropic virus (HTLV) type I and type II in blood donors with isolated gag antibodies to HTLV-I/II. Blood 1992, 80, 544–550. [Google Scholar] [CrossRef]
- Martins, M.L.; Santos, A.C.; Namen-Lopes, M.S.; Barbosa-Stancioli, E.F.; Utsch, D.G.; Carneiro-Proietti, A.B. Long-term serological follow-up of blood donors with an HTLV-indeterminate western blot: Antibody profile of seroconverters and individuals with false reactions. J. Med. Virol. 2010, 82, 1746–1753. [Google Scholar] [CrossRef]
- Caterino-de-Araujo, A.; de los Santos-Fortuna, E.; Meleiro, M.C.; Suleiman, J.; Calabro, M.L.; Favero, A.; De Rossi, A.; Chieco-Bianchi, L. Sensitivity of two enzyme-linked immunosorbent assay tests in relation to western blot in detecting human T-cell lymphotropic virus types I and II infection among HIV-1 infected patients from Sao Paulo, Brazil. Diagn. Microbiol. Infect. Dis. 1998, 30, 173–182. [Google Scholar] [CrossRef]
- Dezzutti, C.S.; Patel, P.P.; Owen, S.M.; Switzer, W.M.; Meshulam, J.; Lal, R.B. Sensitivity and specificity of a DNA polymerase chain reaction nonisotopic-based detection method for the confirmation of infection with human T-lymphotropic virus types I and II. Clin. Diagn. Virol. 1996, 6, 103–110. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Country [Reference] | Number Screened | HTLV-I EIA Reactive | HTLV-I WB + (% of Total EIA Reactive) | HTLV-II WB + (% of Total EIA Reactive) | WB Seroindeterminate (% of Total EIA Reactive) | * HTLV-I PCR + (% of WB Serondeterminates) |
|---|---|---|---|---|---|---|
| Argentina [17] | 86,238 | 0.17% | 20% | 8.0% | 61% | 13.8% |
| Brazil [19] | 351,639 | 1.22% | 24% | Not Shown | 52.2% | 9.2% |
| Iran [17] | 79,687 | 6.4% | 50.7 % | 3.1% | 25.4% | 12.5% |
| Taiwan [35] | 1,122,879 | 0.1% | 41% | 1.0% | 22% | Not Shown |
| US [36] | 267,650 | 0.89% | 3% | Not Shown | 16% | 0.00% |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Abrams, A.; Akahata, Y.; Jacobson, S. The Prevalence and Significance of HTLV-I/II Seroindeterminate Western Blot Patterns. Viruses 2011, 3, 1320-1331. https://doi.org/10.3390/v3081320
Abrams A, Akahata Y, Jacobson S. The Prevalence and Significance of HTLV-I/II Seroindeterminate Western Blot Patterns. Viruses. 2011; 3(8):1320-1331. https://doi.org/10.3390/v3081320
Chicago/Turabian StyleAbrams, Anna, Yoshimi Akahata, and Steven Jacobson. 2011. "The Prevalence and Significance of HTLV-I/II Seroindeterminate Western Blot Patterns" Viruses 3, no. 8: 1320-1331. https://doi.org/10.3390/v3081320
APA StyleAbrams, A., Akahata, Y., & Jacobson, S. (2011). The Prevalence and Significance of HTLV-I/II Seroindeterminate Western Blot Patterns. Viruses, 3(8), 1320-1331. https://doi.org/10.3390/v3081320