The Pneumonia Virus of Mice (PVM) Model of Acute Respiratory Infection

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

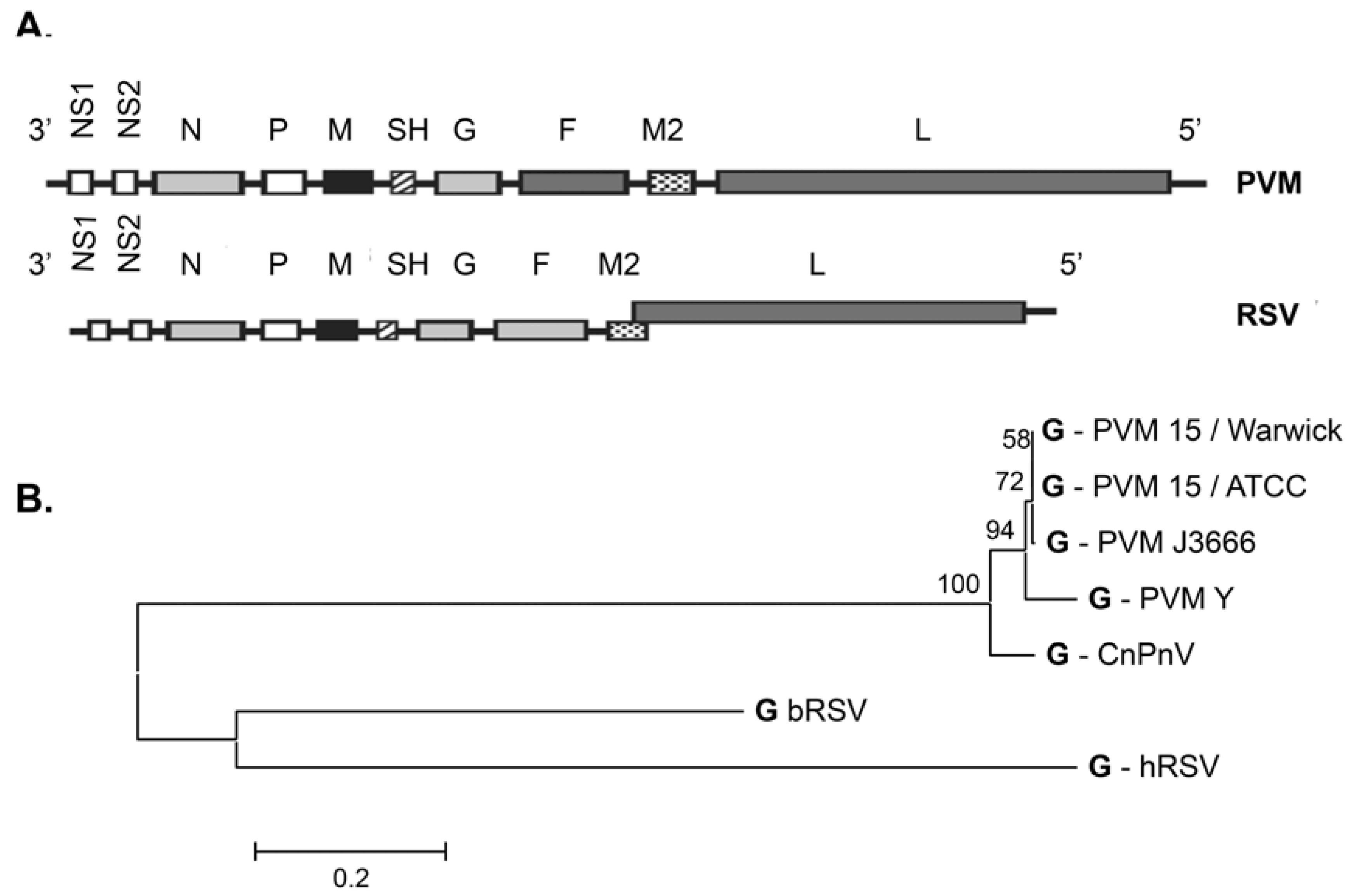
2. The PVM Model of Acute Respiratory Infection
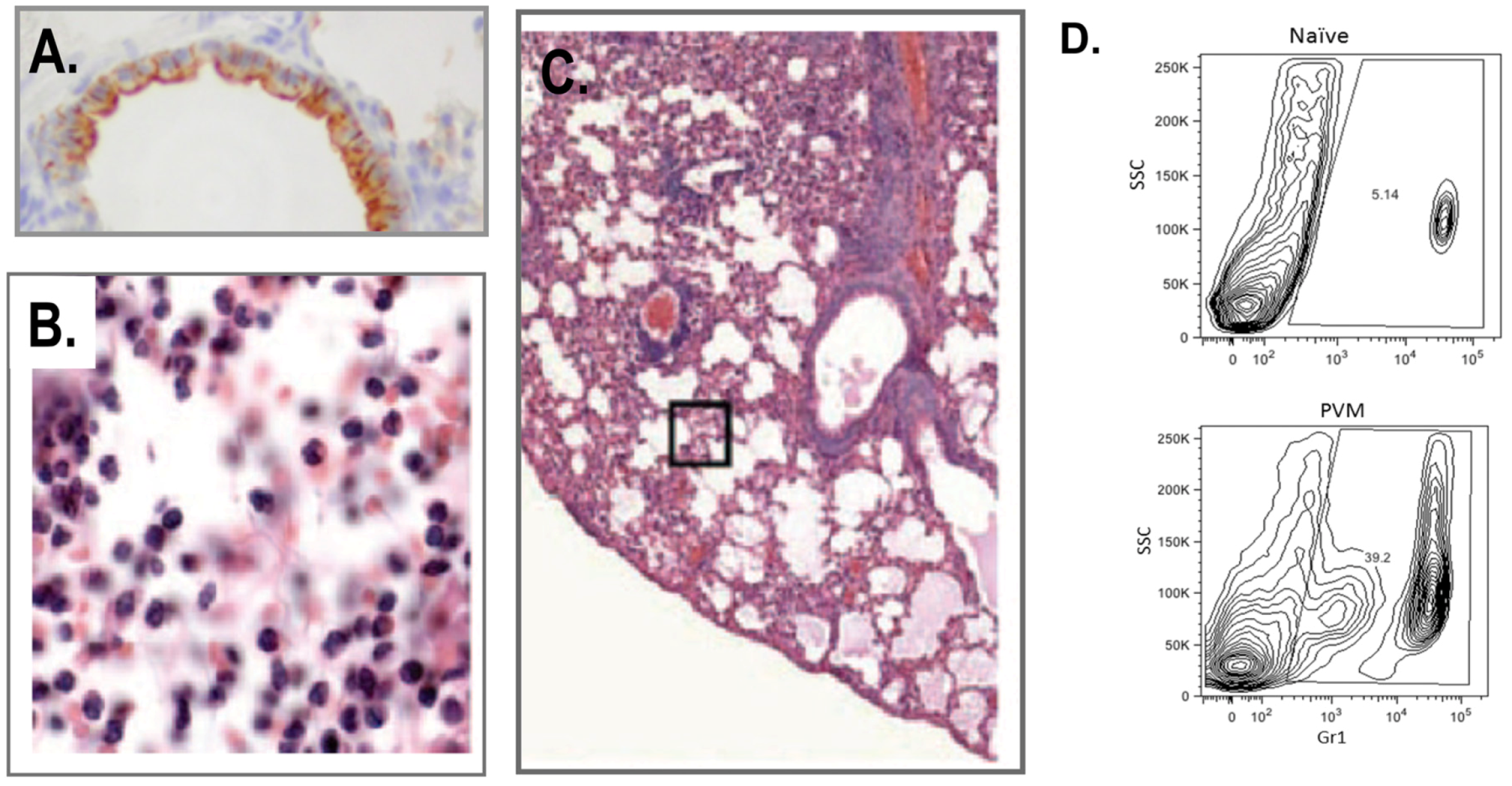
3. Host Immune Response to PVM Infection
3.1. Neutrophils and Eosinophils
3.2. T Lymphocytes

3.3. Macrophages
3.4. Toll-Like Receptors
3.5. Type I Interferons
4. Using the PVM Model to Explore Human Disease
4.1. Inflammation and Acute Infection
4.2. Asthma and Allergic Airway Disease
4.3. Vaccines
4.4. Heterologous Immunity

5. Conclusions
Acknowledgements
Conflict of Interest
References and Notes
- Easton, A.J.; Domachowske, J.B.; Rosenberg, H.F. Animal pneumoviruses: Molecular genetics and pathogenesis. Clin. Microbiol. Rev. 2004, 17, 390–412. [Google Scholar] [CrossRef]
- Horsfall, F.L.; Hahn, R.G. A latent virus in normal mice capable of producing pneumonia in its natural host. J. Exp. Med. 1940, 71, 391–408. [Google Scholar] [CrossRef]
- Miyata, H.; Kishikawa, M.; Kondo, H.; Kai, C.; Watanabe, Y.; Ohsawa, K.; Sato, H. New isolates of pneumonia virus of mice (PVM) from Japanese rat colonies and their characterization. Exp. Anim. 1995, 44, 95–104. [Google Scholar] [CrossRef]
- Zenner, L.; Regnault, J.P. Ten-year long monitoring of laboratory mouse and rat colonies in French facilities: A retrospective study. Lab. Anim. 2000, 34, 76–83. [Google Scholar] [CrossRef]
- Liang, C.T.; Shih, A.; Chang, Y.H.; Liu, C.W.; Lee, Y.T.; Hsieh, W.C.; Huang, Y.L.; Huang, W.T.; Kuang, C.H.; Lee, K.H.; et al. Microbial contaminations of laboratory mice and rats in Taiwan from 2004 to 2007. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 381–386. [Google Scholar]
- Kaplan, C.; Healing, T.D.; Evans, N.; Healing, L.; Prior, A. Evidence of infection by viruses in small British field rodents. J. Hyg. 1980, 84, 285–294. [Google Scholar] [CrossRef]
- Smith, A.L.; Singleton, G.R.; Hansen, G.M.; Shellam, G. A serologic survey for viruses and Mycoplasma pulmonis among wild house mice (Mus domesticus) in southeastern Australia. J. Wildlife Dis. 1993, 29, 219–229. [Google Scholar]
- Pringle, C.R.; Eglin, R.P. Murine pneumonia virus: Seroepidemiological evidence of widespread human infection. J. Gen. Virol. 1986, 67, 975–982. [Google Scholar] [CrossRef]
- Brock, L.G.; Karron, R.A.; Krempl, C.D.; Collins, P.L.; Buchholz, U.J. Evaluation of pneumonia virus of mice as a possible human pathogen. J. Virol. 2012, 86, 5829–5843. [Google Scholar]
- Renshaw, R.; Laverack, M.; Zylich, N.; Glaser, A.; Dubovi, E. Genomic analysis of a pneumovirus isolated from dogs with acute respiratory disease. Vet. Microbiol. 2011, 150, 88–95. [Google Scholar] [CrossRef]
- Renshaw, R.W.; Zylich, N.C.; Laverack, M.A.; Glaser, A.L.; Dubovi, E.J. Pneumovirus in dogs with acute respiratory disease. Emerg. Infect. Dis. 2010, 16, 993–995. [Google Scholar] [CrossRef]
- Percopo, C.M.; Dubovi, E.J.; Renshaw, R.W.; Dyer, K.D.; Domachowske, J.B.; Rosenberg, H.F. Canine pneumovirus replicates in mouse lung tissue and elicits inflammatory pathology. Virology 2011, 416, 26–31. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Horsfall, F.L., Jr. Concurrent infection with influenza virus and mumps virus or pneumonia virus of mice as bearing on the inhibition of virus multiplication by bacterial polysaccharides. J. Exp. Med. 1949, 89, 37–52. [Google Scholar] [CrossRef]
- Ginsberg, H.S.; Horsfall, F.L., Jr. Therapy of infection with pneumonia virus of mice (PVM); effect of a polysaccharide on the multiplication cycles of the virus and on the course of the viral pneumonia. J. Exp. Med. 1951, 93, 161–171. [Google Scholar] [CrossRef]
- Horsfall, F.L., Jr.; Ginsberg, H.S. The dependence of the pathological lesion upon the multiplication of pneumonia virus of mice (PVM); kinetic relation between the degree of viral multiplication and the extent of pneumonia. J. Exp. Med. 1951, 93, 139–150. [Google Scholar] [CrossRef]
- Thorpe, L.C.; Easton, A.J. Genome sequence of the non-pathogenic strain 15 of pneumonia virus of mice and comparison with the genome of the pathogenic strain J3666. J. Gen. Virol. 2005, 86, 159–169. [Google Scholar] [CrossRef]
- Ellis, J.A.; Martin, B.V.; Waldner, C.; Dyer, K.D.; Domachowske, J.B.; Rosenberg, H.F. Mucosal inoculation with an attenuated mouse pneumovirus strain protects against virulent challenge in wild type and interferon-gamma receptor deficient mice. Vaccine 2007, 25, 1085–1095. [Google Scholar] [CrossRef]
- Easton, A.J. Personal communication, University of Warwick: Coventry, UK, 1997.
- Ahmadian, G.; Chambers, P.; Easton, A.J. Detection and characterization of proteins encoded by the second ORF of the M2 gene of pneumoviruses. J. Gen. Virol. 1999, 80, 2011–2016. [Google Scholar]
- Barr, J.; Easton, A.J. Characterisation of the interaction between the nucleoprotein and phosphoprotein of pneumonia virus of mice. Virus Res. 1995, 39, 221–235. [Google Scholar] [CrossRef]
- Chambers, P.; Pringle, C.R.; Easton, A.J. Sequence analysis of the gene encoding the fusion glycoprotein of pneumonia virus of mice suggests possible conserved secondary structure elements in paramyxovirus fusion glycoproteins. J. Gen. Virol. 1992, 73, 1717–1724. [Google Scholar] [CrossRef]
- Easton, A.J.; Chambers, P. Nucleotide sequence of the genes encoding the matrix and small hydrophobic proteins of pneumonia virus of mice. Virus Res. 1997, 48, 27–33. [Google Scholar] [CrossRef]
- Krempl, C.D.; Collins, P.L. Reevaluation of the virulence of prototypic strain 15 of pneumonia virus of mice. J. Virol. 2004, 78, 13362–13365. [Google Scholar] [CrossRef]
- Krempl, C.D.; Wnekowicz, A.; Lamirande, E.W.; Nayebagha, G.; Collins, P.L.; Buchholz, U.J. Identification of a novel virulence factor in recombinant pneumonia virus of mice. J. Virol. 2007, 81, 9490–9501. [Google Scholar] [CrossRef]
- Anh, D.B.; Faisca, P.; Desmecht, D.J. Differential resistance/susceptibility patterns to pneumovirus infection among inbred mouse strains. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 291, L426–L435. [Google Scholar] [CrossRef]
- Glineur, S.; Tran Anh, D.B.; Sarlet, M.; Michaux, C.; Desmecht, D. Characterization of the resistance of SJL/J mice to pneumonia virus of mice, a model for infantile bronchiolitis due to a respiratory syncytial virus. PLoS One 2012, 7, e44581. [Google Scholar]
- Weir, E.C.; Brownstein, D.G.; Smith, A.L.; Johnson, E.A. Respiratory disease and wasting in athymic mice infected with pneumonia virus of mice. Lab. Anim. Sci. 1988, 38, 133–137. [Google Scholar]
- Roths, J.B.; Smith, A.L.; Sidman, C.L. Lethal exacerbation of Pneumocystis carinii pneumonia in severe combined immunodeficiency mice after infection by pneumonia virus of mice. J. Exp. Med. 1993, 177, 1193–1198. [Google Scholar] [CrossRef]
- Compton, C. Personal communication, Yale University: New Haven, CT, USA, 2012.
- Bem, R.A.; Domachowske, J.B.; Rosenberg, H.F. Animal models of human respiratory syncytial virus disease. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 301, L148–L156. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Domachowske, J.B. Pneumonia virus of mice: Severe respiratory infection in a natural host. Immunol. Lett. 2008, 118, 6–12. [Google Scholar] [CrossRef]
- Domachowske, J.B.; Bonville, C.A.; Dyer, K.D.; Easton, A.J.; Rosenberg, H.F. Pulmonary eosinophilia and production of MIP-1alpha are prominent responses to infection with pneumonia virus of mice. Cell Immunol. 2000, 200, 98–104. [Google Scholar] [CrossRef]
- Domachowske, J.B.; Bonville, C.A.; Gao, J.L.; Murphy, P.M.; Easton, A.J.; Rosenberg, H.F. The chemokine macrophage-inflammatory protein-1 alpha and its receptor CCR1 control pulmonary inflammation and antiviral host defense in paramyxovirus infection. J. Immunol. 2000, 165, 2677–2682. [Google Scholar]
- Bonville, C.A.; Bennett, N.J.; Koehnlein, M.; Haines, D.M.; Ellis, J.A.; DelVecchio, A.M.; Rosenberg, H.F.; Domachowske, J.B. Respiratory dysfunction and proinflammatory chemokines in the pneumonia virus of mice (PVM) model of viral bronchiolitis. Virology 2006, 349, 87–95. [Google Scholar] [CrossRef]
- Welliver, T.P.; Garofalo, R.P.; Hosakote, Y.; Hintz, K.H.; Avendano, L.; Sanchez, K.; Velozo, L.; Jafri, H.; Chavez-Bueno, S.; Ogra, P.L.; et al. Severe human lower respiratory tract illness caused by respiratory syncytial virus and influenza virus is characterized by the absence of pulmonary cytotoxic lymphocyte responses. J. Infect. Dis. 2007, 195, 1126–1136. [Google Scholar] [CrossRef]
- Domachowske, J.B.; Bonville, C.A.; Rosenberg, H.F. Animal models for studying respiratory syncytial virus infection and its long term effects on lung function. Pediatr. Infect. Dis. J. 2004, 23, S228–S234. [Google Scholar] [CrossRef]
- Bonville, C.A.; Bennett, N.J.; Percopo, C.M.; Branigan, P.J.; Del Vecchio, A.M.; Rosenberg, H.F.; Domachowske, J.B. Diminished inflammatory responses to natural pneumovirus infection among older mice. Virology 2007, 368, 182–190. [Google Scholar] [CrossRef]
- Bonville, C.A.; Ptaschinski, C.; Percopo, C.M.; Rosenberg, H.F.; Domachowske, J.B. Inflammatory responses to acute pneumovirus infection in neonatal mice. Virol. J. 2010, 7, 320. [Google Scholar] [CrossRef]
- Cook, D.N.; Beck, M.A.; Coffman, T.M.; Kirby, S.L.; Sheridan, J.F.; Pragnell, I.B.; Smithies, O. Requirement of MIP-1 alpha for an inflammatory response to viral infection. Science 1995, 269, 1583–1585. [Google Scholar]
- Bonville, C.A.; Easton, A.J.; Rosenberg, H.F.; Domachowske, J.B. Altered pathogenesis of severe pneumovirus infection in response to combined antiviral and specific immunomodulatory agents. J. Virol. 2003, 77, 1237–1244. [Google Scholar] [CrossRef]
- Bonville, C.A.; Lau, V.K.; DeLeon, J.M.; Gao, J.L.; Easton, A.J.; Rosenberg, H.F.; Domachowske, J.B. Functional antagonism of chemokine receptor CCR1 reduces mortality in acute pneumovirus infection in vivo. J. Virol. 2004, 78, 7984–7989. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Dyer, K.D.; Domachowske, J.B. Eosinophils and their interactions with respiratory virus pathogens. Immunol. Res. 2009, 43, 128–137. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Dyer, K.D.; Domachowske, J.B. Respiratory viruses and eosinophils: Exploring the connections. Antivir. Res. 2009, 83, 1–9. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Dyer, K.D.; Foster, P.S. Eosinophils: Changing perspectives in health and disease. Nat. Rev. Immunol. 2012, in press. [Google Scholar]
- Adamko, D.J.; Yost, B.L.; Gleich, G.J.; Fryer, A.D.; Jacoby, D.B. Ovalbumin sensitization changes the inflammatory response to subsequent parainfluenza infection. Eosinophils mediate airway hyperresponsiveness, m(2) muscarinic receptor dysfunction, and antiviral effects. J. Exp. Med. 1999, 190, 1465–1478. [Google Scholar] [CrossRef]
- Phipps, S.; Lam, C.E.; Mahalingam, S.; Newhouse, M.; Ramirez, R.; Rosenberg, H.F.; Foster, P.S.; Matthaei, K.I. Eosinophils contribute to innate antiviral immunity and promote clearance of respiratory syncytial virus. Blood 2007, 110, 1578–1586. [Google Scholar] [CrossRef]
- Percopo, C.M.; Dyer, K.D.; Ochkur, S.I.; Lee, J.J.; Domachowske, J.B.; Rosenberg, H.F. Activated eosinophils protect against lethal respiratory virus infection. J. Immunol. 2013. to be submitted for publication. [Google Scholar]
- Dyer, K.D.; Percopo, C.M.; Fischer, E.R.; Gabryszewski, S.J.; Rosenberg, H.F. Pneumoviruses infect eosinophils and elicit MyD88-dependent release of chemoattractant cytokines and interleukin-6. Blood 2009, 114, 2649–2656. [Google Scholar] [CrossRef]
- Frey, S.; Krempl, C.D.; Schmitt-Graff, A.; Ehl, S. Role of T cells in virus control and disease after infection with pneumonia virus of mice. J. Virol. 2008, 82, 11619–11627. [Google Scholar] [CrossRef]
- Claassen, E.A.; van der Kant, P.A.; Rychnavska, Z.S.; van Bleek, G.M.; Easton, A.J.; van der Most, R.G. Activation and inactivation of antiviral CD8 T cell responses during murine pneumovirus infection. J. Immunol. 2005, 175, 6597–6604. [Google Scholar]
- Chang, J.; Braciale, T.J. Respiratory syncytial virus infection suppresses lung CD8+ T-cell effector activity and peripheral CD8+ T-cell memory in the respiratory tract. Nat. Med. 2002, 8, 54–60. [Google Scholar] [CrossRef]
- Claassen, E.A.; van Bleek, G.M.; Rychnavska, Z.S.; de Groot, R.J.; Hensen, E.J.; Tijhaar, E.J.; van Eden, W.; van der Most, R.G. Identification of a CD4 T cell epitope in the pneumonia virus of mice glycoprotein and characterization of its role in protective immunity. Virology 2007, 368, 17–25. [Google Scholar] [CrossRef]
- Spolski, R.; Wang, L.; Wan, C.K.; Bonville, C.A.; Domachowske, J.B.; Kim, H.P.; Yu, Z.; Leonard, W.J. IL-21 promotes the pathologic immune response to pneumovirus infection. J. Immunol. 2012, 188, 1924–1932. [Google Scholar] [CrossRef]
- Garvey, T.L.; Dyer, K.D.; Ellis, J.A.; Bonville, C.A.; Foster, B.; Prussin, C.; Easton, A.J.; Domachowske, J.B.; Rosenberg, H.F. Inflammatory responses to pneumovirus infection in IFN-alpha beta R gene-deleted mice. J. Immunol. 2005, 175, 4735–4744. [Google Scholar]
- Rigaux, P.; Killoran, K.E.; Qiu, Z.; Rosenberg, H.F. Depletion of alveolar macrophages prolongs survival in response to acute pneumovirus infection. Virology 2012, 422, 338–345. [Google Scholar] [CrossRef]
- Pribul, P.K.; Harker, J.; Wang, B.; Wang, H.; Tregoning, J.S.; Schwarze, J.; Openshaw, P.J. Alveolar macrophages are a major determinant of early responses to viral lung infection but do not influence subsequent disease development. J. Virol. 2008, 82, 4441–4448. [Google Scholar] [CrossRef]
- Reed, J.L.; Brewah, Y.A.; Delaney, T.; Welliver, T.; Burwell, T.; Benjamin, E.; Kuta, E.; Kozhich, A.; McKinney, L.; Suzich, J.; et al. Macrophage impairment underlies airway occlusion in primary respiratory syncytial virus bronchiolitis. J. Infect. Dis. 2008, 198, 1783–1793. [Google Scholar] [CrossRef]
- Klein Klouwenberg, P.; Tan, L.; Werkman, W.; van Bleek, G.M.; Coenjaerts, F. The role of Toll-like receptors in regulating the immune response against respiratory syncytial virus. Crit. Rev. Immunol. 2009, 29, 531–550. [Google Scholar] [CrossRef]
- Kurt-Jones, E.A.; Popova, L.; Kwinn, L.; Haynes, L.M.; Jones, L.P.; Tripp, R.A.; Walsh, E.E.; Freeman, M.W.; Golenbock, D.T.; Anderson, L.J.; et al. Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat. Immunol. 2000, 1, 398–401. [Google Scholar]
- Ehl, S.; Bischoff, R.; Ostler, T.; Vallbracht, S.; Schulte-Monting, J.; Poltorak, A.; Freudenberg, M. The role of Toll-like receptor 4 versus interleukin-12 in immunity to respiratory syncytial virus. Eur. J. Immunol. 2004, 34, 1146–1153. [Google Scholar] [CrossRef]
- Faisca, P.; Tran Anh, D.B.; Thomas, A.; Desmecht, D. Suppression of pattern-recognition receptor TLR4 sensing does not alter lung responses to pneumovirus infection. Microbes Infect. 2006, 8, 621–627. [Google Scholar] [CrossRef]
- Marr, N.; Turvey, S.E. Role of human TLR4 in respiratory syncytial virus-induced NF-kappaB activation, viral entry and replication. Innate Immun. 2012, 18, 856–865. [Google Scholar] [CrossRef]
- Davidson, S.; Kaiko, G.; Loh, Z.; Lalwani, A.; Zhang, V.; Spann, K.; Foo, S.Y.; Hansbro, N.; Uematsu, S.; Akira, S.; et al. Plasmacytoid dendritic cells promote host defense against acute pneumovirus infection via the TLR7-MyD88-dependent signaling pathway. J. Immunol. 2011, 186, 5938–5948. [Google Scholar] [CrossRef]
- Fontana, J.M.; Bankamp, B.; Rota, P.A. Inhibition of interferon induction and signaling by paramyxoviruses. Immunol. Rev. 2008, 225, 46–67. [Google Scholar] [CrossRef]
- Bossert, B.; Conzelmann, K.K. Respiratory syncytial virus (RSV) nonstructural (NS) proteins as host range determinants: A chimeric bovine RSV with NS genes from human RSV is attenuated in interferon-competent bovine cells. J. Virol. 2002, 76, 4287–4293. [Google Scholar] [CrossRef]
- Ramaswamy, M.; Shi, L.; Monick, M.M.; Hunninghake, G.W.; Look, D.C. Specific inhibition of type I interferon signal transduction by respiratory syncytial virus. Am. J. Respir. Cell Mol. Biol. 2004, 30, 893–900. [Google Scholar]
- Schlender, J.; Hornung, V.; Finke, S.; Gunthner-Biller, M.; Marozin, S.; Brzozka, K.; Moghim, S.; Endres, S.; Hartmann, G.; Conzelmann, K.K. Inhibition of toll-like receptor 7- and 9-mediated alpha/beta interferon production in human plasmacytoid dendritic cells by respiratory syncytial virus and measles virus. J. Virol. 2005, 79, 5507–5515. [Google Scholar]
- Spann, K.M.; Tran, K.C.; Chi, B.; Rabin, R.L.; Collins, P.L. Suppression of the induction of alpha, beta, and lambda interferons by the NS1 and NS2 proteins of human respiratory syncytial virus in human epithelial cells and macrophages [corrected]. J. Virol. 2004, 78, 4363–4369. [Google Scholar] [CrossRef]
- Randall, T.D. Bronchus-associated lymphoid tissue (BALT) structure and function. Adv. Immunol. 2010, 107, 187–241. [Google Scholar] [CrossRef]
- Buchholz, U.J.; Ward, J.M.; Lamirande, E.W.; Heinze, B.; Krempl, C.D.; Collins, P.L. Deletion of nonstructural proteins NS1 and NS2 from pneumonia virus of mice attenuates viral replication and reduces pulmonary cytokine expression and disease. J. Virol. 2009, 83, 1969–1980. [Google Scholar] [CrossRef]
- Heinze, B.; Frey, S.; Mordstein, M.; Schmitt-Graff, A.; Ehl, S.; Buchholz, U.J.; Collins, P.L.; Staeheli, P.; Krempl, C.D. Both nonstructural proteins NS1 and NS2 of pneumonia virus of mice are inhibitors of the interferon type I and type III responses in vivo. J. Virol. 2011, 85, 4071–4084. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Domachowske, J.B. Inflammatory responses to respiratory syncytial virus (RSV) infection and the development of immunomodulatory pharmacotherapeutics. Curr. Med. Chem. 2012, 19, 1424–1431. [Google Scholar] [CrossRef]
- Reassessment of the indications for ribavirin therapy in respiratory syncytial virus infections. American Academy of Pediatrics Committee on Infectious Diseases. Pediatrics 1996, 97, 137–140.
- Van Woensel, J.B.; Kimpen, J.L.; Brand, P.L. Respiratory tract infections caused by respiratory syncytial virus in children. Diagnosis and treatment. Minerva Pediatr. 2001, 53, 99–106. [Google Scholar]
- Bonville, C.A.; Rosenberg, H.F.; Domachowske, J.B. Ribavirin and cysteinyl leukotriene-1 receptor blockade as treatment for severe bronchiolitis. Antivir. Res. 2006, 69, 53–59. [Google Scholar]
- Bondue, B.; Vosters, O.; de Nadai, P.; Glineur, S.; De Henau, O.; Luangsay, S.; van Gool, F.; Communi, D.; de Vuyst, P.; Desmecht, D.; et al. ChemR23 dampens lung inflammation and enhances anti-viral immunity in a mouse model of acute viral pneumonia. PLoS Pathog. 2011, 7, e1002358. [Google Scholar] [CrossRef]
- Buckingham, S.C.; Jafri, H.S.; Bush, A.J.; Carubelli, C.M.; Sheeran, P.; Hardy, R.D.; Ottolini, M.G.; Ramilo, O.; DeVincenzo, J.P. A randomized, double-blind, placebo-controlled trial of dexamethasone in severe respiratory syncytial virus (RSV) infection: Effects on RSV quantity and clinical outcome. J. Infect. Dis. 2002, 185, 1222–1228. [Google Scholar] [CrossRef]
- Patel, H.; Platt, R.; Lozano, J.M.; Wang, E.E. Glucocorticoids for acute viral bronchiolitis in infants and young children. Cochrane Database Syst. Rev. 2004, CD004878. [Google Scholar]
- Van Woensel, J.B.; Lutter, R.; Biezeveld, M.H.; Dekker, T.; Nijhuis, M.; van Aalderen, W.M.; Kuijpers, T.W. Effect of dexamethasone on tracheal viral load and interleukin-8 tracheal concentration in children with respiratory syncytial virus infection. Pediatr. Infect. Dis. J. 2003, 22, 721–726. [Google Scholar] [CrossRef]
- Van Woensel, J.B.; van Aalderen, W.M.; de Weerd, W.; Jansen, N.J.; van Gestel, J.P.; Markhorst, D.G.; van Vught, A.J.; Bos, A.P.; Kimpen, J.L. Dexamethasone for treatment of patients mechanically ventilated for lower respiratory tract infection caused by respiratory syncytial virus. Thorax 2003, 58, 383–387. [Google Scholar] [CrossRef]
- Domachowske, J.B.; Bonville, C.A.; Ali-Ahmad, D.; Dyer, K.D.; Easton, A.J.; Rosenberg, H.F. Glucocorticoid administration accelerates mortality of pneumovirus-infected mice. J. Infect. Dis. 2001, 184, 1518–1523. [Google Scholar] [CrossRef]
- Bem, R.A.; van Woensel, J.B.; Bos, A.P.; Koski, A.; Farnand, A.W.; Domachowske, J.B.; Rosenberg, H.F.; Martin, T.R.; Matute-Bello, G. Mechanical ventilation enhances lung inflammation and caspase activity in a model of mouse pneumovirus infection. Am. J. Physiol. Lung Cell Mol. Physiol. 2009, 296, L46–56. [Google Scholar]
- Van den Berg, E.; van Woensel, J.B.; Bos, A.P.; Bem, R.A.; Altemeier, W.A.; Gill, S.E.; Martin, T.R.; Matute-Bello, G. Role of the Fas/FasL system in a model of RSV infection in mechanically ventilated mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 301, L451–L460. [Google Scholar] [CrossRef]
- Lukacs, N.W.; Smit, J.; Lindell, D.; Schaller, M. Respiratory syncytial virus-induced pulmonary disease and exacerbation of allergic asthma. Contrib. Microbiol. 2007, 14, 68–82. [Google Scholar]
- Xepapadaki, P.; Papadopoulos, N.G. Viral infections and allergies. Immunobiology 2007, 212, 453–459. [Google Scholar] [CrossRef]
- Siegle, J.S.; Hansbro, N.; Dong, C.; Angkasekwinai, P.; Foster, P.S.; Kumar, R.K. Blocking induction of T helper type 2 responses prevents development of disease in a model of childhood asthma. Clin. Exp. Immunol. 2011, 165, 19–28. [Google Scholar] [CrossRef]
- Siegle, J.S.; Hansbro, N.; Herbert, C.; Rosenberg, H.F.; Domachowske, J.B.; Asquith, K.L.; Foster, P.S.; Kumar, R.K. Early-life viral infection and allergen exposure interact to induce an asthmatic phenotype in mice. Respir. Res. 2010, 11, 14. [Google Scholar] [CrossRef]
- Barends, M.; de Rond, L.G.; Dormans, J.; van Oosten, M.; Boelen, A.; Neijens, H.J.; Osterhaus, A.D.; Kimman, T.G. Respiratory syncytial virus, pneumonia virus of mice, and influenza A virus differently affect respiratory allergy in mice. Clin. Exp. Allergy 2004, 34, 488–496. [Google Scholar] [CrossRef]
- Kapikian, A.Z.; Mitchell, R.H.; Chanock, R.M.; Shvedoff, R.A.; Stewart, C.E. An epidemiologic study of altered clinical reactivity to respiratory syncytial (RS) virus infection in children previously vaccinated with an inactivated RS virus vaccine. Am. J. Epidemiol. 1969, 89, 405–421. [Google Scholar]
- Castilow, E.M.; Olson, M.R.; Meyerholz, D.K.; Varga, S.M. Differential role of gamma interferon in inhibiting pulmonary eosinophilia and exacerbating systemic disease in fusion protein-immunized mice undergoing challenge infection with respiratory syncytial virus. J. Virol. 2008, 82, 2196–2207. [Google Scholar] [CrossRef]
- Castilow, E.M.; Olson, M.R.; Varga, S.M. Understanding respiratory syncytial virus (RSV) vaccine-enhanced disease. Immunol. Res. 2007, 39, 225–239. [Google Scholar] [CrossRef]
- Percopo, C.M.; Qiu, Z.J.; Phipps, S.; Foster, P.S.; Domachowske, J.B.; Rosenberg, H.F. Pulmonary eosinophils and their role in immunopathologic responses to formalin-inactivated pneumonia virus of mice. J. Immunol. 2009, 183, 604–612. [Google Scholar] [CrossRef]
- Hurwitz, J.L. Respiratory syncytial virus vaccine development. Expert Rev. Vaccines. 2011, 10, 1415–1433. [Google Scholar] [CrossRef]
- Jones, B.G.; Sealy, R.E.; Rudraraju, R.; Traina-Dorge, V.L.; Finneyfrock, B.; Cook, A.; Takimoto, T.; Portner, A.; Hurwitz, J.L. Sendai virus-based RSV vaccine protects African green monkeys from RSV infection. Vaccine 2012, 30, 959–968. [Google Scholar]
- Van Helden, M.J.; van Kooten, P.J.; Bekker, C.P.; Grone, A.; Topham, D.J.; Easton, A.J.; Boog, C.J.; Busch, D.H.; Zaiss, D.M.; Sijts, A.J. Pre-existing virus-specific CD8(+) T-cells provide protection against pneumovirus-induced disease in mice. Vaccine 2012, 30, 6382–6388. [Google Scholar]
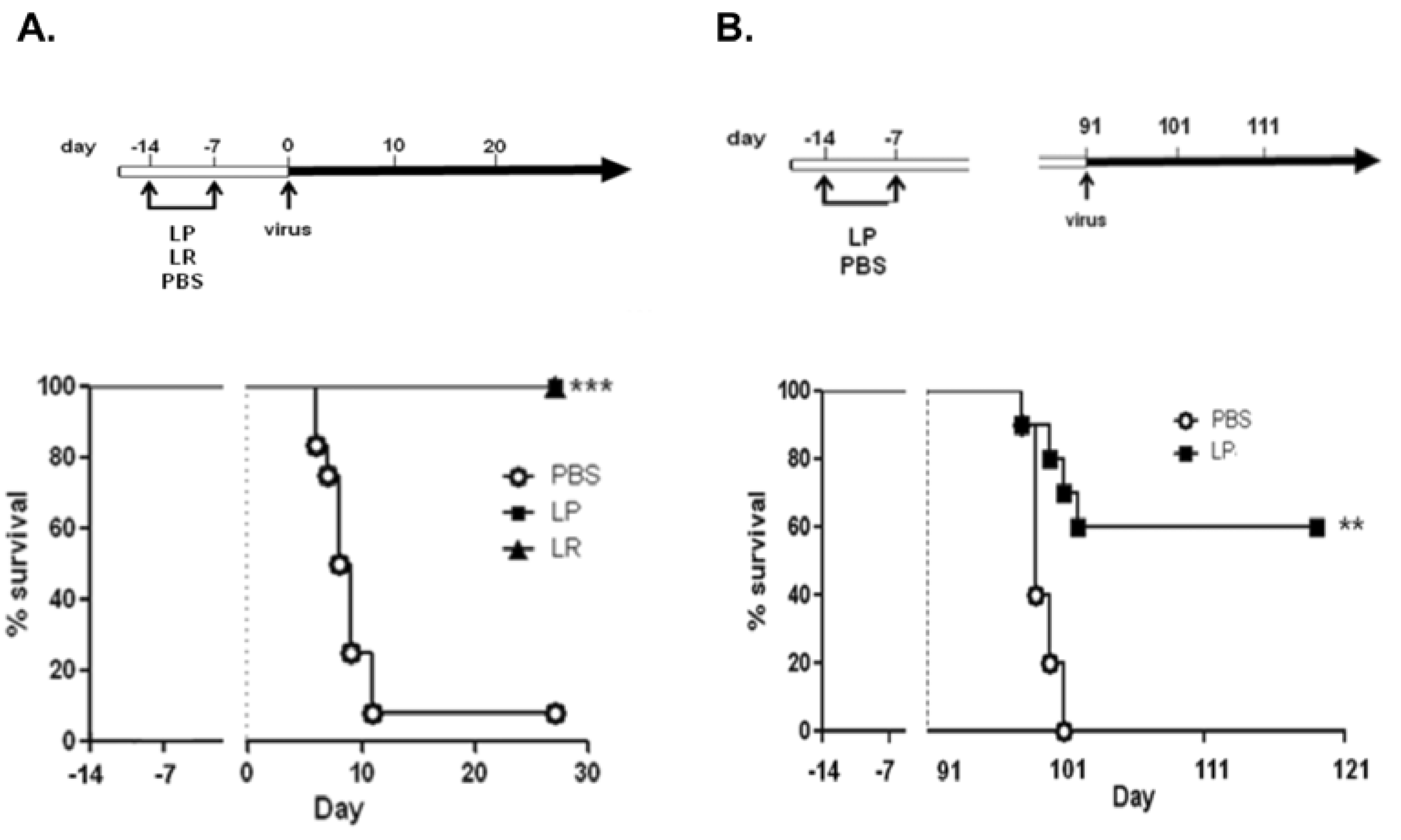
- Gabryszewski, S.J.; Bachar, O.; Dyer, K.D.; Percopo, C.M.; Killoran, K.E.; Domachowske, J.B.; Rosenberg, H.F. Lactobacillus-mediated priming of the respiratory mucosa protects against lethal pneumovirus infection. J. Immunol. 2011, 186, 1151–1161. [Google Scholar] [CrossRef]
- Didierlaurent, A.; Goulding, J.; Hussell, T. The impact of successive infections on the lung microenvironment. Immunology 2007, 122, 457–465. [Google Scholar] [CrossRef]
- Goulding, J.; Snelgrove, R.; Saldana, J.; Didierlaurent, A.; Cavanagh, M.; Gwyer, E.; Wales, J.; Wissinger, E.L.; Hussell, T. Respiratory infections: Do we ever recover? Proc. Am. Thorac. Soc. 2007, 4, 618–625. [Google Scholar] [CrossRef]
- Hussell, T.; Cavanagh, M.M. The innate immune rheostat: Influence on lung inflammatory disease and secondary bacterial pneumonia. Biochem. Soc. Trans. 2009, 37, 811–813. [Google Scholar] [CrossRef]
- Wiley, J.A.; Richert, L.E.; Swain, S.D.; Harmsen, A.; Barnard, D.L.; Randall, T.D.; Jutila, M.; Douglas, T.; Broomell, C.; Young, M. Inducible Bronchus-associated lymphoid tissue elicited by a protein cage nanoparticle enhances protection in mice against diverse respiratory viruses. PLoS One 2009, 4, e7142. [Google Scholar]
- Easton, A.J.; Scott, P.D.; Edworthy, N.L.; Meng, B.; Marriott, A.C.; Dimmock, N.J. A novel broad-spectrum treatment for respiratory virus infections: Influenza-based defective interfering virus provides protection against pneumovirus infection in vivo. Vaccine 2011, 29, 2777–2784. [Google Scholar] [CrossRef]
- Garcia-Crespo, K.E.; Chan, C.C.; Gabryszewski, S.J.; Percopo, C.M.; Rigaux, P.; Dyer, K.D.; Domachowske, J.B.; Rosenberg, H.F. Lactobacillus priming of the respiratory tract: Heterologous immunity and protection against lethal pneumovirus infection. Antivir. Res. 2013. submitted for publication. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dyer, K.D.; Garcia-Crespo, K.E.; Glineur, S.; Domachowske, J.B.; Rosenberg, H.F. The Pneumonia Virus of Mice (PVM) Model of Acute Respiratory Infection. Viruses 2012, 4, 3494-3510. https://doi.org/10.3390/v4123494
Dyer KD, Garcia-Crespo KE, Glineur S, Domachowske JB, Rosenberg HF. The Pneumonia Virus of Mice (PVM) Model of Acute Respiratory Infection. Viruses. 2012; 4(12):3494-3510. https://doi.org/10.3390/v4123494
Chicago/Turabian StyleDyer, Kimberly D., Katia E. Garcia-Crespo, Stephanie Glineur, Joseph B. Domachowske, and Helene F. Rosenberg. 2012. "The Pneumonia Virus of Mice (PVM) Model of Acute Respiratory Infection" Viruses 4, no. 12: 3494-3510. https://doi.org/10.3390/v4123494
APA StyleDyer, K. D., Garcia-Crespo, K. E., Glineur, S., Domachowske, J. B., & Rosenberg, H. F. (2012). The Pneumonia Virus of Mice (PVM) Model of Acute Respiratory Infection. Viruses, 4(12), 3494-3510. https://doi.org/10.3390/v4123494