1. Introduction
Filoviruses are highly lethal zoonotic agents of viral hemorrhagic fevers that are of concern as emerging pathogens and potential biothreat agents. There are no available vaccines or therapeutics approved by the United States Food and Drug Administration (FDA) for the treatment of filovirus infections, so there is a great need for continued drug discovery efforts to protect against these lethal viruses. Recently, a committee comprised of multiple research institutes and government funding agencies, the Filovirus Animal Nonclinical Group (FANG), was assembled to enhance communication, coordinate efforts, and align product development, regulatory, and scientific resources regarding all aspects of filovirus medical countermeasures (MCM) development and FDA approval thereof. To streamline product development activities funded by government agencies, the FANG has suggested concentration of MCM development efforts against two ebolaviruses, Ebola virus (EBOV) variant Kikwit (isolate 9510621, referring to the sample’s Centers for Disease Control and Prevention (CDC) registration number), Sudan virus (SUDV) variant Gulu (isolate/CDC #200011676), and one Marburg virus (MARV) variant, Angola (isolate/CDC #200501379) [
1]. Pivotal efficacy studies supporting development of a filovirus therapeutic under the Animal Rule must be conducted as Good Laboratory Practice-compliant studies (GLP, 21 CFR Part 58) to prove safety and efficacy of the compound, and ensure the integrity of the data collected. Here we have explored the viral plaque assay as the assay preferred by the FANG for filovirus titration, and which could play a role in approval of new anti‑filovirus vaccines and therapeutics under the FDA’s Animal Rule of Efficacy (21 CFR 314.600). Viral titer in a variety of biological samples from Animal Rule-driven studies can be quantified by performance of a FANG-standardized and validated viral plaque assay and included in the total data package for evaluation by the FDA [
2]. We envision that the plaque assay can be used in these therapeutic or vaccine studies to quantify virus stocks shared among institutes and verify the challenge dose administered to the animals on these studies. Most importantly, the plaque assay can be used to measure the live viral load in the serum and tissues of treated
versus untreated animals as a measure of therapeutic efficacy [
3,
4,
5].
Accurate quantitation of virus particles in a sample is essential for any antiviral research program. Presence of virus, viral genetic material, or viral antigen can be measured in a variety of ways, such as fluorescence or electron microscopy, serology, polymerase chain reaction (PCR), or quantitative reverse transcription (RT) PCR methods. However, these methods do not assess the presence of replication-competent virus in a sample. Many types of viruses can be quantitated by TCID
50 assay or plaque assay, in which one or more permissive cells in a monolayer can be infected by single plaque forming units of virus, each of which are sometimes equivalent to a single virion. Under the right conditions, infection will spread to neighboring cells to form a visible hole or “plaque” in the cell monolayer. The cell monolayer can then be stained with a dye such as crystal violet or neutral red, all plaques in the monolayer counted either by eye or under a microscope, and the titer of infectious virus in the original sample calculated. A full discussion of animal virus plaque assay theory, methods, and parameters is reviewed by Cooper [
6].
Certain assay parameters will affect the ability of a virus plaque to form. The indicator cell line must be permissive for viral replication and plaque formation, and must achieve a confluent monolayer, which provides the platform for the assay. Historically, Vero cells, which are African green monkey (
Chlorocebus sabaeus) kidney cells, have been widely used as an indicator cell line for hemorrhagic fever virus plaque formation since their discovery in 1962 [
7]. Three lineages of Vero cells have been subcultured: Vero, Vero 76 and Vero E6 (C1008), each with different properties which make them useful for bunyavirus, arenavirus, flavivirus and filovirus plaque assays [
7,
8,
9,
10,
11]. EBOV plaques were reported in early studies with fetal rhesus lung, SW-13 and Vero cells [
8,
9], while more recently published methods detail the use of Vero E6 for EBOV and MARV plaque assays [
12,
13,
14]. In addition to cell line choice, the volume of virus inocula, optimal overlay matrix and concentration, and the staining product and its optimal concentration will affect the appearance and quality of plaques and whether they can be accurately counted [
6,
8,
10,
15]. The plaque assay has enabled progress in many important areas of filovirus research, such as epidemiology, diagnostics and vaccine and therapeutics discovery and development [
3,
4,
14,
16]. To date, an optimized and standardized plaque assay protocol does not exist among filovirus researchers at various institutions, a tool which would enable comparison of results among laboratories performing FANG-driven preclinical studies to develop filovirus MCMs.
The FANG has considered that optimization and validation of a single assay protocol are the first steps towards executing GLP-compliant studies for the FDA. Therefore, the work presented here was performed to evaluate the effect of various critical parameters on the filovirus plaque assay in order to optimize the assay. We designed experiments to investigate these parameters, including cell line, inoculum volume, overlay concentration, and their effect on visualization of the plaques. Our goal was to identify the optimal reagent or process for each of these parameters, which could ultimately enable validation of a standardized plaque assay protocol for use on FANG-driven GLP studies, as well as any other filovirus research initiative choosing to make use of this assay.
3. Experimental Section
Virus. A USAMRIID cell culture stock of Ebola virus (EBOV, isolate CDC#9510621) originated from an outbreak in Kikwit, Zaire in 1995, and was passaged twice at the Centers for Disease Control and Prevention (Atlanta, GA, USA), and once in Vero E6 cells at USAMRIID for a total of 3 passages. This stock is known to have a titer of 1.5 × 106 PFU/mL using general plaque assay protocol methods at USAMRIID. A USAMRIID cell culture stock of an Angolan isolate of Marburg virus (MARV, isolate CDC#200501379), originated from an outbreak in Angola in 2005, and was passaged 4 times in Vero E6 cells. This stock is known to have a titer of 7.0 × 107 PFU/mL using general plaque assay protocol methods at USAMRIID.
Cells. Vero, Vero 76 or Vero E6 cells were purchased from American Type Culture Collection (ATCC, Manassas, VA). Catalog, lot and passage numbers at time of receipt at USAMRIID are as follows: Vero (Cat. No. CCL-81, lot 58484194, passage 123); Vero 76 (Cat. No. CRL-1587, lot 58078685, passage 33); Vero E6 (Cat. No. CRL-1586, 58027482, passage 23). Vero E6 cells were obtained from BEI Resources (Manassas, VA, USA). BEI Vero E6 cells are Cat NR-596, lot 3956812, passage 19. Passage numbers presented in the results section denote cumulative passage numbers upon receipt plus subsequent passages at USAMRIID. For example, ATCC Vero E6 cells were received at passage 23 and cultured and used up to passage 54 in the described work (
Figure 1,
Figure 2,
Figure 3). All cells were cultured under well-documented conditions, where all medium, supplements and procedures for passaging and culturing cells were performed and recorded by qualified technicians. Cells were counted by use of either a Cedex HiRes Multi Sampler System (Roche, Indianapolis, IN, USA) or a hemocytometer. Cells were plated in 6-well tissue culture dishes (CoStar, Corning Inc. Corning, NY, USA) at appropriate concentrations and days prior to use, to achieve the desired percentage confluency, as determined by the experimental design. For most experiments, cells were used at 85%–100% confluency and seeded at 500,000 to 700,000 cells per well. This is a range of densities which will provide confluent cells for use at 24 hours (closer to 700,000 cells) to 48 hours post-seeding (closer to 500,000 cells).
Cell culture media, supplements and stains. Reagents for culturing of cells: 1× MEM (Cat. No. 10010249, Cellgro, Manassas, VA, USA) in plates, supplemented with 5 to 10% FBS (vol/vol) (Cat. No. 16000-044, Gibco, Life Technologies, Grand Island, NY, USA), 5 mM L-glutamine (Cat. No. SH330034.01, Thermo Scientific HyClone, Logan, UT, USA). Reagents for plaque assay: 2× EBME (Cat. No. 05-5068EF, Gibco, Life Technologies, Grand Island, NY, USA), FBS (Cat. No. 16000-044, Gibco, Life Technologies, Grand Island, NY, USA), penicillin-streptomycin (Cat. No. 30-022-CI, Cellgro, Manassas, VA, USA), SeaKem ME Agarose (Cat. No. 50014, Lonza, Rockland, ME, USA), Neutral red from Gibco (Cat. No. 02-0066DG, Gibco, Life Technologies, Grand Island, NY, USA), Sigma (Cat. No. N2889, Sigma-Aldrich, St. Louis, MO, USA) and Ricca (Cat. No. 5271-4, Arlington, TX, USA).
Filovirus Plaque Assay. Cells were plated in 6-well plates 24 to 72 hours prior to use in assay, and plates were carried into the BSL-4 lab. SeaKem ME Agarose (Lonza, Rockland, ME, USA) was prepared for overlays by mixing an appropriate amount of agarose with an appropriate amount of distilled water to achieve the desired concentration (i.e., 1 g of agarose mixed with 100 mL of distilled water to achieve 1% agarose concentration). Agarose was autoclaved, allowed to solidify, and then melted by microwave at BSL-4 prior to use. Samples were diluted in 1× MEM, supplemented with 5% FBS (vol/vol), in a 10-fold fashion according to the experimental design. The culture medium in the 6-well plate was then removed by either swift decanting or aspiration by pipette and virus inocula of various concentrations were added to duplicate or replicate wells in volumes of 100 μL to 500 μL. Inoculum fluid was distributed by gentle rocking, manually. The plates were incubated at 37 °C for about 1 hour, with or without plate rocking every 15 to 20 minutes according to the experimental design. Without removal of the inoculum or washing the virus off of the monolayer in any way, a primary liquid overlay containing a 1:1 ratio of 2× EBME medium (supplemented with 10% FBS, 400 IU penicillin, 400 µg/mL streptomycin) equilibrated to 37–42 °C and 1% melted agarose equilibrated to 60°C degrees was overlaid in 2 ml aliquots onto the inoculated monolayer and allowed to solidify. Plates were incubated for approximately 7 days and stained with 2 mL of secondary overlay which consisted of 1:1 volume ratio of 2× EBME plus supplements: 1% agarose and the combined volumes were supplemented with 4% neutral red. Plaques were counted and recorded 24–48 hours post-staining. All work with filoviruses was carried out under maximum containment in the biosafety level 4 laboratories at USAMRIID.
Plaque counting and titer calculation. Plates were inverted on a light box and plaques were counted by hand with the aid of a marker, usually by one operator. For some experiments, plates were photographed to demonstrate plaque appearance and quality under various test conditions. Based on well size and plaque diameter, wells with less than 150 plaques and more than 10 plaques were used in the calculation of the titer [
6]. If there were wells at two different serial dilutions with counts within this range, then the counts from wells with the more concentrated dilution were chosen for calculating titer. Counts from replicate wells were averaged, and that number was multiplied by the dilution factor of the inoculum which produced that count, and the volume of inoculum plated to calculate the plaque forming units (PFU) per ml of the original stock virus preparation. The calculation is: average value of plaques in replicate wells × mL of virus inoculum × dilution factor = titer in PFU/mL.
Variables under test. Variables such as choice of Vero cell line, percentage confluency of cells in well monolayer, amount of time (in days) cells can be cultured in 6-well plates before use in assay, method of cell culture medium removal for initiation of assay, amount of virus inoculum volume, sample dilution procedures, requirement for interim plate rocking during incubation, percentage agarose in primary overlay, source of neutral red stain, concentration of stain(s) in secondary overlay, and optimal days upon which to stain and count plaques were all assessed as part of assay optimization. As experiments progressed over time, the “stability” of the cells, or effect of cell age (passage number) on plaque number and quality was evaluated in a specific experiment, and compared across all experiments, as appropriate. Specific details of the experimental design evaluating each of these variables are presented with the results for each variable.
Statistical tests. Raw data values were log-transformed for data analysis, and back transformed to present average viral titer values as mean ± standard deviation. Comparisons of sample groups were done assuming equal variance using two-tailed Student’s t-test in Microsoft Excel or GraphPad Prism software [
17], with Bonferroni correction for multiple comparisons. Coefficients of variation (% CV) were calculated and are presented in the results where possible [
18].
4. Conclusions
Generating consistent viral titer data in a plaque assay is contingent upon a number of parameters. These include monolayer cell type, cell culturing practices, cell confluency, virus sample inoculum volumes, final agarose concentrations in the primary and secondary overlays, staining product and concentration, and the optimal day for staining and counting plaques. Some of these parameters may be more critical than others. For instance, in early studies, ebolavirus plaques appeared under neutral red stain after 5–7 days in cultures which were incubated at 37 °C, but were not supplied with humidity or CO
2 at the time, due to limitations in practices at BSL-4 [
8]. The method of atmospheric control at that time was to wrap the plates in aluminum foil, and to leave them undisturbed in the incubator. Today, BSL-4 laboratories are fully equipped with humidity and CO
2-controlled incubators, but ebolavirus plaques still tend to develop in 7 days, indicating culture conditions could be somewhat flexible. On the other hand, monolayer cell type is of critical importance. Vero, fetal rhesus lung, and human adenocarcinoma cells (SW-13) have all been reported to produce filovirus plaques with varying degrees of success, and a direct comparison of many lines indicated Vero cells were superior [
8,
9,
19]. All of these parameters of varying importance must work in concert to produce clear countable plaques of a consistent size that result in reliable plaque titers.
Similarly, parameters tested in this filovirus plaque assay optimization effort, were found to vary in their effect on virus titer outcome. For example, the results demonstrated that Vero E6 cells from either ATCC or BEI were better than Vero 76 or Vero cells from ATCC for the formation of filovirus plaques, which would suggest that it is important to assess the performance of possible cell types before choosing one type on which the assay will be based. While both BEI and its parent company, ATCC, provide Vero E6 cells which performed equivalently well, the BEI cell line has been chosen for use in FANG-standardized filovirus assays. BEI Resources has generated this stock of Vero E6 cells to be set aside as a special reagent resource dedicated to Department of Health and Human Services (DHHS) and Department of Defense (DoD) government grant and contract holders, and researchers performing work funded through these contracts will likely be required to procure and use these cells in filovirus plaque assays, especially for advanced development studies. Should groups running non-DHHS or non-DoD funded projects, but perhaps related projects, not be able to obtain these cells, our data support the use of ATCC Vero E6 cells as an acceptable substitute, if needed.
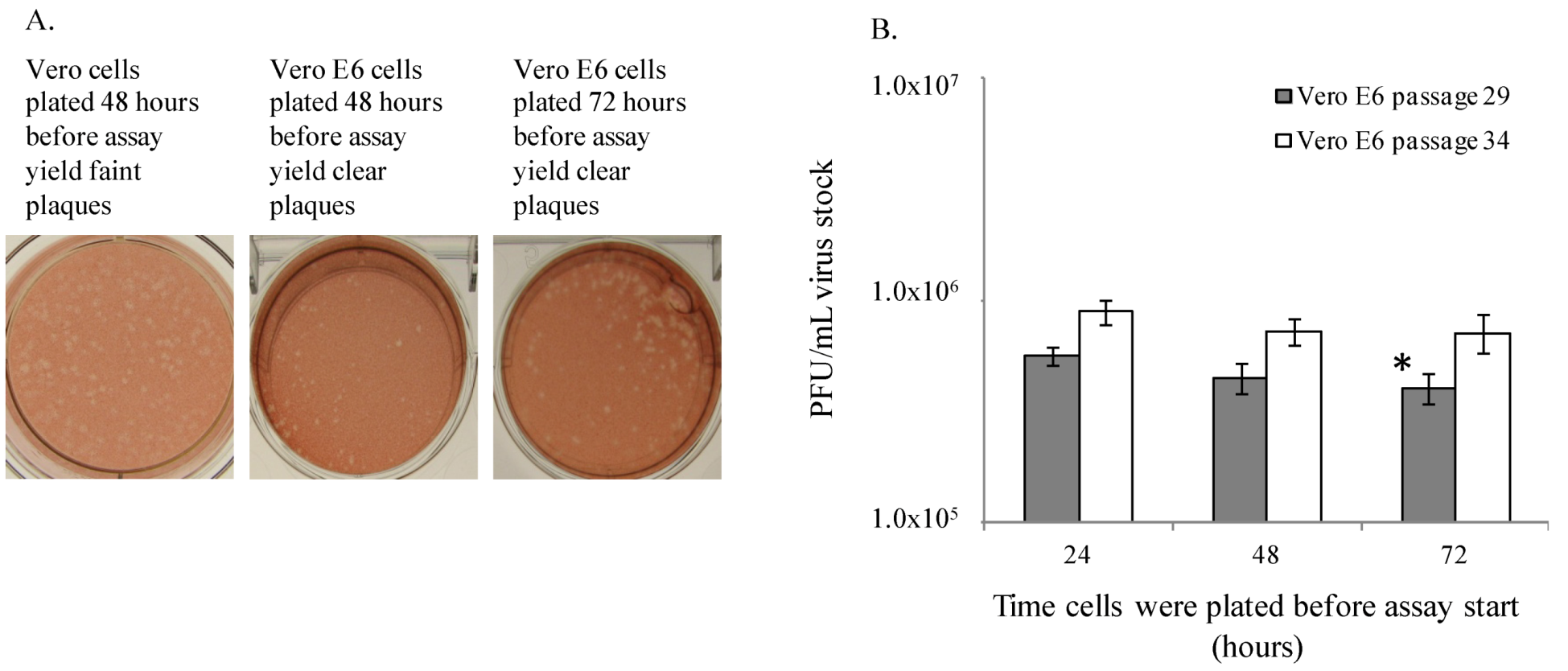
The length of time cells were seeded prior to culture (24–72 hours) before assay start did not appear to affect virus titer (
Figure 1B). There was only one statistically significant difference calculated between titers measured in cells plated at 24
vs. 72 hours before assay start (
p = 0.006), but this observation was not repeatable. Therefore, it can be concluded that the small differences in titer across the days are not biologically significant in the assay, and the one significant difference measured may have been due to sample set size and low variation in the numeric values. Monolayer confluencies above 50% were important to virus titration, but surprisingly a heavily confluent monolayer was not required for consistent plaque formation and virus titration. Additionally, an inoculum volume of approximately 300 µL provided for optimal plaque titration for both EBOV and MARV, yet MARV may be more sensitive to this phenomenon, since a 100 µL increase in inoculum volume reduced titers by almost 40% (
p < 0.008). Volumes lower than 300 µL resulted in monolayer damage, while higher volumes yielded decreased titers. Final agarose concentration, brand of neutral red product, and amount of stain did not affect plaque titer, but these variables can affect the quality of the monolayer in which plaques develop. If a monolayer were improperly overlaid or stained, the cells could be compromised and the plaque count confounded. Therefore, optimal overlay content and procedure are important parameters for collection of reproducible results.
Other staining methods or overlay matrices were not evaluated in this filovirus plaque assay, although the semi-fluid Avicel® (FMC Biopolymer, Philadelphia, PA, USA) or methyl celluose overlays work well for influenza and hemorrhagic fever viruses [
20,
21] and are sometimes used at the institutes participating in these studies (USAMRIID, unpublished data [
22]). Monolayers can also be fixed with 10% neutral buffered formalin and stained with crystal violet in ethanol, a practice which inactivates virus so completely that plaque assay plates can even be removed from containment laboratories, if desired. Photographs of the 6-well plates, rather than retaining the plates themselves, generally serve as sufficient records of plaque appearance, if required for future consideration.
Together with Department of Defense and the National Institutes of Allergy and Infectious Diseases funding agencies, USAMRIID and other BSL-4 laboratories in the United States, Canada and the United Kingdom that participate in the FANG have spearheaded research to develop filovirus medical countermeasures (MCMs) and the non-clinical assays to support this research. This research has resulted in the optimization and standardization of the FANG filovirus plaque assay protocol as a tool for use across multiple institutions, which will aid in the direct comparison of data generated at multiple worksites in support of FANG-directed studies. Government agencies intend to fund large intra-institutional advanced development contracts for filovirus therapeutics and vaccine development. Reproducibility of virus titer data internally and across institutions is paramount to the success of these programs. Additionally, the assay must be validated at each performing institution for use in GLP studies supporting therapeutic development under the FDA Animal Rule. A further intent of this study is to provide the filovirus research community with an assay protocol that could serve as a useful resource for performance of such a standardized assay. The studies presented here identified particular parameters that should be followed for performance of the standardized assay protocol, and helped to narrow the ranges of acceptable test variables when seeking to design SOPs and validation protocols for regulated studies (See
Table 3). Furthermore, the results indicated that the assay performs well for both Ebola virus and Marburg virus.
Previous experience at USAMRIID with Sudan virus (SUDV), which can behave slightly differently in culture from EBOV, leads the authors to hypothesize that this basic assay may function well for evaluation of plaques from other ebolaviruses or other filoviruses, but there may be some differences in the appropriate day to stain and count, or possibly other parameters. Continued collection of assay data and evaluation through formal validation protocols will be the best method to assess assay performance for all of the filoviruses of interest in advanced development studies.
Table 3.
Standardized filovirus plaque assay suggested for use in FANG studies.
Table 3.
Standardized filovirus plaque assay suggested for use in FANG studies.
| Assay Parameter | Recommendation for Standardized Assay based on Optimization Studies |
|---|
| Cell Type | Vero E6 cells from BEI Resources |
| Cell Passage Number | 25 to at least 40, and higher passages may be acceptable upon further testing or institutional validation of the assay |
| Cell Plating Density | 85%–100% confluent at time of usage, plates set up 24 to 48 hours prior using 500 to 700,000 cells per well |
| Inoculum volume | 300 µL total inoculum volume |
| Final agarose concentration | 0.5% SeaKem agarose |
| Neutral red | 4% Gibco Neutral Red; 5% Sigma Neutral Red |
| Staining Day | 7 days post-infection |
| Plaque Counting Day(s) | Gibco-24 and 48 hours post-staining |
| Sigma-24 hours post-staining |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
