Isolation and Identification of Feline Herpesvirus Type 1 from a South China Tiger in China

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. Preliminary Identification of FHV-1 by Molecular Biological Methods
2.1.1.1. PCR/RT-PCR Assays of Clinical Samples for Three Suspicious Pathogens
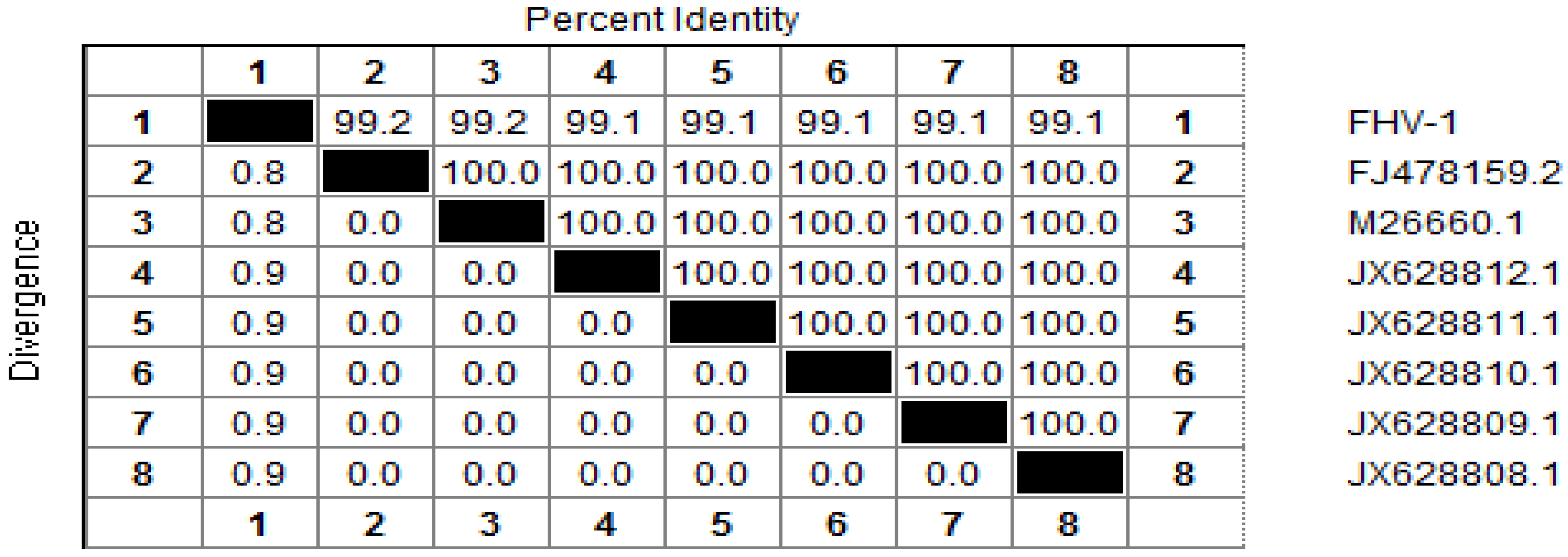
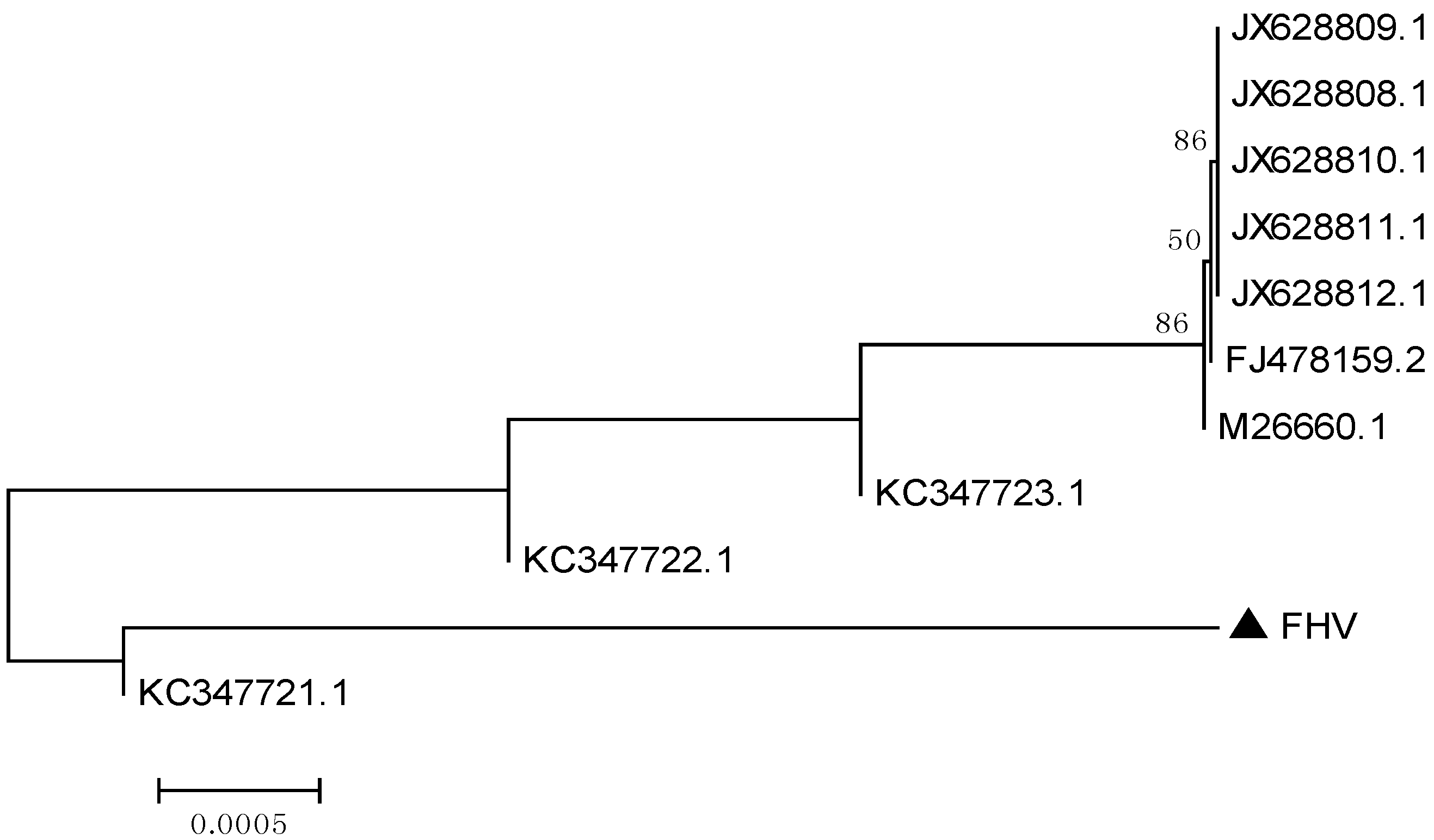
2.1.1.2. Phylogenetic Analysis Based on Two Cloned Gene Fragments of FHV-1


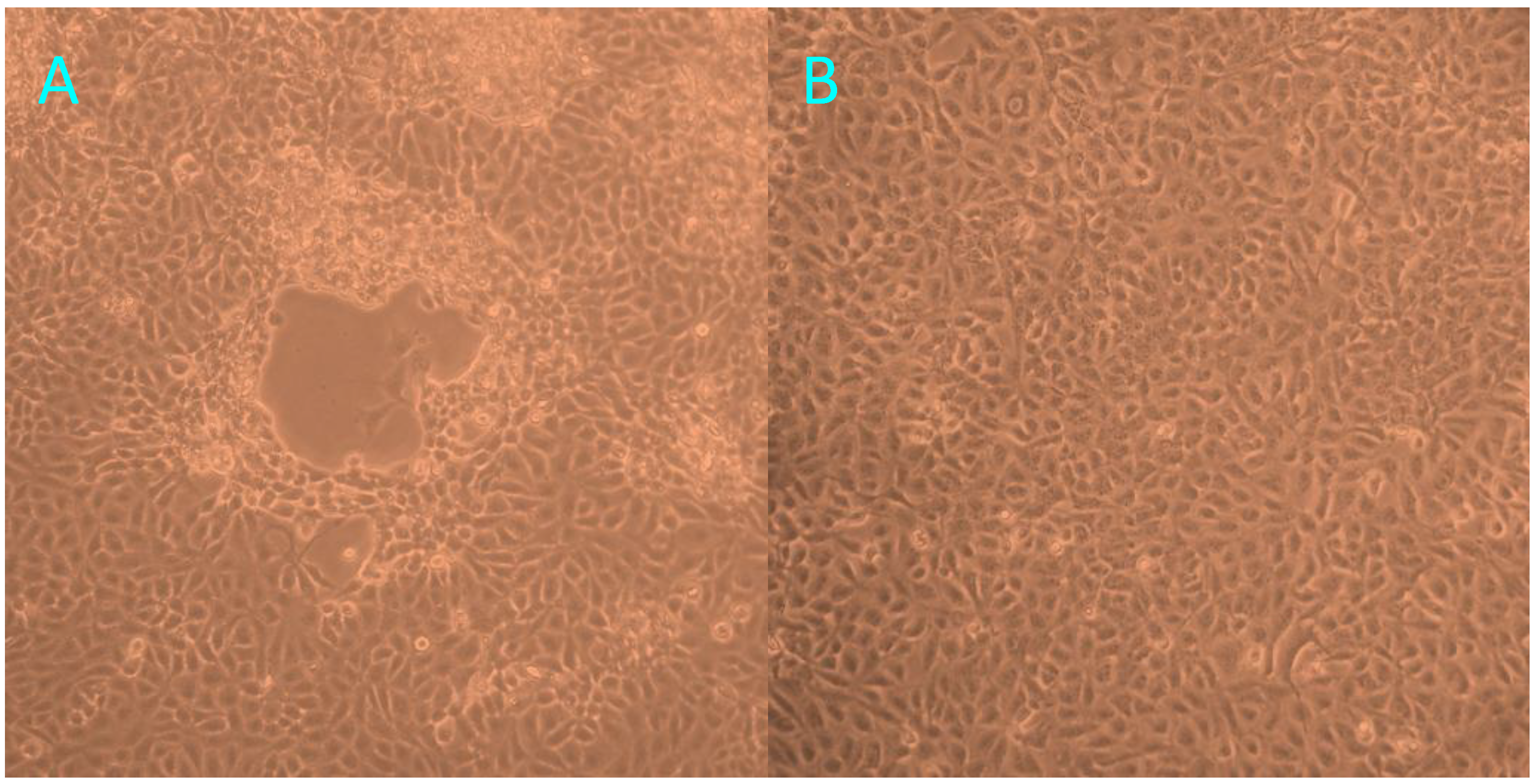
2.1.2. Virus Isolation and TEM Examination of the Cell Cultures


2.1.3. Cat Challenge Test with the Virus Isolate
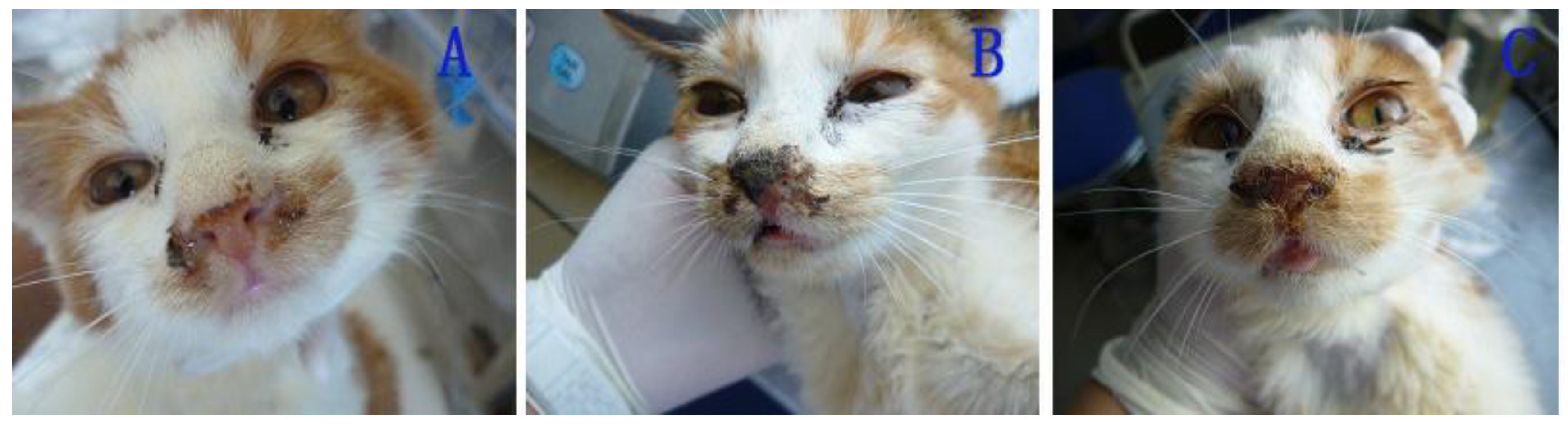
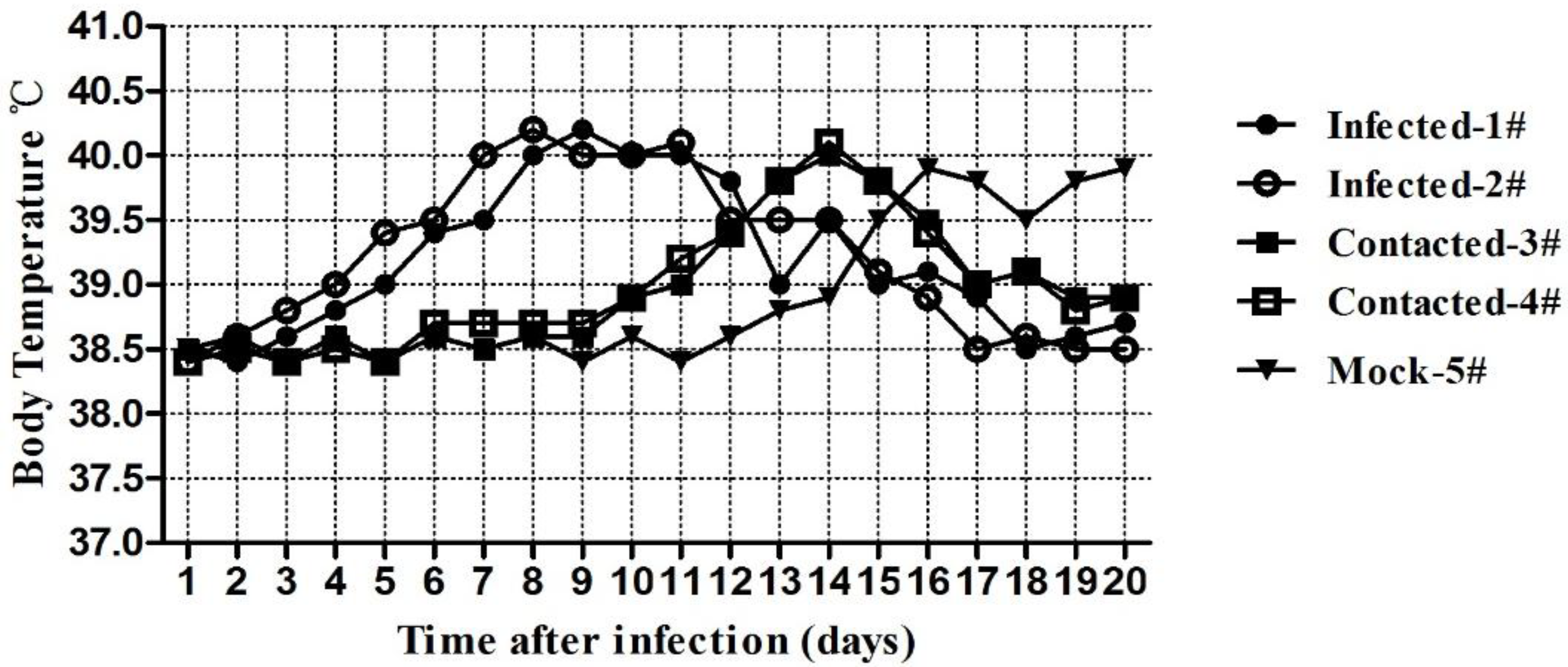
2.1.3.1. Clinical Symptoms of the Challenged Cats


2.1.3.2. Virus Shedding in the Infected Cats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cat No. | Days post-inoculation | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | |
| 1# | − | − | + | + | + | + | + | + | + | + |
| 2# | − | − | + | + | + | + | + | + | + | + |
| 3# | − | − | − | − | + | + | + | + | + | + |
| 4# | − | − | − | − | + | + | + | + | + | + |
| 5# | − | − | − | − | − | − | − | + | + | + |
2.2. Discussion
3. Experimental Section
3.1. Case Description and Sample Collection
3.2. Specimen Treatment
3.3. Molecular Identification
3.3.1. PCR/RT-PCR Assays
| Virus | Target gene | Primer sequence 5'-3' | Amplified fragment | Reaction systems (50 μL) | Conditions for PCR | Reference |
|---|---|---|---|---|---|---|
| FHV-1 | TK-F TK-R | GACGTGGTGAATTATCAGC CAACTAGATTTCCACCAGGA | 292 bp | 10× ExTaq Buffer, 5 μLdNTP (2.5 mmol/L), 4 μL F/R primer, 1 μL Template, 2 μL ExTaq polymerase, 1 μL ddH2O, 36 μL | Fore- denaturalization: 94 °C, 5 min. Denaturalization: 94 °C, 30 s. Anneal: 56 °C, 30 s. Extension: 72 °C, 30 s. 30 cycles, extension: 72 °C, 10 min. | [7] |
| FCV | Cali1 Cali2 | AACCTGCGCTAACGTGCTTA CAGTGACAATACACCCAGAAG | 924 bp | 10× ExTaq Buffer, 5 μL dNTP (2.5 mmol/L), 4 μL A/B primer, 1 μL Template, 2 μL ExTaq polymerase, 1 μL ddH2O, 36 μL | Fore- denaturalization: 94 °C, 2 min. Denaturalization: 94 °C, 60 s. Anneal: 56 °C, 45 s. Extension: 72 °C, 60 s. 35 cycles, extension: 72 °C, 7 min. | [22] |
| CDV | NfpNrp | GCTGGTTGGAGAATAAGG CCAACTCCCATAGCATAA | 586 bp | 10× ExTaq Buffer, 5 μL dNTP (2.5 mmol/L), 4 μL A/B primer, 1 μL Template, 2 μL ExTaq polymerase, 1 μL ddH2O, 36 μL | Fore- denaturalization: 94 °C, 3 min. Denaturalization: 94 °C, 30 s. Anneal: 60 °C, 30 s. Extension: 72 °C, 30 s. 33 cycles, extension: 72 °C, 10 min. | [23] |
3.3.2. Gene Cloning and Sequence Analysis
3.4. Virus Culture
3.5. Electron Microscopy Examination
3.6. Animal Challenge Test
4. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References and Notes
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef]
- Maes, R.K.; Fritsch, S.L.; Herr, L.L.; Rota, P.A. Immunogenic proteins of feline rhinotracheitis virus. J. Virol. 1984, 51, 259–262. [Google Scholar]
- Povey, R.C. A review of feline viral rhinotracheitis (feline herpesvirus I infection). Comp. Immunol. Microbiol. Infect. Dis. 1979, 2, 373–387. [Google Scholar] [CrossRef]
- Dowers, K.L.; Hawley, J.R.; Brewer, M.M.; Morris, A.K.; Radecki, S.V.; Lappin, M.R. Association of Bartonella species, feline calicivirus, and feline herpesvirus 1 infection with gingivostomatitis in cats. J. Feline Med. Surg. 2010, 12, 314–321. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Wang, B.; Liu, J.; Tian, K. Isolation and identification of feline herpesvirus type 1. Lab. Anim. Sci. 2010, 27, 21–25. [Google Scholar]
- Tilson, R.; Defu, H.; Muntifering, J.; Nyhus, P.J. Dramatic decline of wild South China tigers Panthera tigris amoyensis: Field survey of priority tiger reserves. Oryx 2004, 38, 40–47. [Google Scholar]
- Sykes, J.E.; Allen, J.L.; Studdert, V.P.; Browning, G.F. Detection of feline calicivirus, feline herpesvirus 1 and Chlamydia psittaci mucosal swabs by multiplex RT-PCR/PCR. Vet. Microbiol. 2001, 81, 95–108. [Google Scholar] [CrossRef]
- Willoughby, K.; Bennett, M.; McCracken, C.M.; Gaskell, R.M. Molecular phylogenetic analysis of felid herpesvirus 1. Vet. Microbiol. 1999, 69, 93–97. [Google Scholar] [CrossRef]
- Herrmann, S.C.; Gaskell, R.M.; Ehlers, B.; Ludwig, H. Characterization of the feline herpesvirus genome and molecular epidemiology of isolates from natural outbreaks and latent infections. In Latent Herpesvirus Infections in Veterinary Medicine; Wittmann, G., Gaskell, R.M., Rziha, H.-J., Eds.; Springer: Heidelberg, Germany, 1984; pp. 321–336. [Google Scholar]
- MEGA-5, Molecular Evolutionary Genetics Analysis Version 5.2. Available online: http://www.megasoftware.net/ (accessed on 24 February 2014).
- Gaskell, R.M.; Povey, R.C. The dose response of cats to experimental infection with feline viral rhinotracheitis virus. J. Comp. Pathol. 1979, 89, 179–191. [Google Scholar] [CrossRef]
- Hoover, E.A.; Rohovsky, M.W.; Griesemer, R.A. Experimental feline viral rhinotracheitis in the germfree cat. Am. J. Pathol. 1970, 58, 269–282. [Google Scholar]
- Gaskell, R.M.; Povey, R.C. Experimental induction of feline viral rhinotracheitis (FVR) virus re-excretion in FVR-recovered cats. Vet. Rec. 1977, 100, 128–133. [Google Scholar]
- Shields, R.P.; Gaskin, J.M. Fatal generalized feline viral rhinotracheitis in a young adult cat. J. Am. Vet. Med. Assoc. 1977, 170, 439–441. [Google Scholar]
- Spradbrow, P.B.; Carlisle, C.; Watt, D.A. The association of a herpesvirus with generalized disease in a kitten. Vet. Rec. 1971, 89, 542–544. [Google Scholar]
- Van Pelt, C.S.; Crandell, R.A. Pancreatitis associated with a feline herpesvirus infection. Comp. Anim. Pract. 1987, 1, 7. [Google Scholar]
- Gaskell, R.M.; Radford, A.D.; Dawson, S. Feline infectious respiratory disease. In Feline Medicine and Therapeutics; Chandler, E.A., Gaskell, C.J., Gaskell, R.M., Eds.; Blackwell Publishing: Oxford, UK, 2004; pp. 577–595. [Google Scholar]
- Eleraky, N.Z.; Potgieter, L.N.; Kennedy, M.A. Virucidal efficacy of four new disinfectants. J. Am. Anim. Hosp. Assoc. 2002, 38, 231–234. [Google Scholar]
- Evermann, J.F.; Laurenson, M.; McKeirnan, A.J.; Caro, T.M. Infectious disease surveillance in captive and free-living cheetahs—An integral part of the species survival plan. Zoo Biol. 1993, 12, 125–133. [Google Scholar] [CrossRef]
- Hofmann-Lehmann, R.; Fehr, D.; Grob, M.; Elgizoli, M.; Packer, C.; Martenson, J.S.; O’Brien, S.J.; Lutz, H. Prevalence of antibodies to feline parvovirus, calicivirus, herpesvirus, coronavirus, and immunodeficiency virus and of feline leukemia virus antigen and the interrelationship of these viral infections in free-ranging lions in east Africa. Clin. Diagn. Lab. Immunol. 1996, 3, 554–562. [Google Scholar]
- Daniels, M.J.; Golder, M.C.; Jarrett, O.; MacDonald, D.W. Feline viruses in wild cats from Scotland. J. Wildl. Dis. 1999, 35, 121–124. [Google Scholar] [CrossRef]
- Marsilio, F.; di Martino, B.; Decaro, N.; Buonavoglia, C. A novel nested PCR for the diagnosis of calicivirus infections in the cat. Vet. Microbiol. 2005, 105, 1–7. [Google Scholar] [CrossRef]
- Ju, H.; Xia, X.; Gao, Y.; Yang, S.; Feng, N.; Wang, T.; Li, Y. Detection of canine distemper virus from samples of dead animals by RT-PCR. J. Jilin Agric. Univ. 2006, 28, 317–320. [Google Scholar]
- Miranda-Vizuete, A.; Damdimopoulos, A.E.; Pedrajas, J.R.; Gustafsson, J.Å.; Spyrou, G. Human mitochondrial thioredoxin reductase. Euro. J. Biochem. 1999, 261, 405–412. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sun, H.; Li, Y.; Jiao, W.; Liu, C.; Liu, X.; Wang, H.; Hua, F.; Dong, J.; Fan, S.; Yu, Z.; et al. Isolation and Identification of Feline Herpesvirus Type 1 from a South China Tiger in China. Viruses 2014, 6, 1004-1014. https://doi.org/10.3390/v6031004
Sun H, Li Y, Jiao W, Liu C, Liu X, Wang H, Hua F, Dong J, Fan S, Yu Z, et al. Isolation and Identification of Feline Herpesvirus Type 1 from a South China Tiger in China. Viruses. 2014; 6(3):1004-1014. https://doi.org/10.3390/v6031004
Chicago/Turabian StyleSun, Heting, Yuanguo Li, Weiyi Jiao, Cunfa Liu, Xiujuan Liu, Haijun Wang, Fuyou Hua, Jianxiu Dong, Shengtao Fan, Zhijun Yu, and et al. 2014. "Isolation and Identification of Feline Herpesvirus Type 1 from a South China Tiger in China" Viruses 6, no. 3: 1004-1014. https://doi.org/10.3390/v6031004
APA StyleSun, H., Li, Y., Jiao, W., Liu, C., Liu, X., Wang, H., Hua, F., Dong, J., Fan, S., Yu, Z., Gao, Y., & Xia, X. (2014). Isolation and Identification of Feline Herpesvirus Type 1 from a South China Tiger in China. Viruses, 6(3), 1004-1014. https://doi.org/10.3390/v6031004