
Identifying Recent HIV Infections: From Serological Assays to Genomics

 and
and
Abstract
:1. Introduction
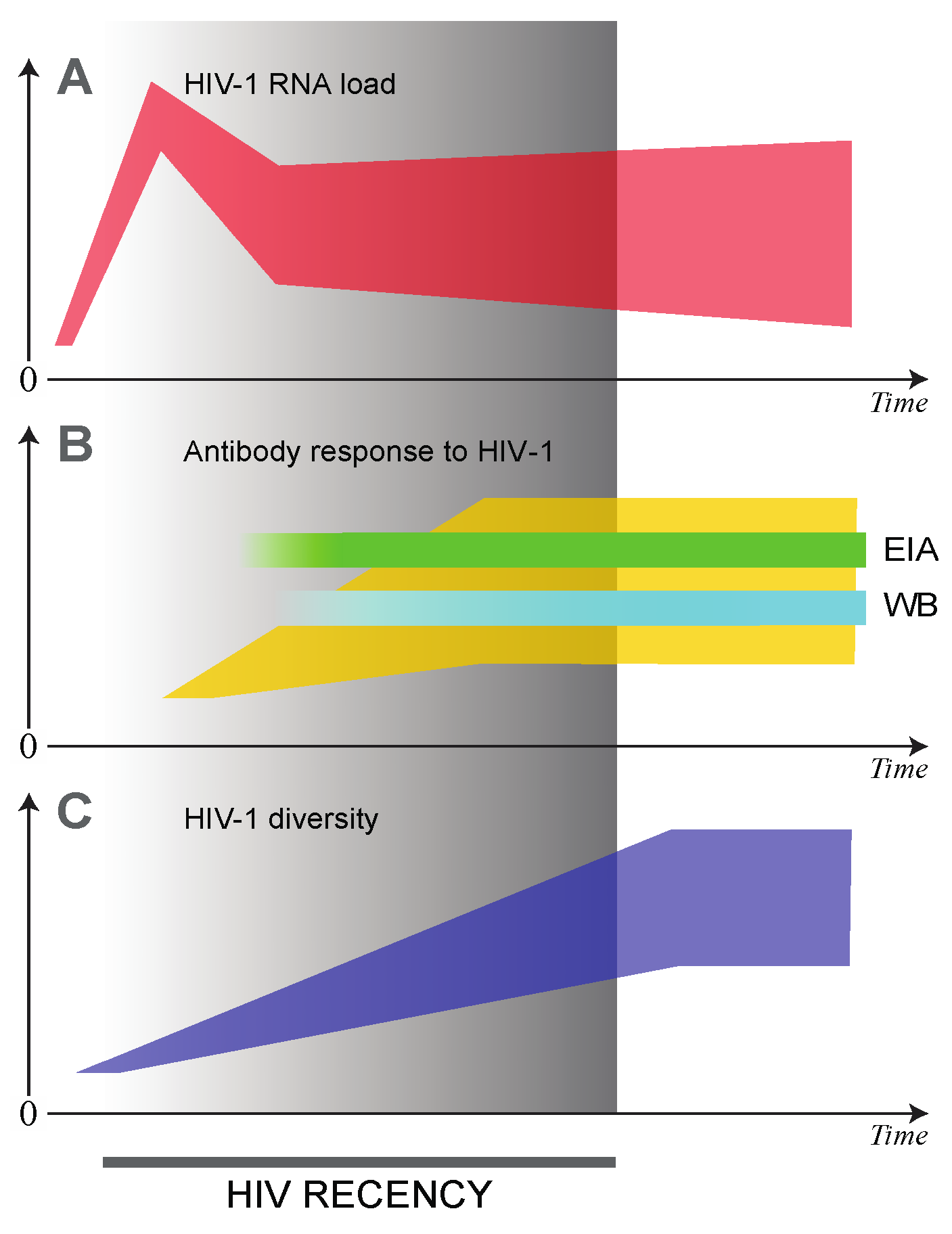
2. Early Dynamics of HIV Infection

3. Serological Tests of Recent HIV Infection
3.1. Less Sensitive Enzyme Immunoassays (LS-EIA)
3.2. Proportion of HIV-1 Specific IgG Antibodies
3.3. Antibody Avidity Assays
3.4. Anti-p24 IgG3
3.5. Inno-LIA HIV Adaptation
3.6. Challenges Associated with the Application of Serological Assays in Cross-Sectional Data

{kind=link}
| Assay or Test Type or Recent Infection Algorithm | Brief Summary of the Assay or Test | Strengths | Limitations | References |
|---|---|---|---|---|
| High Resolution Melting Assay | Measures diversity of generated amplicons by using melting temperature of DNA duplexes. |
|
| [69,70,71] |
| Counting Sequence Ambiguities | Ambiguous bases in the viral sequence indicate heterogeneous virus population. The number of ambiguous bases is small in recently infected individuals and increased overtime. |
|
| [18,19,72,73] |
| Naïve Bayes Classifier (NBC) | Utilized the frequency of ambiguous sites together with CD4+ cell counts and any concurrent AIDS defining illness. The Bayesian probability framework estimates the probability of a patient to be in one of four stages of HIV infection. |
|
| [74] |
| Hamming Distance (HD) | The HD is a number that denotes the difference between two sequences of equal length. It is the simplest measure of HIV diversity. HD can measure the number of nucleotide differences between a pair of virus sequences. If applied to viral quasispecies from the host, HD can estimate the stage of HIV infection. |
|
| [21,75] |
| Sequence Clustering Based Diversity Measure (SCBD) | Intra-cluster genetic diversity is used as the measure of time since infection. Inter-cluster diversity is used to determine whether there were multiple founder strains and the dot matrix incorporates information on indels and recombination. |
|
| [76] |
| Multi-Assay Algorithms (MMA) | Results of serology-based test of recent infection combined with: Clinical data (e.g., CD4+ cell counts, HIV-1 RNA load, ART status) Measure of HIV diversity Combination of Assays can be optimized to increase accuracy. |
|
| [69,77,78] |
4. Molecular Tests of Recent HIV Infection
4.1. High Resolution Melting Assay
4.2. Sequence Ambiguities as a Marker of Recent HIV Infection
4.3. Naïve Bayes Classifiers
4.4. Hamming Distances
4.5. Sequence Clustering Based Diversity Measure
5. Multi-Assay Algorithms
6. Discussion
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fiamma, A.; Lissouba, P.; Amy, O.E.; Singh, B.; Laeyendecker, O.; Quinn, T.C.; Taljaard, D.; Auvert, B. Can HIV incidence testing be used for evaluating HIV intervention programs? A reanalysis of the Orange Farm male circumcision trial (ANRS-1265). BMC Infect. Dis. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.H.; Li, X.; Kigozi, G.; Serwadda, D.; Nalugoda, F.; Watya, S.; Reynolds, S.J.; Wawer, M. The impact of male circumcision on HIV incidence and cost per infection prevented: A stochastic simulation model from Rakai, Uganda. AIDS 2007, 21, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Hall, H.I.; Green, T.A.; Wolitski, R.J.; Holtgrave, D.R.; Rhodes, P.; Lehman, J.S.; Durden, T.; Fenton, K.A.; Mermin, J.H. Estimated future HIV prevalence, incidence, and potential infections averted in the United States: A multiple scenario analysis. J. Acquir. Immune Defic. Syndr. 2010, 55, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B.G.; Roger, M.; Routy, J.-P.; Moisi, D.; Ntemgwa, M.; Matte, C.; Baril, J.-G.; Thomas, R.; Rouleau, D.; Bruneau, J. High rates of forward transmission events after acute/early HIV-1 infection. J. Infect. Dis. 2007, 195, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Lutalo, T.; Wawer, M.; Serwadda, D.; Sewankambo, N.; Nalugoda, F.; Makumbi, F.; Wabwire-Mangen, F.; Kiwanuka, N.; Kigozi, G. HIV incidence and sexually transmitted disease prevalence associated with condom use: A population study in Rakai, Uganda. AIDS 2001, 15, 2171–2179. [Google Scholar] [CrossRef] [PubMed]
- Wawer, M.J.; Gray, R.H.; Sewankambo, N.K.; Serwadda, D.; Li, X.; Laeyendecker, O.; Kiwanuka, N.; Kigozi, G.; Kiddugavu, M.; Lutalo, T.; et al. Rates of HIV-1 transmission per coital act, by stage of HIV-1 infection, in Rakai, Uganda. J. Infect. Dis. 2005, 191, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, M.-R.; Anderson, J.A.; Giorgi, E.E.; Seoighe, C.; Mlisana, K.; Ping, L.-H.; Athreya, G.S.; Treurnicht, F.K.; Keele, B.F.; Wood, N.; et al. Quantitating the multiplicity of infection with human immunodeficiency virus type 1 subtype C reveals a non-poisson distribution of transmitted variants. J. Virol. 2009, 83, 3556–3567. [Google Scholar] [CrossRef] [PubMed]
- Lagakos, S.; Gable, A. Methodological Challenges in Biomedical HIV Prevention Trials; National Academy Press: Washington, DC, USA, 2008. [Google Scholar]
- Barnighausen, T.; Tanser, F.; Gqwede, Z.; Mbizana, C.; Herbst, K.; Newell, M. High HIV incidence in a community with high HIV prevalence in rural South Africa: Findings from a prospective population-based study. AIDS 2008, 22, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Kumwenda, N.; Hoffman, I.; Chirenje, M.; Kelly, C.; Coletti, A.; Ristow, A.; Martinson, F.; Brown, J.; Chilongozi, D.; Richardson, B. HIV incidence among women of reproductive age in Malawi and Zimbabwe. Sex. Transm. Dis. 2006, 33, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Ghys, P.; Kufa, E.; George, M. Measuring trends in prevalence and incidence of HIV infection in countries with generalised epidemics. Sex. Transm. Infect. 2006, 82 (Suppl. 1), i52–i56. [Google Scholar] [CrossRef] [PubMed]
- Brookmeyer, R.; Konikoff, J.; Laeyendecker, O.; Eshleman, S.H. Estimation of HIV incidence using multiple biomarkers. Am. J. Epidemiol. 2013, 177, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Busch, M.P.; Pilcher, C.D.; Mastro, T.D.; Kaldor, J.; Vercauteren, G.; Rodriguez, W.; Rousseau, C.; Rehle, T.M.; Welte, A.; Averill, M.D. Beyond detuning: 10 Years of progress and new challenges in the development and application of assays for HIV incidence estimation. AIDS 2010, 24, 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Guy, R.; Gold, J.; Calleja, J.M.; Kim, A.A.; Parekh, B.; Busch, M.; Rehle, T.; Hargrove, J.; Remis, R.S.; Kaldor, J.M. Accuracy of serological assays for detection of recent infection with HIV and estimation of population incidence: A systematic review. Lancet Infect. Dis. 2009, 9, 747–759. [Google Scholar] [CrossRef]
- Incidence Assay Critical Path Working Group. More and better information to tackle HIV epidemics: Towards improved HIV incidence assays. PLoS Med. 2011, 8, e1001045. [Google Scholar]
- Le Vu, S.; Pillonel, J.; Semaille, C.; Bernillon, P.; le Strat, Y.; Meyer, L.; Desenclos, J. Principles and uses of HIV incidence estimation from recent infection testing—A review. Eur. Surveill. 2008, 13, 537–545. [Google Scholar]
- Murphy, G.; Parry, J.V. Assays for the detection of recent infections with human immunodeficiency virus type 1. Euro. Surveill. 2008, 13, 314–320. [Google Scholar]
- Kouyos, R.D.; von Wyl, V.; Yerly, S.; Böni, J.; Rieder, P.; Joos, B.; Taffé, P.; Shah, C.; Bürgisser, P.; Klimkait, T. Ambiguous nucleotide calls from population-based sequencing of HIV-1 are a marker for viral diversity and the age of infection. Clin. Infect. Dis. 2011, 52, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Ragonnet-Cronin, M.; Aris-Brosou, S.; Joanisse, I.; Merks, H.; Vallée, D.; Caminiti, K.; Rekart, M.; Krajden, M.; Cook, D.; Kim, J. Genetic diversity as a marker for timing infection in HIV-infected patients: Evaluation of a 6-month window and comparison with BED. J. Infect. Dis. 2012, 206, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.K.; Schito, M.; Welte, A.; Rousseau, C.; Fitzgibbon, J.; Keele, B.; Shapiro, S.; McMichael, A.; Burns, D.N. Workshop summary: Novel biomarkers for HIV incidence assay development. AIDS Res. Hum. Retrovir. 2012, 28, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Allam, O.; Samarani, S.; Ahmad, A. Hammering out HIV-1 incidence with Hamming distance. AIDS 2011, 25, 2047–2048. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.; Khaki, L.; Lindsey, J.C.; Fry, C.; Cousins, M.M.; Siliciano, R.F.; Violari, A.; Palumbo, P.; Eshleman, S.H. Association of pol diversity with antiretroviral treatment outcomes among HIV-infected African children. PLoS ONE 2013, 8, e81213. [Google Scholar] [CrossRef] [PubMed]
- Welte, A.; McWalter, T.A.; Laeyendecker, O.; Hallett, T.B. Using tests for recent infection to estimate incidence: Problems and prospects for HIV. Eur. Surveill. 2010, 15. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19589 (accessed on 1 July 2015). [Google Scholar]
- Mastro, T.D.; Kim, A.A.; Hallett, T.; Rehle, T.; Welte, A.; Laeyendecker, O.; Oluoch, T.; Garcia-Calleja, J.M. Estimating HIV incidence in populations using tests for recent infection: Issues, challenges and the way forward. J. HIV AIDS Surveill. Epidemiol. 2010, 2, 1–14. [Google Scholar] [PubMed]
- Kassanjee, R.; McWalter, T.A.; Barnighausen, T.; Welte, A. A new general biomarker-based incidence estimator. Epidemiology 2012, 23, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Kassanjee, R.; Pilcher, C.D.; Keating, S.M.; Facente, S.N.; McKinney, E.; Price, M.A.; Martin, J.N.; Little, S.; Hecht, F.M.; Kallas, E.G.; et al. Independent assessment of candidate HIV incidence assays on specimens in the CEPHIA repository. AIDS 2014, 28, 2439–2449. [Google Scholar] [CrossRef] [PubMed]
- Duong, Y.T.; Qiu, M.; De, A.K.; Jackson, K.; Dobbs, T.; Kim, A.A.; Nkengasong, J.N.; Parekh, B.S. Detection of recent HIV-1 infection using a new limiting-antigen avidity assay: Potential for HIV-1 incidence estimates and avidity maturation studies. PLoS ONE 2012, 7, e33328. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.S.; Gay, C.L.; Busch, M.P.; Hecht, F.M. The detection of acute HIV infection. J. Infect. Dis. 2010, 202, S270–S277. [Google Scholar] [CrossRef] [PubMed]
- McMichael, A.J.; Borrow, P.; Tomaras, G.D.; Goonetilleke, N.; Haynes, B.F. The immune response during acute HIV-1 infection: Clues for vaccine development. Nat. Rev. Immunol. 2010, 10, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Powers, K.A.; Miller, W.C.; Pilcher, C.D.; Mapanje, C.; Martinson, F.E.; Fiscus, S.A.; Chilongozi, D.A.; Namakhwa, D.; Price, M.A.; Galvin, S.R.; et al. Improved detection of acute HIV-1 infection in sub-Saharan Africa: Development of a risk score algorithm. AIDS 2007, 21, 2237–2242. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.E.; Pilcher, C.D.; Busch, M.P.; Cohen, M.S. How can we better identify early HIV infections? Curr. Opin. HIV AIDS 2015, 10, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Aulagnier, M.; Janssens, W.; de Beer, I.; van Rooy, G.; Gaeb, E.; Hesp, C.; van der Gaag, J.; Rinke de Wit, T.F. Incidence of HIV in Windhoek, Namibia: Demographic and socio-economic associations. PLoS ONE 2011, 6, e25860. [Google Scholar] [CrossRef] [PubMed]
- Draenert, R.; Verrill, C.L.; Tang, Y.; Allen, T.M.; Wurcel, A.G.; Boczanowski, M.; Lechner, A.; Kim, A.Y.; Suscovich, T.; Brown, N.V.; et al. Persistent recognition of autologous virus by high-avidity CD8 T cells in chronic, progressive human immunodeficiency virus type 1 infection. J. Virol. 2004, 78, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Parekh, B.S.; Kennedy, M.S.; Dobbs, T.; Pau, C.P.; Byers, R.; Green, T.; Hu, D.J.; Vanichseni, S.; Young, N.L.; Choopanya, K.; et al. Quantitative detection of increasing HIV type 1 antibodies after seroconversion: A simple assay for detecting recent HIV infection and estimating incidence. AIDS Res. Hum. Retrovir. 2002, 18, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Suligoi, B.; Massi, M.; Galli, C.; Sciandra, M.; di Sora, F.; Pezzotti, P.; Recchia, O.; Montella, F.; Sinicco, A.; Rezza, G. Identifying recent HIV infections using the avidity index and an automated enzyme immunoassay. J. Acquir. Immune Defic. Syndr. 2003, 32, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Laeyendecker, O.; Brookmeyer, R.; Cousins, M.M.; Mullis, C.E.; Konikoff, J.; Donnell, D.; Celum, C.; Buchbinder, S.P.; Seage, G.R., 3rd; Kirk, G.D.; et al. HIV incidence determination in the United States: A multiassay approach. J. Infect. Dis. 2013, 207, 232–239. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. When and How to Use Assays for Recent Infection to Estimate HIV Incidence at a Population Level; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Janssen, R.S.; Satten, G.A.; Stramer, S.L.; Rawal, B.D.; O’Brien, T.R.; Weiblen, B.J.; Hecht, F.M.; Jack, N.; Cleghorn, F.R.; Kahn, J.O.; et al. New testing strategy to detect early HIV-1 infection for use in incidence estimates and for clinical and prevention purposes. J. Am. Med. Assoc. 1998, 280, 42–48. [Google Scholar] [CrossRef]
- Kothe, D.; Byers, R.H.; Caudill, S.P.; Satten, G.A.; Janssen, R.S.; Hannon, W.H.; Mei, J.V. Performance characteristics of a new less sensitive HIV-1 enzyme immunoassay for use in estimating HIV seroincidence. J. Acquir. Immune Defic. Syndr. 2003, 33, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Soroka, S.D.; Granade, T.C.; Candal, D.; Parekh, B.S. Modification of rapid human immunodeficiency virus (HIV) antibody assay protocols for detecting recent HIV seroconversion. Clin. Diagn. Lab. Immunol. 2005, 12, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Soroka, S.D.; Granade, T.C.; Phillips, S.; Parekh, B. The use of simple, rapid tests to detect antibodies to human immunodeficiency virus types 1 and 2 in pooled serum specimens. J. Clin. Virol. 2003, 27, 90–96. [Google Scholar] [CrossRef]
- Young, C.L.; Hu, D.J.; Byers, R.; Vanichseni, S.; Young, N.L.; Nelson, R.; Mock, P.A.; Choopanya, K.; Janssen, R.; Mastro, T.D.; et al. Evaluation of a sensitive/less sensitive testing algorithm using the bioMerieux Vironostika-LS assay for detecting recent HIV-1 subtype B or E infection in Thailand. AIDS Res. Hum. Retrovir. 2003, 19, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Calypte Biomedical Corporation. AwareTM BEDTM EIA HIV-1 Incidence (IgG-Capture HIV-EIA) Enzyme Immunoassay for Population Estimates of HIV-1 Incidence; Calypte Biomedical Corporation: Portland, OR, USA, 2008. [Google Scholar]
- Marinda, E.T.; Hargrove, J.; Preiser, W.; Slabbert, H.; van Zyl, G.; Levin, J.; Moulton, L.H.; Welte, A.; Humphrey, J. Significantly diminished long-term specificity of the BED capture enzyme immunoassay among patients with HIV-1 with very low CD4 counts and those on antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2010, 53, 496–499. [Google Scholar] [CrossRef] [PubMed]
- Moyo, S.; LeCuyer, T.; Wang, R.; Gaseitsiwe, S.; Weng, J.; Musonda, R.; Bussmann, H.; Mine, M.; Engelbrecht, S.; Makhema, J.; et al. Evaluation of the false recent classification rates of multiassay algorithms in estimating HIV type 1 subtype C incidence. AIDS Res. Hum. Retrovir. 2014, 29, 29–36. [Google Scholar] [CrossRef]
- Karita, E.; Price, M.; Hunter, E.; Chomba, E.; Allen, S.; Fei, L.; Kamali, A.; Sanders, E.J.; Anzala, O.; Katende, M. Investigating the utility of the HIV-1 BED capture enzyme immunoassay using cross-sectional and longitudinal seroconverter specimens from Africa. AIDS 2007, 21, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Laeyendecker, O.; Brookmeyer, R.; Oliver, A.E.; Mullis, C.E.; Eaton, K.P.; Mueller, A.C.; Jacobson, L.P.; Margolick, J.B.; Brown, J.; Rinaldo, C.R.; et al. Factors associated with incorrect identification of recent HIV infection using the BED capture immunoassay. AIDS Res. Hum. Retrovir. 2012, 28, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Longosz, A.F.; Serwadda, D.; Nalugoda, F.; Kigozi, G.; Franco, V.; Gray, R.H.; Quinn, T.C.; Eshleman, S.H.; Laeyendecker, O. Impact of HIV subtype on performance of the limiting antigen-avidity enzyme immunoassay, the bio-rad avidity assay, and the BED capture immunoassay in Rakai, Uganda. AIDS Res. Hum. Retrovir. 2014, 30, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Hargrove, J.; van Schalkwyk, C.; Eastwood, H. Bed estimates of HIV incidence: Resolving the differences, making things simpler. PLoS ONE 2012, 7, e29736. [Google Scholar] [CrossRef] [PubMed]
- Brookmeyer, R. On the statistical accuracy of biomarker assays for HIV incidence. J. Acquir. Immune Defic. Syndr. 2010, 54, 406–414. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS. UNAIDS Reference Group on estimates, modelling and projections—Statement on the use of the BED assay for the estimation of HIV-1 incidence for surveillance or epidemic monitoring. Wkly Epidemiol. Rec. 2006, 81, 40. [Google Scholar]
- Wei, X.; Liu, X.; Dobbs, T.; Kuehl, D.; Nkengasong, J.N.; Hu, D.J.; Parekh, B.S. Development of two avidity-based assays to detect recent HIV type 1 seroconversion using a multisubtype gp41 recombinant protein. AIDS Res. Hum. Retrovir. 2010, 26, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.I.; Wilson, S.; O’Toole, C.M.; Lister, C.M.; Saeed, A.M.; Watkins, R.P.; Morgan-Capner, P. Differential maturation of avidity of IgG antibodies to gp41, p24 and p17 following infection with HIV-1. Clin. Exp. Immunol. 1996, 103, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Broliden, P.A.; Morfeldt-Mansson, L.; Rosen, J.; Jondal, M.; Wahren, B. Fine specificity of IgG subclass response to group antigens in HIV-1-infected patients. Clin. Exp. Immunol. 1989, 76, 216–221. [Google Scholar] [PubMed]
- Tomaras, G.D.; Haynes, B.F. HIV-1-specific antibody responses during acute and chronic HIV-1 infection. Curr. Opin. HIV AIDS 2009, 4, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.M.; Johnson, E.I.; Croom, H.A.; Richards, K.M.; Doughty, L.; Cunningham, P.H.; Kemp, B.E.; Branson, B.M.; Dax, E.M. Incidence immunoassay for distinguishing recent from established HIV-1 infection in therapy-naive populations. AIDS 2004, 18, 2253–2259. [Google Scholar] [CrossRef] [PubMed]
- McDougal, J.S.; Kennedy, M.S.; Nicholson, J.K.; Spira, T.J.; Jaffe, H.W.; Kaplan, J.E.; Fishbein, D.B.; O’Malley, P.; Aloisio, C.H.; Black, C.M.; et al. Antibody response to human immunodeficiency virus in homosexual men. Relation of antibody specificity, titer, and isotype to clinical status, severity of immunodeficiency, and disease progression. J. Clin. Investig. 1987, 80, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Beniguel, L.; Begaud, E.; Peruchon, S.; Cognasse, F.; Gabrie, P.; Marovich, M.; Lucht, F.; Genin, C.; Garraud, O. Isotype profiles of anti-gp160 antibodies from HIV-infected patients in plasma and culture supernatants. Immunol. Lett. 2004, 93, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Curtis, K.A.; Ambrose, K.M.; Kennedy, M.S.; Owen, S.M. Evaluation of dried blood spots with a multiplex assay for measuring recent HIV-1 infection. PLoS ONE 2014, 9, e107153. [Google Scholar] [CrossRef] [PubMed]
- Tinguely, C.; Schild-Spycher, T.; Bahador, Z.; Gowland, P.; Stolz, M.; Niederhauser, C. Comparison of a conventional HIV 1/2 line immunoassay with a rapid confirmatory HIV 1/2 assay. J. Virol. Methods 2014, 206, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Schupbach, J.; Gebhardt, M.D.; Tomasik, Z.; Niederhauser, C.; Yerly, S.; Burgisser, P.; Matter, L.; Gorgievski, M.; Dubs, R.; Schultze, D.; et al. Assessment of recent HIV-1 infection by a line immunoassay for HIV-1/2 confirmation. PLoS Med. 2007, 4, e343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendel, S.K.; Mullis, C.E.; Eshleman, S.H.; Blankson, J.N.; Moore, R.D.; Keruly, J.C.; Brookmeyer, R.; Quinn, T.C.; Laeyendecker, O. Effect of natural and ARV-induced viral suppression and viral breakthrough on anti-HIV antibody proportion and avidity in patients with HIV-1 subtype B infection. PLoS ONE 2013, 8, e55525. [Google Scholar] [CrossRef] [PubMed]
- Rawal, B.D.; Degula, A.; Lebedeva, L.; Janssen, R.S.; Hecht, F.M.; Sheppard, H.W.; Busch, M.P. Development of a new less-sensitive enzyme immunoassay for detection of early HIV-1 infection. J. Acquir. Immune Defic. Syndr. 2003, 33, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Burgers, W.A.; Riou, C.; Mlotshwa, M.; Maenetje, P.; de Assis Rosa, D.; Brenchley, J.; Mlisana, K.; Douek, D.C.; Koup, R.; Roederer, M.; et al. Association of HIV-specific and total CD8+ T memory phenotypes in subtype C HIV-1 infection with viral set point. J. Immunol. 2009, 182, 4751–4761. [Google Scholar] [CrossRef] [PubMed]
- Laeyendecker, O.; Brookmeyer, R.; Mullis, C.; Donnell, D.; Lingappa, J.; Celum, C.; Baeten, J.M.; Campbell, M.S.; Essex, M.; de Bruyn, G.; et al. Specificity of four laboratory approaches for cross-sectional HIV incidence determination: Analysis of samples from adults with known non-recent HIV infection from five African countries. AIDS Res. Hum. Retrovir. 2012, 28, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Hallett, T.B.; Ghys, P.; Barnighausen, T.; Yan, P.; Garnett, G.P. Errors in “BED”-derived estimates of HIV incidence will vary by place, time and age. PLoS ONE 2009, 4, e5720. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS. WHO/UNAIDS Technical Update on HIV Incidence Assays for Surveillance and Epidemic Monitoring; UNAIDS: Geneva, Swtizerland, 2013. [Google Scholar]
- Rehle, T.; Johnson, L.; Hallett, T.; Mahy, M.; Kim, A.; Odido, H.; Onoya, D.; Jooste, S.; Shisana, O.; Puren, A.; et al. A comparison of South African national HIV incidence estimates: A critical appraisal of different methods. PLoS ONE 2015, 10, e0133255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousins, M.M.; Laeyendecker, O.; Beauchamp, G.; Brookmeyer, R.; Towler, W.I.; Hudelson, S.E.; Khaki, L.; Koblin, B.; Chesney, M.; Moore, R.D. Use of a high resolution melting (HRM) assay to compare gag, pol, and env diversity in adults with different stages of HIV infection. PLoS ONE 2011, 6, e27211. [Google Scholar] [CrossRef] [PubMed]
- Cousins, M.M.; Konikoff, J.; Laeyendecker, O.; Celum, C.; Buchbinder, S.P.; Seage, G.R., 3rd; Kirk, G.D.; Moore, R.D.; Mehta, S.H.; Margolick, J.B.; et al. HIV diversity as a biomarker for HIV incidence estimation: including a high resolution melting diversity assay in a multi-assay algorithm. J. Clin. Microbiol. 2013, 52, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Cousins, M.M.; Donnell, D.; Eshleman, S.H. Impact of mutation type and amplicon characteristics on genetic diversity measures generated using a high-resolution melting diversity assay. J. Mol. Diagn. 2013, 15, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Meixenberger, K.; Hauser, A.; Jansen, K.; Yousef, K.P.; Fiedler, S.; von Kleist, M.; Norley, S.; Somogyi, S.; Hamouda, O.; Bannert, N.; et al. Assessment of ambiguous base calls in HIV-1 pol population sequences as a biomarker for identification of recent infections in HIV-1 incidence studies. J. Clin. Microbiol. 2014, 52, 2977–2983. [Google Scholar] [CrossRef] [PubMed]
- Andersson, E.; Shao, W.; Bontell, I.; Cham, F.; Cuong, D.D.; Wondwossen, A.; Morris, L.; Hunt, G.; Sönnerborg, A.; Bertagnolio, S. Evaluation of sequence ambiguities of the HIV-1 pol gene as a method to identify recent HIV-1 infection in transmitted drug resistance surveys. Infect. Genet. Evol. 2013, 18, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Volz, E.M.; Ionides, E.; Romero-Severson, E.O.; Brandt, M.-G.; Mokotoff, E.; Koopman, J.S. HIV-1 transmission during early infection in men who have sex with men: A phylodynamic analysis. PLoS Med. 2013, 10, e1001568. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Love, T.M.; Nelson, J.; Thurston, S.W.; Perelson, A.S.; Lee, H.Y. Designing a genome-based HIV incidence assay with high sensitivity and specificity. AIDS 2011, 25, F13–F19. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.-Y.; Ge, M.; Hsi, J.H.; He, X.; Ruan, Y.-H.; Wang, Z.-X.; Shao, Y.-M.; Pan, X.-M. High-accuracy identification of incident HIV-1 infections using a sequence clustering based diversity measure. PLoS ONE 2014, 9, e100081. [Google Scholar] [CrossRef] [PubMed]
- Laeyendecker, O.; Rothman, R.E.; Henson, C.; Horne, B.J.; Ketlogetswe, K.S.; Kraus, C.K.; Shahan, J.; Kelen, G.D.; Quinn, T.C. The effect of viral suppression on cross-sectional incidence testing in the Johns Hopkins hospital emergency department. J. Acquir. Immune Defic. Syndr. 2008, 48, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Konikoff, J.; Brookmeyer, R.; Longosz, A.F.; Cousins, M.M.; Celum, C.; Buchbinder, S.P.; Seage, G.R., 3rd; Kirk, G.D.; Moore, R.D.; Mehta, S.H.; et al. Performance of a limiting-antigen avidity enzyme immunoassay for cross-sectional estimation of HIV incidence in the United States. PLoS ONE 2013, 8, e82772. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.B.; Swanstrom, R.; Kashuba, A.D.; Cohen, M.S. Bottlenecks in HIV-1 transmission: Insights from the study of founder viruses. Nat. Rev. Microbiol. 2015, 13, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 2008, 105, 7552–7557. [Google Scholar] [CrossRef] [PubMed]
- Parker, Z.F.; Iyer, S.S.; Wilen, C.B.; Parrish, N.F.; Chikere, K.C.; Lee, F.-H.; Didigu, C.A.; Berro, R.; Klasse, P.J.; Lee, B.; et al. Transmitted/founder and chronic HIV-1 envelope proteins are distinguished by differential utilization of CCR5. J. Virol. 2013, 87, 2401–2411. [Google Scholar] [CrossRef] [PubMed]
- Parrish, N.F.; Wilen, C.B.; Banks, L.B.; Iyer, S.S.; Pfaff, J.M.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Decker, J.M.; Parrish, E.H.; Berg, A.; et al. Transmitted/founder and chronic subtype C HIV-1 use CD4 and CCR5 receptors with equal efficiency and are not inhibited by blocking the integrin α4β7. PLoS Pathog. 2012, 8, e1002686. [Google Scholar] [CrossRef] [PubMed]
- Novitsky, V.; Lagakos, S.; Herzig, M.; Bonney, C.; Kebaabetswe, L.; Rossenkhan, R.; Nkwe, D.; Margolin, L.; Musonda, R.; Moyo, S. Evolution of proviral gp120 over the first year of HIV-1 subtype C infection. Virology 2009, 383, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Kearney, M.; Maldarelli, F.; Shao, W.; Margolick, J.B.; Daar, E.S.; Mellors, J.W.; Rao, V.; Coffin, J.M.; Palmer, S. Human immunodeficiency virus type 1 population genetics and adaptation in newly infected individuals. J. Virol. 2009, 83, 2715–2727. [Google Scholar] [CrossRef] [PubMed]
- Palmer, S.; Kearney, M.; Maldarelli, F.; Halvas, E.K.; Bixby, C.J.; Bazmi, H.; Rock, D.; Falloon, J.; Davey, R.T., Jr.; Dewar, R.L.; et al. Multiple, linked human immunodeficiency virus type 1 drug resistance mutations in treatment-experienced patients are missed by standard genotype analysis. J. Clin. Microbiol. 2005, 43, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Cousins, M.M.; Ou, S.S.; Wawer, M.J.; Munshaw, S.; Swan, D.; Magaret, C.A.; Mullis, C.E.; Serwadda, D.; Porcella, S.F.; Gray, R.H.; et al. Comparison of a high-resolution melting assay to next-generation sequencing for analysis of HIV diversity. J. Clin. Microbiol. 2012, 50, 3054–3059. [Google Scholar] [CrossRef] [PubMed]
- Towler, W.I.; James, M.M.; Ray, S.C.; Wang, L.; Donnell, D.; Mwatha, A.; Guay, L.; Nakabiito, C.; Musoke, P.; Jackson, J.B.; et al. Analysis of HIV diversity using a high-resolution melting assay. AIDS Res. Hum. Retrovir. 2010, 26, 913–918. [Google Scholar] [CrossRef] [PubMed]
- James, M.M.; Wang, L.; Donnell, D.; Cousins, M.M.; Barlow-Mosha, L.; Fogel, J.M.; Towler, W.I.; Agwu, A.L.; Bagenda, D.; Mubiru, M.; et al. Use of a high resolution melting assay to analyze HIV diversity in HIV-infected Ugandan children. Pediatr. Infect. Dis. J. 2012, 31, e222–e228. [Google Scholar] [CrossRef] [PubMed]
- Delwart, E.L.; Sheppard, H.W.; Walker, B.D.; Goudsmit, J.; Mullins, J.I. Human immunodeficiency virus type 1 evolution in vivo tracked by DNA heteroduplex mobility assays. J. Virol. 1994, 68, 6672–6683. [Google Scholar] [PubMed]
- Cousins, M.M.; Konikoff, J.; Sabin, D.; Khaki, L.; Longosz, A.F.; Laeyendecker, O.; Celum, C.; Buchbinder, S.P.; Seage, G.R., 3rd; Kirk, G.D.; et al. A comparison of two measures of HIV diversity in multi-assay algorithms for HIV incidence estimation. PLoS ONE 2014, 9, e101043. [Google Scholar] [CrossRef] [PubMed]
- Shankarappa, R.; Margolick, J.B.; Gange, S.J.; Rodrigo, A.G.; Upchurch, D.; Farzadegan, H.; Gupta, P.; Rinaldo, C.R.; Learn, G.H.; He, X.; et al. Consistent viral evolutionary changes associated with the progression of human immunodeficiency virus type 1 infection. J. Virol. 1999, 73, 10489–10502. [Google Scholar] [PubMed]
- Lee, H.Y.; Perelson, A.S.; Park, S.-C.; Leitner, T. Dynamic correlation between intrahost HIV-1 quasispecies evolution and disease progression. PLoS Comput. Biol. 2008, 4, e1000240. [Google Scholar] [CrossRef] [PubMed]
- Wolbers, M.; Babiker, A.; Sabin, C.; Young, J.; Dorrucci, M.; Chêne, G.; Mussini, C.; Porter, K.; Bucher, H.C. Pretreatment CD4 cell slope and progression to AIDS or death in HIV-infected patients initiating antiretroviral therapy—The CASCADE collaboration: A collaboration of 23 cohort studies. PLoS Med. 2010, 7, e1000239. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Goeken, N.; Lee, H.J.; Bolan, R.; Dube, M.P.; Lee, H.Y. Developing high-throughput HIV incidence assay with pyrosequencing platform. J. Virol. 2014, 88, 2977–2990. [Google Scholar] [CrossRef] [PubMed]
- Stramer, S.L.; Glynn, S.A.; Kleinman, S.H.; Strong, D.M.; Sally, C.; Wright, D.J.; Dodd, R.Y.; Busch, M.P. Detection of HIV-1 and HCV infections among antibody-negative blood donors by nucleic acid-amplification testing. N. Engl. J. Med. 2004, 351, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Hallett, T.B. Estimating the HIV incidence rate: Recent and future developments. Curr. Opin. HIV AIDS 2011, 6, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xia, X.; He, X.; Yang, S.; Ruan, Y.; Zhao, Q.; Wang, Z.; Shao, Y.; Pan, X. A new pattern-based method for identifying recent HIV-1 infections from the viral env sequence. Sci. Chin. Life Sci. 2012, 55, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Ypma, R.J.; van Ballegooijen, W.M.; Wallinga, J. Relating phylogenetic trees to transmission trees of infectious disease outbreaks. Genetics 2013, 195, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Eshleman, S.H.; Hudelson, S.E.; Redd, A.D.; Wang, L.; Debes, R.; Chen, Y.Q.; Martens, C.A.; Ricklefs, S.M.; Selig, E.J.; Porcella, S.F.; et al. Analysis of genetic linkage of HIV from couples enrolled in the HIV Prevention Trials Network 052 trial. J. Infect. Dis. 2011, 204, 1918–1926. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.S.; Chen, Y.Q.; McCauley, M.; Gamble, T.; Hosseinipour, M.C.; Kumarasamy, N.; Hakim, J.G.; Kumwenda, J.; Grinsztejn, B.; Pilotto, J.H.; et al. Prevention of HIV-1 infection with early antiretroviral therapy. N. Engl. J. Med. 2011, 365, 493–505. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moyo, S.; Wilkinson, E.; Novitsky, V.; Vandormael, A.; Gaseitsiwe, S.; Essex, M.; Engelbrecht, S.; De Oliveira, T. Identifying Recent HIV Infections: From Serological Assays to Genomics. Viruses 2015, 7, 5508-5524. https://doi.org/10.3390/v7102887
Moyo S, Wilkinson E, Novitsky V, Vandormael A, Gaseitsiwe S, Essex M, Engelbrecht S, De Oliveira T. Identifying Recent HIV Infections: From Serological Assays to Genomics. Viruses. 2015; 7(10):5508-5524. https://doi.org/10.3390/v7102887
Chicago/Turabian StyleMoyo, Sikhulile, Eduan Wilkinson, Vladimir Novitsky, Alain Vandormael, Simani Gaseitsiwe, Max Essex, Susan Engelbrecht, and Tulio De Oliveira. 2015. "Identifying Recent HIV Infections: From Serological Assays to Genomics" Viruses 7, no. 10: 5508-5524. https://doi.org/10.3390/v7102887
APA StyleMoyo, S., Wilkinson, E., Novitsky, V., Vandormael, A., Gaseitsiwe, S., Essex, M., Engelbrecht, S., & De Oliveira, T. (2015). Identifying Recent HIV Infections: From Serological Assays to Genomics. Viruses, 7(10), 5508-5524. https://doi.org/10.3390/v7102887