
Incorporation of Spike and Membrane Glycoproteins into Coronavirus Virions

Abstract
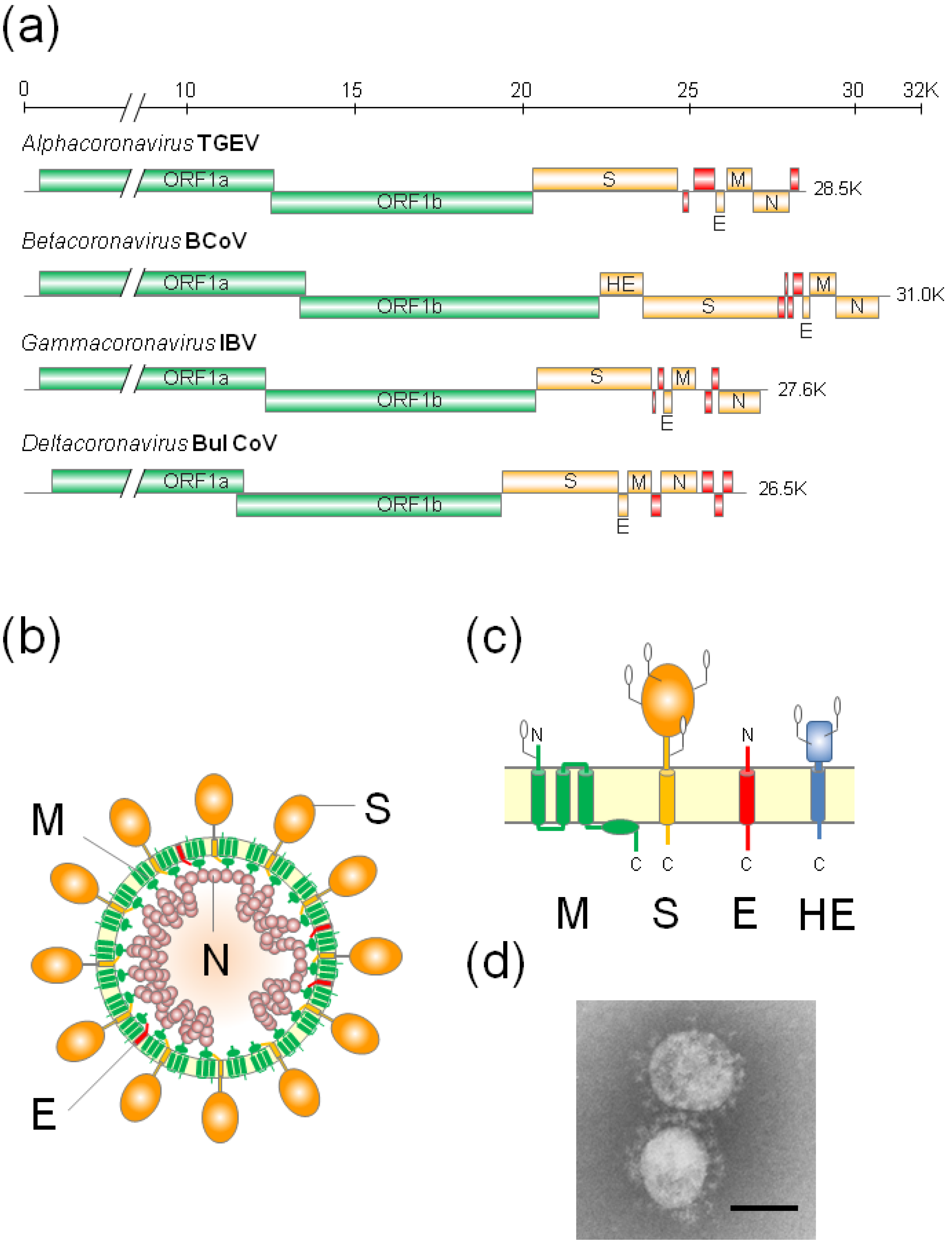
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
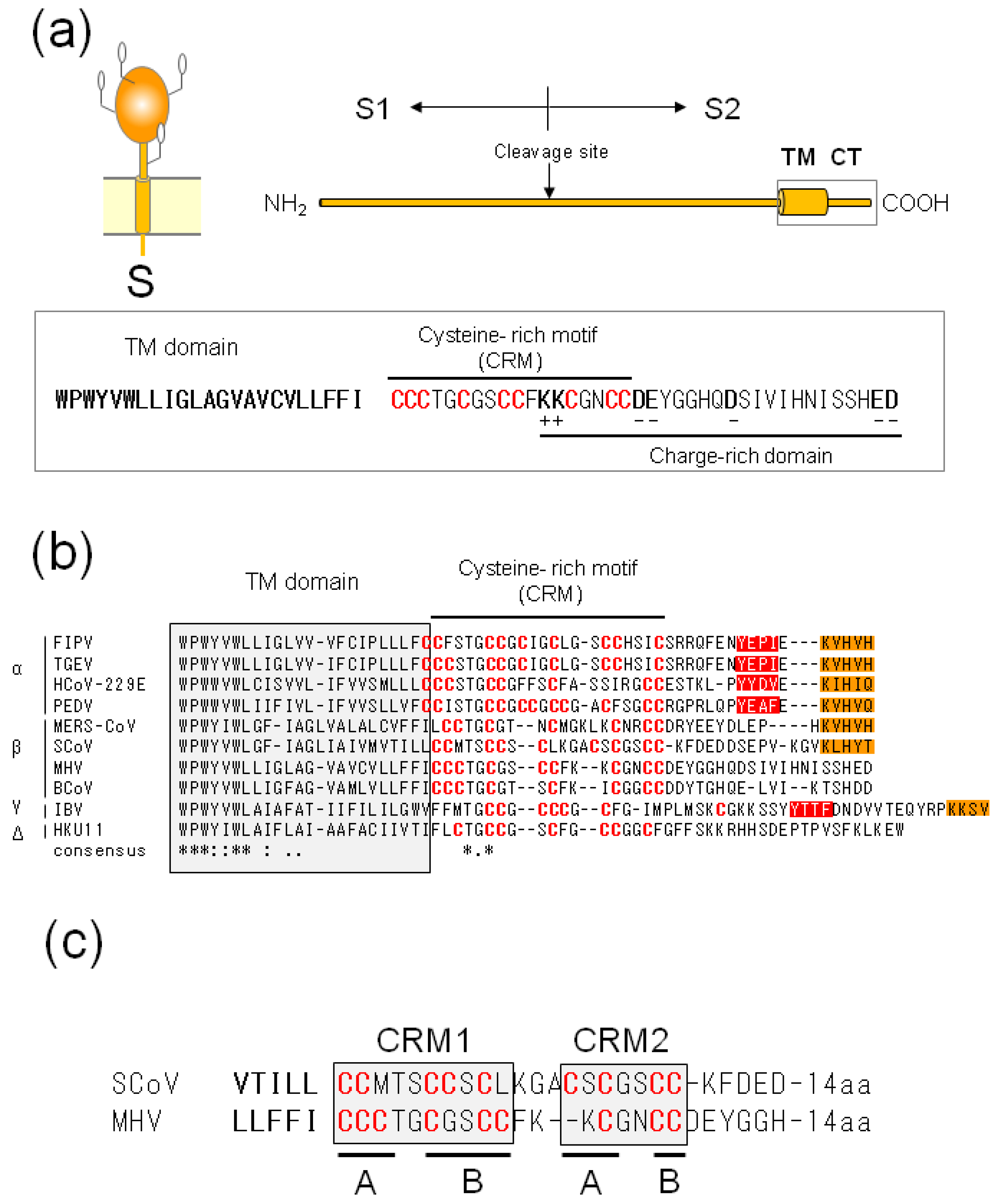{kind=link}
| Genus | Species |
|---|---|
| Alphacoronavirus | Feline Coronavirus (FCoV) |
| Transmissible Gastroenteritis Virus (TGEV) | |
| Porcine Epidemic Diarrhea Coronavirus (PEDV) | |
| Human Coronavirus 229E | |
| Human Coronavirus NL63 | |
| Betacoronavirus | Bovine Coronavirus (BCoV) |
| Mouse Hepatitis Virus (MHV) | |
| Human Coronavirus OC43 | |
| Human Coronavirus HKU-1 | |
| Severe Acute Respiratory Syndrome Coronavirus (SCoV) | |
| Middle East respiratory Syndrome Coronavirus (MERS-CoV) | |
| Gammacoronavirus | Infectious Bronchitis Virus (IBV) |
| Deltacoronavirus | Bulbul Coronavirus HKU11 |

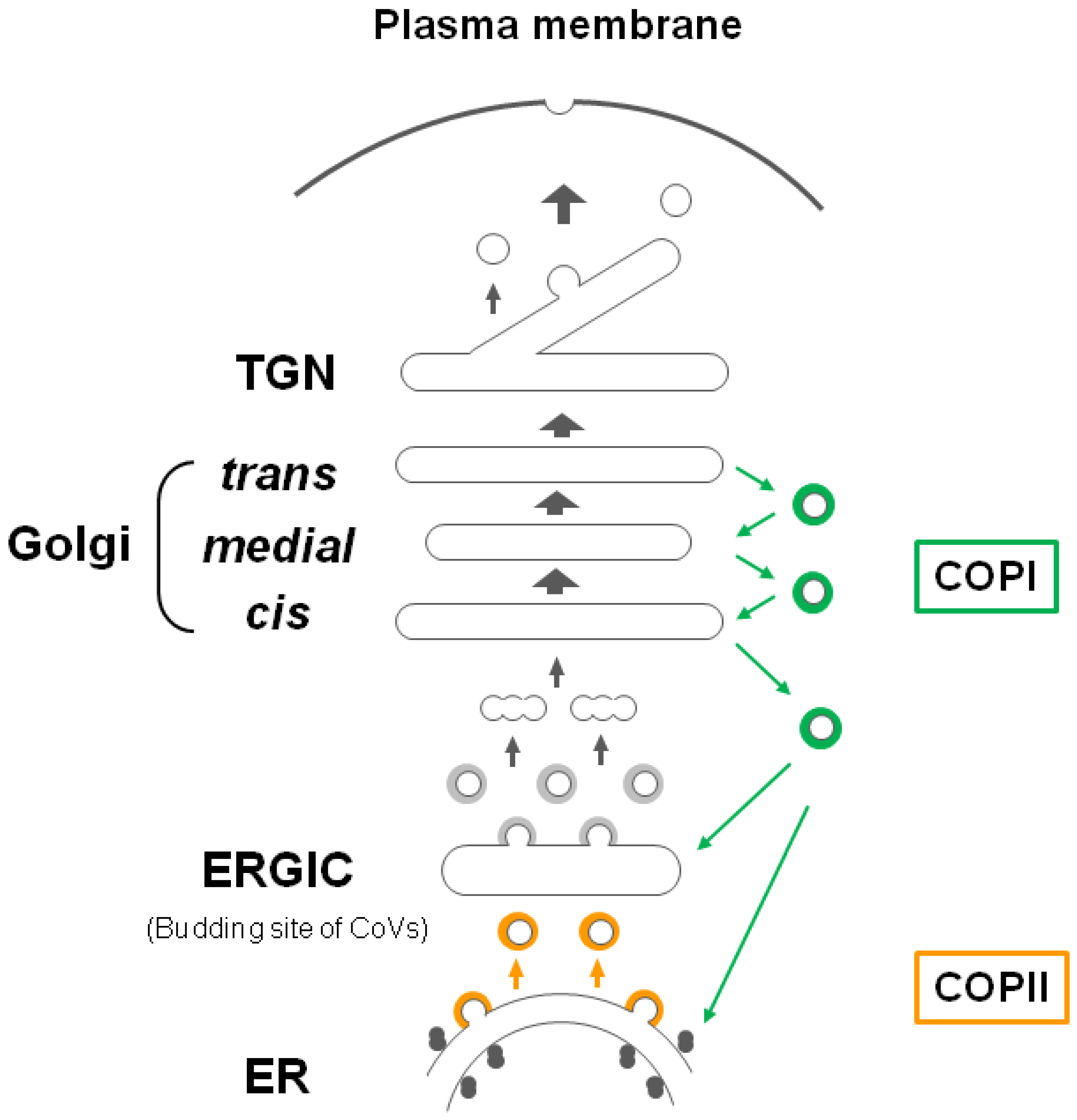
2. The Glycoprotein Trafficking and Intracellular Retention Mechanism

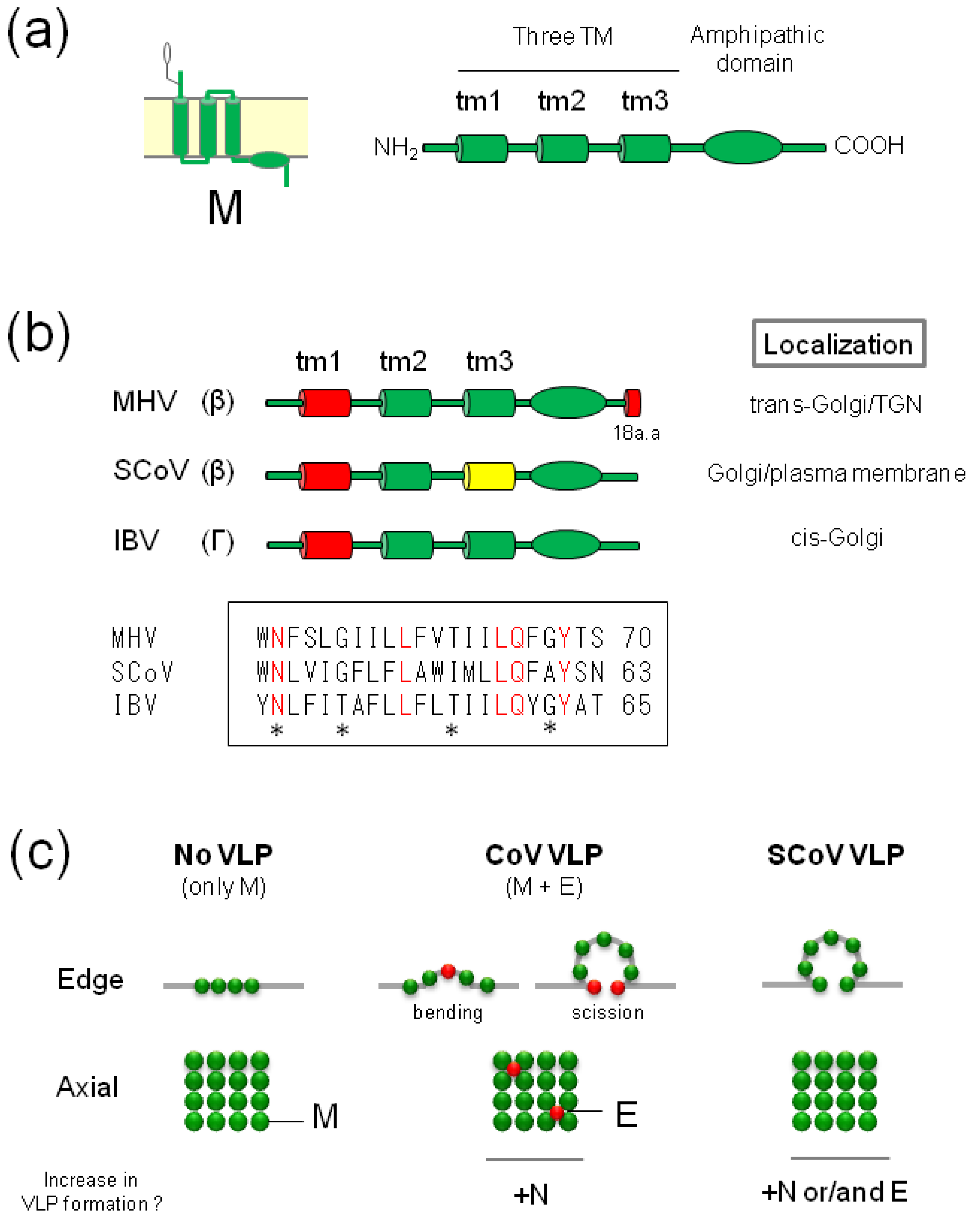
3. M Proteins
3.1. General Properties
3.2. Golgi Retention Signal of CoV-M Proteins


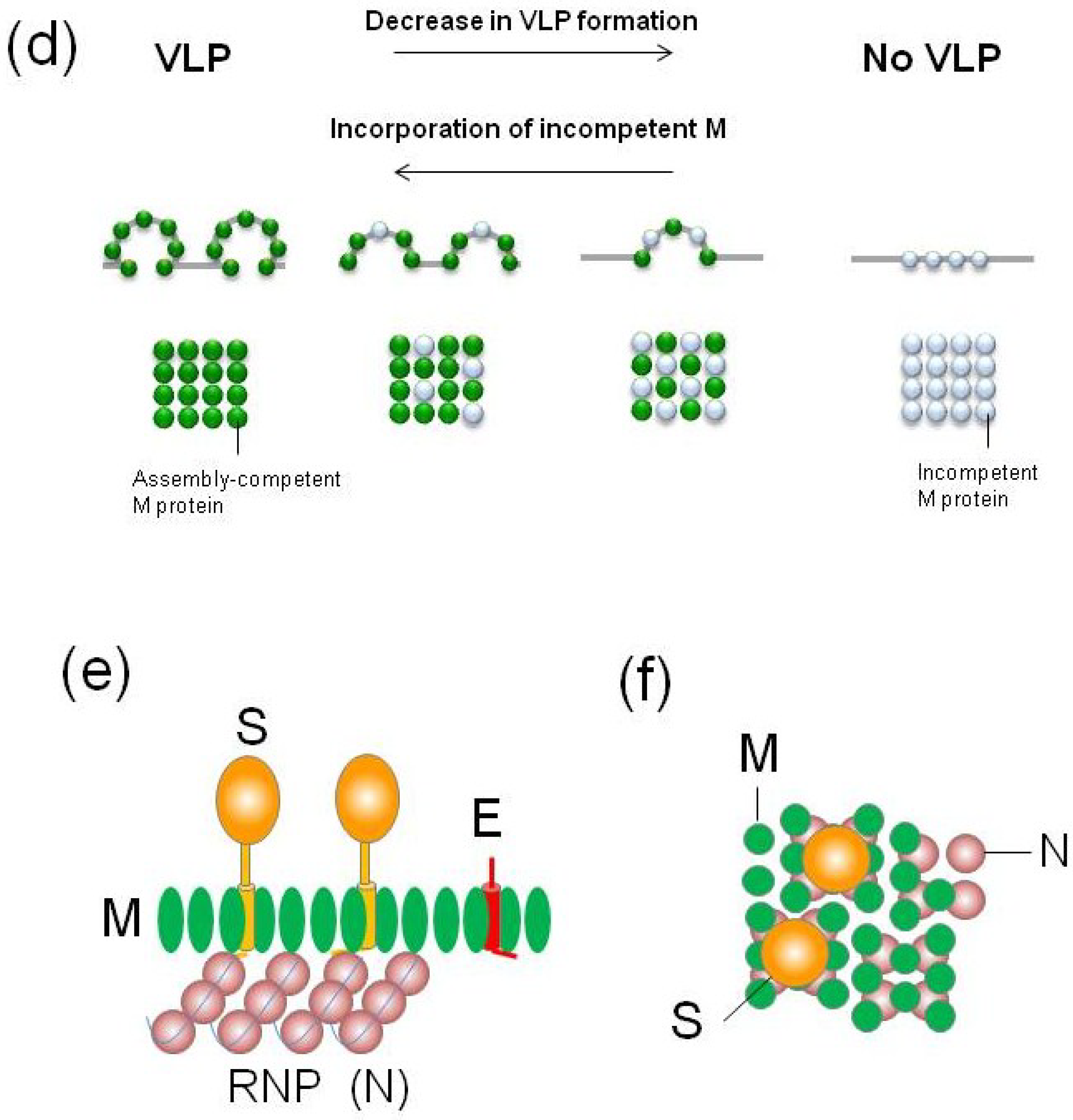
3.3. Incorporation of CoV-M into VLPs or Virions
4. S Proteins
4.1. General Properties
4.2. Intracellular Retention Signal of CoV-S Proteins

| Genus | Species | Tyrosine-Dependent Localization/Internalization Signals (I.S.) | ER Retrieval Signal |
|---|---|---|---|
| Alpha CoV | TGEV | YxxI motif Main retention signal. Not I.S. [109] | KxHxx motif Not main retention signal [109] Retention of reporter protein [38] |
| PEDV | YxxF motif Not studied | KxHxx motif Retention signal [114] | |
| Beta CoV | SCoV | KxHxx motif Weak retention signal [110] | |
| Gamma CoV | IBV | YxxF motif Main retention signal. Not I.S. [111] I.S. Not main retention signal [38] | KKxx motif Main retention signal [38] Not main retention signal [111] |
4.3. Incorporation of CoV-S Proteins into VLPs or Virions
| Genus | Species | M-S Interaction | VLP/Virion Incorporation (or Infectivity **) | |
|---|---|---|---|---|
| Palmitoylation | Position specificity | |||
| Alpha CoV | TGEV | Independent [127] | Dependent [127] | No specific position [127] |
| Beta CoV | MHV | Dependent [123,124] | Dependent [123,124] | Specific position (CRM1B) ** [123] Specific position (CRM2) [124] No specific position [125] ** |
| SCoV | Independent [122,126] | Dependent [122] | Specific position (CRM2B)[122] | |
5. Conclusions and Perspective
Acknowledgments
Conflicts of Interest
References
- Hamre, D.; Procknow, J.J. A new virus isolated from the human respiratory tract. Proc. Soc. Exp. Biol. Med. 1966, 121, 190–193. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, K.; Dees, J.H.; Becker, W.B.; Kapikian, A.Z.; Chanock, R.M. Recovery in tracheal organ cultures of novel viruses from patients with respiratory disease. Proc. Natl. Acad. Sci. USA 1967, 57, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, D.A.; Bynoe, M.L. Cultivation of a novel type of common-cold virus in organ cultures. Br. Med. J. 1965, 1, 1467–1470. [Google Scholar] [CrossRef] [PubMed]
- van der Hoek, L.; Pyrc, K.; Jebbink, M.F.; Vermeulen-Oost, W.; Berkhout, R.J.; Wolthers, K.C.; Wertheim-van Dillen, P.M.; Kaandorp, J.; Spaargaren, J.; Berkhout, B. Identification of a new human coronavirus. Nat. Med. 2004, 10, 368–373. [Google Scholar]
- Woo, P.C.; Lau, S.K.; Tsoi, H.W.; Huang, Y.; Poon, R.W.; Chu, C.M.; Lee, R.A.; Luk, W.K.; Wong, G.K.; Wong, B.H.; et al. Clinical and molecular epidemiological features of coronavirus hku1-associated community-acquired pneumonia. J. Infect. Dis. 2005, 192, 1898–1907. [Google Scholar] [CrossRef] [Green Version]
- Pyrc, K.; Berkhout, B.; van der Hoek, L. The novel human coronaviruses nl63 and hku1. J. Virol. 2007, 81, 3051–3057. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Gunther, S.; Preiser, W.; van der Werf, S.; Brodt, H.R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.; et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef]
- Fouchier, R.A.; Kuiken, T.; Schutten, M.; van Amerongen, G.; van Doornum, G.J.; van den Hoogen, B.G.; Peiris, M.; Lim, W.; Stohr, K.; Osterhaus, A.D. Aetiology: Koch’s postulates fulfilled for SARS virus. Nature 2003, 423, 240. [Google Scholar] [CrossRef] [PubMed]
- Ksiazek, T.G.; Erdman, D.; Goldsmith, C.S.; Zaki, S.R.; Peret, T.; Emery, S.; Tong, S.; Urbani, C.; Comer, J.A.; Lim, W.; et al. A novel coronavirus associated with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1953–1966. [Google Scholar] [CrossRef]
- Kuiken, T.; Fouchier, R.A.; Schutten, M.; Rimmelzwaan, G.F.; van Amerongen, G.; van Riel, D.; Laman, J.D.; de Jong, T.; van Doornum, G.; Lim, W.; et al. Newly discovered coronavirus as the primary cause of severe acute respiratory syndrome. Lancet 2003, 362, 263–270. [Google Scholar] [CrossRef]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in saudi arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Middle East Respiratory Syndrome Coronavirus (Mers-cov)—Saudi Arabia. Available online: http://www.who.int/csr/don/07-november-2014-mers/en/ (accessed on 1 December 2014).
- Enjuanes, L.; Zeijst, B.A.M.V.D. Molecular basis of transmissible gastroenteritis virus epidemiology. In The Coronaviridae; Siddell, S.G., Ed.; Plenum Press: New York, NY, USA, 1995; pp. 337–376. [Google Scholar]
- Huang, Y.W.; Dickerman, A.W.; Pineyro, P.; Li, L.; Fang, L.; Kiehne, R.; Opriessnig, T.; Meng, X.J. Origin, evolution, and genotyping of emergent porcine epidemic diarrhea virus strains in the united states. MBio 2013, 4, e00737–e00713. [Google Scholar] [CrossRef] [PubMed]
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Sjaak de Wit, J.J.; Cook, J.K.; van der Heijden, H.M. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef] [PubMed]
- OIE. Oie-Listed Diseases, Infections and Infestations in Force in 2014. Available online: http://www.oie.int/animal-health-in-the-world/oie-listed-diseases-2014/ (accessed on 1 December 2014).
- De Groot, R.J.; Baker, S.C.; Baric, R.; Enjuanes, L.; Gorbalenya, A.E.; Holmes, K.V.; Perlman, S.; Poon, L.; Rottier, P.J.M.; Talbot, P.J.; et al. Family Coronaviridae; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Woo, P.C.; Lau, S.K.; Lam, C.S.; Lau, C.C.; Tsang, A.K.; Lau, J.H.; Bai, R.; Teng, J.L.; Tsang, C.C.; Wang, M.; et al. Discovery of seven novel mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Brian, D.A.; Baric, R.S. Coronavirus genome structure and replication. Curr. Top. Microbiol. Immunol. 2005, 287, 1–30. [Google Scholar] [PubMed]
- Namy, O.; Moran, S.J.; Stuart, D.I.; Gilbert, R.J.; Brierley, I. A mechanical explanation of rna pseudoknot function in programmed ribosomal frameshifting. Nature 2006, 441, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Ziebuhr, J.; Snijder, E.J.; Gorbalenya, A.E. Virus-encoded proteinases and proteolytic processing in the nidovirales. J. Gen. Virol. 2000, 81, 853–879. [Google Scholar] [PubMed]
- Gorbalenya, A.E.; Enjuanes, L.; Ziebuhr, J.; Snijder, E.J. Nidovirales: Evolving the largest rna virus genome. Virus. Res. 2006, 117, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Lissenberg, A.; Vrolijk, M.M.; van Vliet, A.L.; Langereis, M.A.; de Groot-Mijnes, J.D.; Rottier, P.J.; de Groot, R.J. Luxury at a cost? Recombinant mouse hepatitis viruses expressing the accessory hemagglutinin esterase protein display reduced fitness in vitro. J. Virol. 2005, 79, 15054–15063. [Google Scholar] [CrossRef] [PubMed]
- Vennema, H.; Godeke, G.J.; Rossen, J.W.; Voorhout, W.F.; Horzinek, M.C.; Opstelten, D.J.; Rottier, P.J. Nucleocapsid-independent assembly of coronavirus-like particles by co-expression of viral envelope protein genes. EMBO. J. 1996, 15, 2020–2028. [Google Scholar] [PubMed]
- DeDiego, M.L.; Alvarez, E.; Almazan, F.; Rejas, M.T.; Lamirande, E.; Roberts, A.; Shieh, W.J.; Zaki, S.R.; Subbarao, K.; Enjuanes, L. A severe acute respiratory syndrome coronavirus that lacks the E gene is attenuated in vitro and in vivo. J. Virol. 2007, 81, 1701–1713. [Google Scholar] [CrossRef] [PubMed]
- Kuo, L.; Masters, P.S. The small envelope protein E is not essential for murine coronavirus replication. J. Virol. 2003, 77, 4597–4608. [Google Scholar] [CrossRef] [PubMed]
- Fischer, F.; Stegen, C.F.; Masters, P.S.; Samsonoff, W.A. Analysis of constructed E gene mutants of mouse hepatitis virus confirms a pivotal role for E protein in coronavirus assembly. J. Virol. 1998, 72, 7885–7894. [Google Scholar] [PubMed]
- Baudoux, P.; Carrat, C.; Besnardeau, L.; Charley, B.; Laude, H. Coronavirus pseudoparticles formed with recombinant m and E proteins induce alpha interferon synthesis by leukocytes. J. Virol. 1998, 72, 8636–8643. [Google Scholar] [PubMed]
- Heald-Sargent, T.; Gallagher, T. Ready, set, fuse! The coronavirus spike protein and acquisition of fusion competence. Viruses 2012, 4, 557–580. [Google Scholar] [CrossRef] [PubMed]
- Li, F. Receptor recognition and cross-species infections of SARS coronavirus. Antiviral Res. 2013, 100, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [PubMed]
- Desforges, M.; Desjardins, J.; Zhang, C.; Talbot, P.J. The acetyl-esterase activity of the hemagglutinin-esterase protein of human coronavirus oc43 strongly enhances the production of infectious virus. J. Virol. 2013, 87, 3097–3107. [Google Scholar] [CrossRef] [PubMed]
- Stertz, S.; Reichelt, M.; Krijnse-Locker, J.; Mackenzie, J.; Simpson, J.C.; Haller, O.; Kochs, G. Interferon-induced, antiviral human mxa protein localizes to a distinct subcompartment of the smooth endoplasmic reticulum. J. Interferon Cytokine Res. 2006, 26, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Tooze, J.; Tooze, S.; Warren, G. Replication of coronavirus mhv-a59 in sac- cells: Determination of the first site of budding of progeny virions. Eur. J. Cell Biol. 1984, 33, 281–293. [Google Scholar] [PubMed]
- Krijnse-Locker, J.; Ericsson, M.; Rottier, P.J.; Griffiths, G. Characterization of the budding compartment of mouse hepatitis virus: Evidence that transport from the rer to the golgi complex requires only one vesicular transport step. J. Cell Biol. 1994, 124, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Lontok, E.; Corse, E.; Machamer, C.E. Intracellular targeting signals contribute to localization of coronavirus spike proteins near the virus assembly site. J. Virol. 2004, 78, 5913–5922. [Google Scholar] [CrossRef] [PubMed]
- Corse, E.; Machamer, C.E. Infectious bronchitis virus E protein is targeted to the golgi complex and directs release of virus-like particles. J. Virol. 2000, 74, 4319–4326. [Google Scholar] [CrossRef] [PubMed]
- Machamer, C.E.; Rose, J.K. A specific transmembrane domain of a coronavirus e1 glycoprotein is required for its retention in the golgi region. J. Cell Biol. 1987, 105, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.K.; Chang, S.C.; Huang, C.C.; Lee, T.T.; Hsiao, C.W.; Kou, Y.H.; Chen, I.Y.; Chang, C.K.; Huang, T.H.; Chang, M.F. Assembly of severe acute respiratory syndrome coronavirus rna packaging signal into virus-like particles is nucleocapsid dependent. J. Virol. 2005, 79, 13848–13855. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, Z.Y.; Kong, W.P.; Nabel, G.J. Generation of synthetic severe acute respiratory syndrome coronavirus pseudoparticles: Implications for assembly and vaccine production. J. Virol. 2004, 78, 12557–12565. [Google Scholar] [CrossRef] [PubMed]
- Siu, Y.L.; Teoh, K.T.; Lo, J.; Chan, C.M.; Kien, F.; Escriou, N.; Tsao, S.W.; Nicholls, J.M.; Altmeyer, R.; Peiris, J.S.; et al. The M, E, and N structural proteins of the severe acute respiratory syndrome coronavirus are required for efficient assembly, trafficking, and release of virus-like particles. J. Virol. 2008, 82, 11318–11330. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.T.; Wang, S.M.; Huang, K.J.; Lee, A.I.; Chiang, C.C.; Wang, C.T. Self-assembly of severe acute respiratory syndrome coronavirus membrane protein. J. Biol. Chem. 2010, 285, 12862–12872. [Google Scholar] [CrossRef] [PubMed]
- De Haan, C.A.; Kuo, L.; Masters, P.S.; Vennema, H.; Rottier, P.J. Coronavirus particle assembly: Primary structure requirements of the membrane protein. J. Virol. 1998, 72, 6838–6850. [Google Scholar] [PubMed]
- De Haan, C.A.; Vennema, H.; Rottier, P.J. Assembly of the coronavirus envelope: Homotypic interactions between the M proteins. J. Virol. 2000, 74, 4967–4978. [Google Scholar] [CrossRef] [PubMed]
- Masters, P.S.; Kuo, L.; Ye, R.; Hurst, K.R.; Koetzner, C.A.; Hsue, B. Genetic and molecular biological analysis of protein-protein interactions in coronavirus assembly. Adv. Exp. Med. Biol. 2006, 581, 163–173. [Google Scholar] [PubMed]
- Nguyen, V.P.; Hogue, B.G. Protein interactions during coronavirus assembly. J. Virol. 1997, 71, 9278–9284. [Google Scholar] [PubMed]
- Opstelten, D.J.; Raamsman, M.J.; Wolfs, K.; Horzinek, M.C.; Rottier, P.J. Envelope glycoprotein interactions in coronavirus assembly. J. Cell Biol. 1995, 131, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, K.; Maeda, A.; Maeda, J.; Makino, S. Characterization of the coronavirus m protein and nucleocapsid interaction in infected cells. J. Virol. 2000, 74, 8127–8134. [Google Scholar] [CrossRef] [PubMed]
- Raamsman, M.J.; Locker, J.K.; de Hooge, A.; de Vries, A.A.; Griffiths, G.; Vennema, H.; Rottier, P.J. Characterization of the coronavirus mouse hepatitis virus strain a59 small membrane protein e. J. Virol. 2000, 74, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- De Haan, C.A.; Rottier, P.J. Molecular interactions in the assembly of coronaviruses. Adv. Virus. Res. 2005, 64, 165–230. [Google Scholar] [PubMed]
- Appenzeller-Herzog, C.; Hauri, H.P. The er-golgi intermediate compartment (ergic): In search of its identity and function. J. Cell. Sci. 2006, 119, 2173–2183. [Google Scholar] [CrossRef] [PubMed]
- Geva, Y.; Schuldiner, M. The back and forth of cargo exit from the endoplasmic reticulum. Curr. Biol. 2014, 24, R130–R136. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.S.; Luini, A. Models for golgi traffic: A critical assessment. Cold Spring Harb. Perspect. Biol. 2011, 3, a005215. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.S.; Nakano, A. Membrane traffic within the golgi apparatus. Annu. Rev. Cell Dev. Biol. 2009, 25, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Luini, A.; Mironov, A.A.; Polishchuk, E.V.; Polishchuk, R.S. Morphogenesis of post-golgi transport carriers. Histochem. Cell Biol. 2008, 129, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.L. Mechanisms of transport through the golgi complex. J. Cell. Sci. 2009, 122, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, T.; Warren, G. Retention and retrieval in the endoplasmic reticulum and the golgi apparatus. Curr. Opin. Cell Biol. 1994, 6, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Banfield, D.K. Mechanisms of protein retention in the golgi. Cold Spring Harb. Perspect. Biol. 2011, 3, a005264. [Google Scholar] [CrossRef] [PubMed]
- Ben-Tekaya, H.; Miura, K.; Pepperkok, R.; Hauri, H.P. Live imaging of bidirectional traffic from the ergic. J. Cell. Sci. 2005, 118, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, T.; Hoe, M.H.; Slusarewicz, P.; Rabouille, C.; Watson, R.; Hunte, F.; Watzele, G.; Berger, E.G.; Warren, G. Kin recognition between medial golgi enzymes in hela cells. EMBO. J. 1994, 13, 562–574. [Google Scholar] [PubMed]
- Nilsson, T.; Slusarewicz, P.; Hoe, M.H.; Warren, G. Kin recognition. A model for the retention of golgi enzymes. FEBS Lett. 1993, 330, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.; Niemann, H.; Smeekens, S.; Rottier, P.; Warren, G. Sequence and topology of a model intracellular membrane protein, e1 glycoprotein, from a coronavirus. Nature 1984, 308, 751–752. [Google Scholar] [CrossRef] [PubMed]
- Rottier, P.J.; Welling, G.W.; Welling-Wester, S.; Niesters, H.G.; Lenstra, J.A.; van der Zeijst, B.A. Predicted membrane topology of the coronavirus protein e1. Biochemistry 1986, 25, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Voss, D.; Pfefferle, S.; Drosten, C.; Stevermann, L.; Traggiai, E.; Lanzavecchia, A.; Becker, S. Studies on membrane topology, n-glycosylation and functionality of SARS-cov membrane protein. Virol. J. 2009, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Escors, D.; Camafeita, E.; Ortego, J.; Laude, H.; Enjuanes, L. Organization of two transmissible gastroenteritis coronavirus membrane protein topologies within the virion and core. J. Virol. 2001, 75, 12228–12240. [Google Scholar] [CrossRef] [PubMed]
- Risco, C.; Anton, I.M.; Sune, C.; Pedregosa, A.M.; Martin-Alonso, J.M.; Parra, F.; Carrascosa, J.L.; Enjuanes, L. Membrane protein molecules of transmissible gastroenteritis coronavirus also expose the carboxy-terminal region on the external surface of the virion. J. Virol. 1995, 69, 5269–5277. [Google Scholar] [PubMed]
- De Haan, C.A.; Roestenberg, P.; de Wit, M.; de Vries, A.A.; Nilsson, T.; Vennema, H.; Rottier, P.J. Structural requirements for o-glycosylation of the mouse hepatitis virus membrane protein. J. Biol. Chem. 1998, 273, 29905–29914. [Google Scholar] [CrossRef] [PubMed]
- Niemann, H.; Geyer, R.; Klenk, H.D.; Linder, D.; Stirm, S.; Wirth, M. The carbohydrates of mouse hepatitis virus (MHV) a59: Structures of the O-glycosidically linked oligosaccharides of glycoprotein e1. EMBO. J. 1984, 3, 665–670. [Google Scholar] [PubMed]
- Oostra, M.; de Haan, C.A.; de Groot, R.J.; Rottier, P.J. Glycosylation of the severe acute respiratory syndrome coronavirus triple-spanning membrane proteins 3a and M. J. Virol. 2006, 80, 2326–2336. [Google Scholar] [CrossRef] [PubMed]
- Klumperman, J.; Locker, J.K.; Meijer, A.; Horzinek, M.C.; Geuze, H.J.; Rottier, P.J. Coronavirus M proteins accumulate in the golgi complex beyond the site of virion budding. J. Virol. 1994, 68, 6523–6534. [Google Scholar] [PubMed]
- Rottier, P.J.; Rose, J.K. Coronavirus e1 glycoprotein expressed from cloned cdna localizes in the golgi region. J. Virol. 1987, 61, 2042–2045. [Google Scholar] [PubMed]
- Nal, B.; Chan, C.; Kien, F.; Siu, L.; Tse, J.; Chu, K.; Kam, J.; Staropoli, I.; Crescenzo-Chaigne, B.; Escriou, N.; et al. Differential maturation and subcellular localization of severe acute respiratory syndrome coronavirus surface proteins S, M and E. J. Gen. Virol. 2005, 86, 1423–1434. [Google Scholar] [CrossRef] [PubMed]
- Mayer, T.; Tamura, T.; Falk, M.; Niemann, H. Membrane integration and intracellular transport of the coronavirus glycoprotein e1, a class III membrane glycoprotein. J. Biol. Chem. 1988, 263, 14956–14963. [Google Scholar] [PubMed]
- Machamer, C.E.; Mentone, S.A.; Rose, J.K.; Farquhar, M.G. The e1 glycoprotein of an avian coronavirus is targeted to the cis golgi complex. Proc. Natl. Acad. Sci. USA 1990, 87, 6944–6948. [Google Scholar] [CrossRef] [PubMed]
- Locker, J.K.; Griffiths, G.; Horzinek, M.C.; Rottier, P.J. O-Glycosylation of the coronavirus M protein. Differential localization of sialyltransferases in n- and o-linked glycosylation. J. Biol. Chem. 1992, 267, 14094–14101. [Google Scholar] [PubMed]
- Laviada, M.D.; Videgain, S.P.; Moreno, L.; Alonso, F.; Enjuanes, L.; Escribano, J.M. Expression of swine transmissible gastroenteritis virus envelope antigens on the surface of infected cells: Epitopes externally exposed. Virus. Res. 1990, 16, 247–254. [Google Scholar] [CrossRef] [PubMed]
- To, L.T.; Bernard, S.; Lantier, I. Fixed-cell immunoperoxidase technique for the study of surface antigens induced by the coronavirus of transmissible gastroenteritis (tgev). Vet. Microbiol. 1991, 29, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Jacobse-Geels, H.E.; Horzinek, M.C. Expression of feline infectious peritonitis coronavirus antigens on the surface of feline macrophage-like cells. J. Gen. Virol. 1983, 64, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Voss, D.; Kern, A.; Traggiai, E.; Eickmann, M.; Stadler, K.; Lanzavecchia, A.; Becker, S. Characterization of severe acute respiratory syndrome coronavirus membrane protein. FEBS. Lett. 2006, 580, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Machamer, C.E.; Grim, M.G.; Esquela, A.; Chung, S.W.; Rolls, M.; Ryan, K.; Swift, A.M. Retention of a cis golgi protein requires polar residues on one face of a predicted alpha-helix in the transmembrane domain. Mol. Biol. Cell 1993, 4, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.; Patel, S.; Riddle, P. Lysosomal sorting mutants of coronavirus e1 protein, a golgi membrane protein. J. Cell. Sci. 1990, 95, 191–197. [Google Scholar] [PubMed]
- Locker, J.K.; Klumperman, J.; Oorschot, V.; Horzinek, M.C.; Geuze, H.J.; Rottier, P.J. The cytoplasmic tail of mouse hepatitis virus m protein is essential but not sufficient for its retention in the golgi complex. J. Biol. Chem. 1994, 269, 28263–28269. [Google Scholar] [PubMed]
- Armstrong, J.; Patel, S. The golgi sorting domain of coronavirus e1 protein. J. Cell. Sci. 1991, 98, 567–575. [Google Scholar] [PubMed]
- Weisz, O.A.; Swift, A.M.; Machamer, C.E. Oligomerization of a membrane protein correlates with its retention in the golgi complex. J. Cell Biol. 1993, 122, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Locker, J.K.; Opstelten, D.J.; Ericsson, M.; Horzinek, M.C.; Rottier, P.J. Oligomerization of a trans-golgi/trans-golgi network retained protein occurs in the golgi complex and may be part of its retention. J. Biol. Chem. 1995, 270, 8815–8821. [Google Scholar] [CrossRef] [PubMed]
- Arndt, A.L.; Larson, B.J.; Hogue, B.G. A conserved domain in the coronavirus membrane protein tail is important for virus assembly. J. Virol. 2010, 84, 11418–11428. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.T.; Chang, C.H.; Wang, S.M.; Huang, K.J.; Wang, C.T. Identifying SARS-cov membrane protein amino acid residues linked to virus-like particle assembly. PLoS ONE 2013, 8, e64013. [Google Scholar] [CrossRef] [PubMed]
- Ruch, T.R.; Machamer, C.E. The coronavirus E protein: Assembly and beyond. Viruses 2012, 4, 363–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boscarino, J.A.; Logan, H.L.; Lacny, J.J.; Gallagher, T.M. Envelope protein palmitoylations are crucial for murine coronavirus assembly. J. Virol. 2008, 82, 2989–2999. [Google Scholar] [CrossRef] [PubMed]
- Corse, E.; Machamer, C.E. The cytoplasmic tails of infectious bronchitis virus E and M proteins mediate their interaction. Virology 2003, 312, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.P.; Liu, D.X. The missing link in coronavirus assembly. Retention of the avian coronavirus infectious bronchitis virus envelope protein in the pre-golgi compartments and physical interaction between the envelope and membrane proteins. J. Biol. Chem. 2001, 276, 17515–17523. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Ye, F.; Sun, T.; Yue, L.; Peng, S.; Chen, J.; Li, G.; Du, Y.; Xie, Y.; Yang, Y.; et al. In vitro biochemical and thermodynamic characterization of nucleocapsid protein of SARS. Biophys. Chem. 2004, 112, 15–25. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Dobie, F.; Ballantine, M.; Leeson, A.; Li, Y.; Bastien, N.; Cutts, T.; Andonov, A.; Cao, J.; Booth, T.F.; et al. Analysis of multimerization of the SARS coronavirus nucleocapsid protein. Biochem. Biophys. Res. Commun. 2004, 316, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Chen, J.; Chen, K.; Shen, X.; Jiang, H. Carboxyl terminus of severe acute respiratory syndrome coronavirus nucleocapsid protein: Self-association analysis and nucleic acid binding characterization. Biochemistry 2006, 45, 11827–11835. [Google Scholar] [CrossRef] [PubMed]
- Robbins, S.G.; Frana, M.F.; McGowan, J.J.; Boyle, J.F.; Holmes, K.V. Rna-binding proteins of coronavirus mhv: Detection of monomeric and multimeric N protein with an rna overlay-protein blot assay. Virology 1986, 150, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Yu, I.M.; Gustafson, C.L.; Diao, J.; Burgner, J.W., 2nd; Li, Z.; Zhang, J.; Chen, J. Recombinant severe acute respiratory syndrome (SARS) coronavirus nucleocapsid protein forms a dimer through its c-terminal domain. J. Biol. Chem. 2005, 280, 23280–23286. [Google Scholar] [CrossRef] [PubMed]
- Surjit, M.; Liu, B.; Kumar, P.; Chow, V.T.; Lal, S.K. The nucleocapsid protein of the SARS coronavirus is capable of self-association through a C-terminal 209 amino acid interaction domain. Biochem. Biophys. Res. Commun. 2004, 317, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Neuman, B.W.; Adair, B.D.; Yoshioka, C.; Quispe, J.D.; Orca, G.; Kuhn, P.; Milligan, R.A.; Yeager, M.; Buchmeier, M.J. Supramolecular architecture of severe acute respiratory syndrome coronavirus revealed by electron cryomicroscopy. J. Virol. 2006, 80, 7918–7928. [Google Scholar] [CrossRef] [PubMed]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Ujike, M.; Morikawa, S.; Tashiro, M.; Taguchi, F. Protease-mediated enhancement of severe acute respiratory syndrome coronavirus infection. Proc. Natl. Acad. Sci. USA 2005, 102, 12543–12547. [Google Scholar] [CrossRef] [PubMed]
- Ujike, M.; Nishikawa, H.; Otaka, A.; Yamamoto, N.; Matsuoka, M.; Kodama, E.; Fujii, N.; Taguchi, F. Heptad repeat-derived peptides block protease-mediated direct entry from the cell surface of severe acute respiratory syndrome coronavirus but not entry via the endosomal pathway. J. Virol. 2008, 82, 588–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, B.J.; de Haan, C.A.; Smits, S.L.; Rottier, P.J. Spike protein assembly into the coronavirion: Exploring the limits of its sequence requirements. Virology 2005, 334, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Bos, E.C.; Heijnen, L.; Luytjes, W.; Spaan, W.J. Mutational analysis of the murine coronavirus spike protein: Effect on cell-to-cell fusion. Virology 1995, 214, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.W.; Sheng, Y.; Gombold, J.L. Coronavirus-induced membrane fusion requires the cysteine-rich domain in the spike protein. Virology 2000, 269, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Petit, C.M.; Chouljenko, V.N.; Iyer, A.; Colgrove, R.; Farzan, M.; Knipe, D.M.; Kousoulas, K.G. Palmitoylation of the cysteine-rich endodomain of the SARS-coronavirus spike glycoprotein is important for spike-mediated cell fusion. Virology 2007, 360, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Vennema, H.; Heijnen, L.; Zijderveld, A.; Horzinek, M.C.; Spaan, W.J. Intracellular transport of recombinant coronavirus spike proteins: Implications for virus assembly. J. Virol. 1990, 64, 339–346. [Google Scholar] [PubMed]
- Schwegmann-Wessels, C.; Al-Falah, M.; Escors, D.; Wang, Z.; Zimmer, G.; Deng, H.; Enjuanes, L.; Naim, H.Y.; Herrler, G. A novel sorting signal for intracellular localization is present in the S protein of a porcine coronavirus but absent from severe acute respiratory syndrome-associated coronavirus. J. Biol. Chem. 2004, 279, 43661–43666. [Google Scholar] [CrossRef] [PubMed]
- McBride, C.E.; Li, J.; Machamer, C.E. The cytoplasmic tail of the severe acute respiratory syndrome coronavirus spike protein contains a novel endoplasmic reticulum retrieval signal that binds copi and promotes interaction with membrane protein. J. Virol. 2007, 81, 2418–2428. [Google Scholar] [CrossRef] [PubMed]
- Winter, C.; Schwegmann-Wessels, C.; Neumann, U.; Herrler, G. The spike protein of infectious bronchitis virus is retained intracellularly by a tyrosine motif. J. Virol. 2008, 82, 2765–2771. [Google Scholar] [CrossRef] [PubMed]
- Bonifacino, J.S.; Dell’Angelica, E.C. Molecular bases for the recognition of tyrosine-based sorting signals. J. Cell Biol. 1999, 145, 923–926. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.T.; McCoy, A.J.; Spate, K.; Miller, S.E.; Evans, P.R.; Honing, S.; Owen, D.J. A structural explanation for the binding of endocytic dileucine motifs by the ap2 complex. Nature 2008, 456, 976–979. [Google Scholar] [CrossRef] [PubMed]
- Shirato, K.; Maejima, M.; Matsuyama, S.; Ujike, M.; Miyazaki, A.; Takeyama, N.; Ikeda, H.; Taguchi, F. Mutation in the cytoplasmic retrieval signal of porcine epidemic diarrhea virus spike (S) protein is responsible for enhanced fusion activity. Virus. Res. 2011, 161, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Teasdale, R.D.; Jackson, M.R. Signal-mediated sorting of membrane proteins between the endoplasmic reticulum and the golgi apparatus. Annu. Rev. Cell Dev. Biol. 1996, 12, 27–54. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.V.; Doller, E.W.; Sturman, L.S. Tunicamycin resistant glycosylation of coronavirus glycoprotein: Demonstration of a novel type of viral glycoprotein. Virology 1981, 115, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Rottier, P.J.; Horzinek, M.C.; van der Zeijst, B.A. Viral protein synthesis in mouse hepatitis virus strain a59-infected cells: Effect of tunicamycin. J. Virol. 1981, 40, 350–357. [Google Scholar] [PubMed]
- De Haan, C.A.; Smeets, M.; Vernooij, F.; Vennema, H.; Rottier, P.J. Mapping of the coronavirus membrane protein domains involved in interaction with the spike protein. J. Virol. 1999, 73, 7441–7452. [Google Scholar] [PubMed]
- McBride, C.E.; Machamer, C.E. A single tyrosine in the severe acute respiratory syndrome coronavirus membrane protein cytoplasmic tail is important for efficient interaction with spike protein. J. Virol. 2010, 84, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.J.; Evans, P.R. A structural explanation for the recognition of tyrosine-based endocytotic signals. Science 1998, 282, 1327–1332. [Google Scholar] [CrossRef] [PubMed]
- Godeke, G.J.; de Haan, C.A.; Rossen, J.W.; Vennema, H.; Rottier, P.J. Assembly of spikes into coronavirus particles is mediated by the carboxy-terminal domain of the spike protein. J. Virol. 2000, 74, 1566–1571. [Google Scholar] [CrossRef] [PubMed]
- Ujike, M.; Huang, C.; Shirato, K.; Matsuyama, S.; Makino, S.; Taguchi, F. Two palmitylated cysteine residues of the severe acute respiratory syndrome coronavirus spike (S) protein are critical for s incorporation into virus-like particles, but not for m-s co-localization. J. Gen. Virol. 2012, 93, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Thorp, E.B.; Boscarino, J.A.; Logan, H.L.; Goletz, J.T.; Gallagher, T.M. Palmitoylations on murine coronavirus spike proteins are essential for virion assembly and infectivity. J. Virol. 2006, 80, 1280–1289. [Google Scholar] [CrossRef] [PubMed]
- Shulla, A.; Gallagher, T. Role of spike protein endodomains in regulating coronavirus entry. J. Biol. Chem. 2009, 284, 32725–32734. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lv, J.; Wang, Y.; Gao, S.; Yao, Q.; Qu, D.; Ye, R. Replication of murine coronavirus requires multiple cysteines in the endodomain of spike protein. Virology 2012, 427, 98–106. [Google Scholar] [CrossRef] [PubMed]
- McBride, C.E.; Machamer, C.E. Palmitoylation of SARS-cov s protein is necessary for partitioning into detergent-resistant membranes and cell-cell fusion but not interaction with m protein. Virology 2010, 405, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Gelhaus, S.; Thaa, B.; Eschke, K.; Veit, M.; Schwegmann-Wessels, C. Palmitoylation of the alphacoronavirus tgev spike protein S is essential for incorporation into virus-like particles but dispensable for s-m interaction. Virology 2014, 464–465, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Youn, S.; Collisson, E.W.; Machamer, C.E. Contribution of trafficking signals in the cytoplasmic tail of the infectious bronchitis virus spike protein to virus infection. J. Virol. 2005, 79, 13209–13217. [Google Scholar] [CrossRef] [PubMed]
- Shirato, K.; Maejima, M.; Hirai, A.; Ami, Y.; Takeyama, N.; Tsuchiya, K.; Kusanagi, K.; Nunoya, T.; Taguchi, F. Enhanced cell fusion activity in porcine epidemic diarrhea virus adapted to suckling mice. Arch. Virol. 2010, 155, 1989–1995. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ujike, M.; Taguchi, F. Incorporation of Spike and Membrane Glycoproteins into Coronavirus Virions. Viruses 2015, 7, 1700-1725. https://doi.org/10.3390/v7041700
Ujike M, Taguchi F. Incorporation of Spike and Membrane Glycoproteins into Coronavirus Virions. Viruses. 2015; 7(4):1700-1725. https://doi.org/10.3390/v7041700
Chicago/Turabian StyleUjike, Makoto, and Fumihiro Taguchi. 2015. "Incorporation of Spike and Membrane Glycoproteins into Coronavirus Virions" Viruses 7, no. 4: 1700-1725. https://doi.org/10.3390/v7041700
APA StyleUjike, M., & Taguchi, F. (2015). Incorporation of Spike and Membrane Glycoproteins into Coronavirus Virions. Viruses, 7(4), 1700-1725. https://doi.org/10.3390/v7041700