
In Ovo Delivery of CpG DNA Reduces Avian Infectious Laryngotracheitis Virus Induced Mortality and Morbidity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
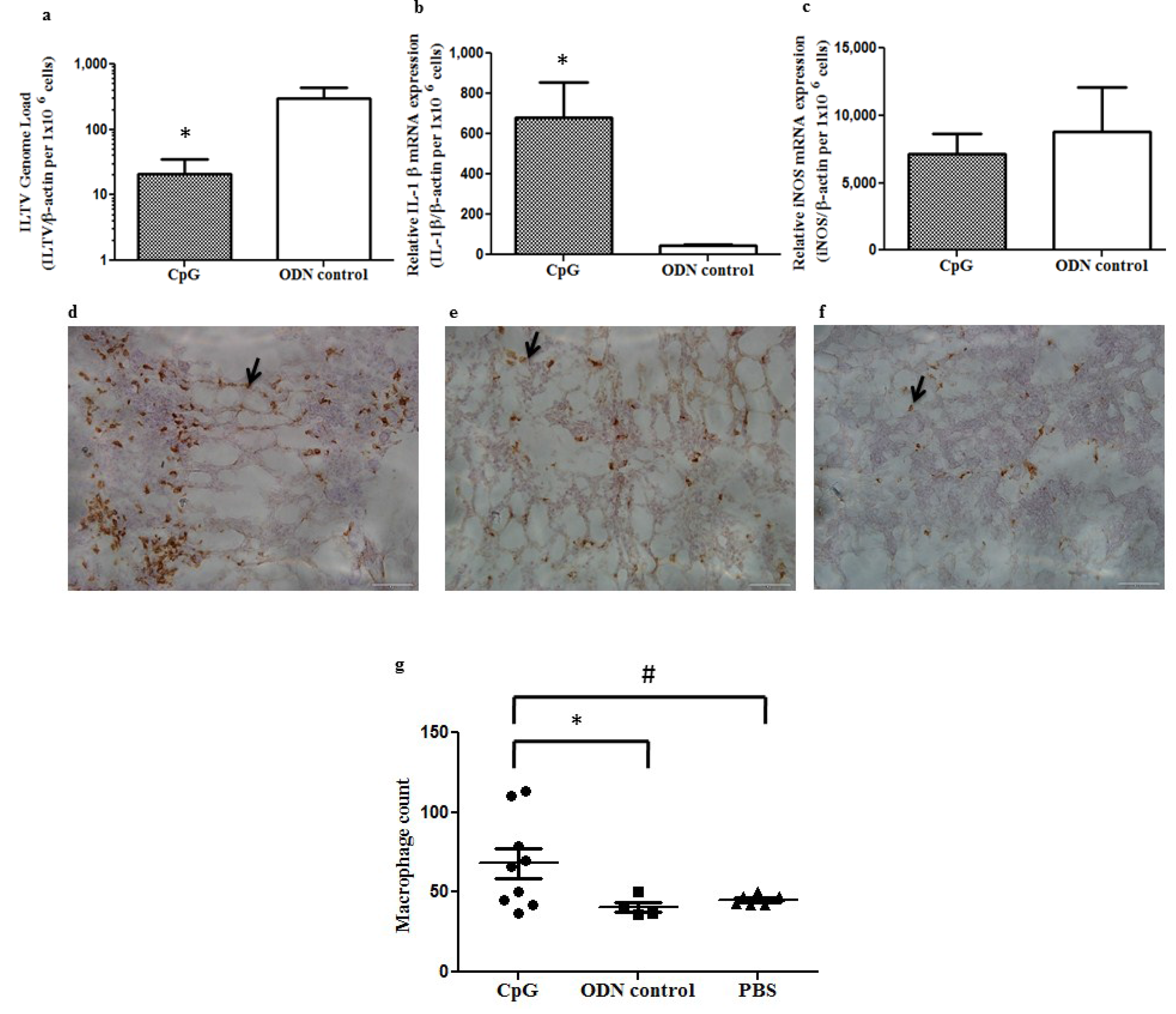
2.1. In Ovo Delivery of CpG DNA Reduces Pre-Hatch ILTV Replication in Lung of Embryos Correlating with Expansion of Macrophage Populations and Increased mRNA Expression of IL-1β in Lungs

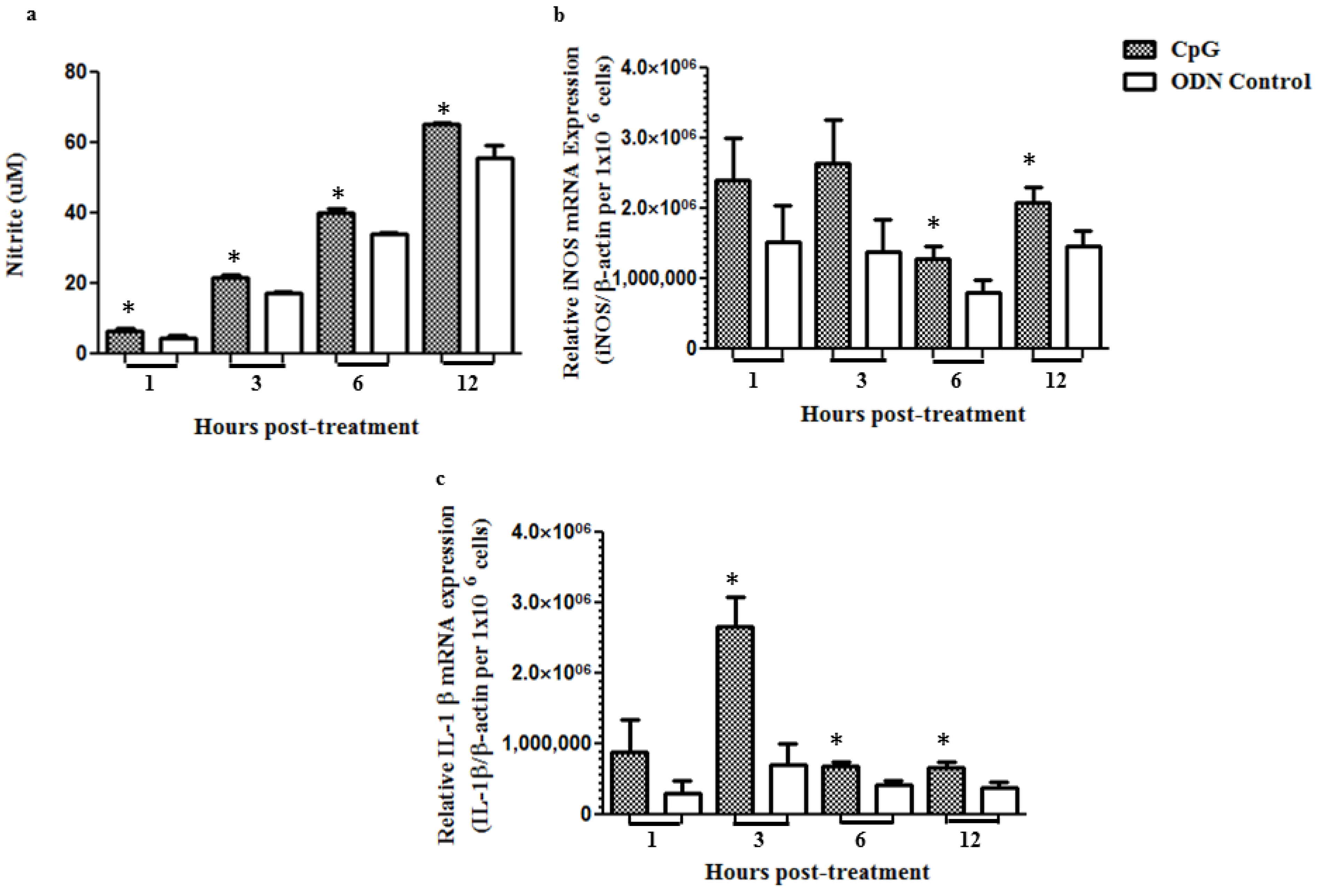
2.2. Stimulation of Macrophages in Vitro with CpG DNA up Regulates mRNA Expression of Pro-Inflammatory Mediators and Increases NO Production


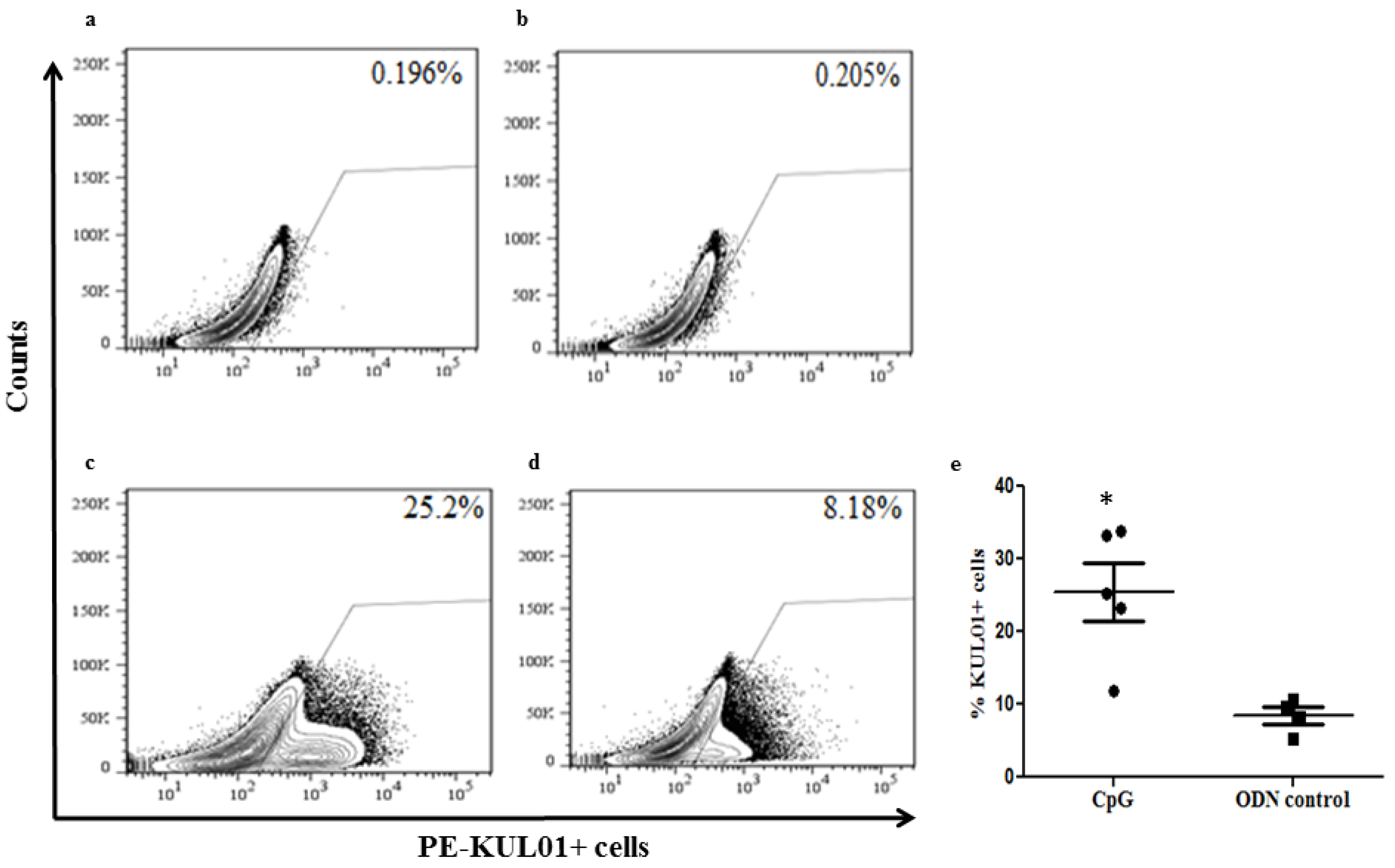
2.3. In Ovo Delivery of CpG DNA Expands Macrophage Numbers in Lungs Post-Hatch

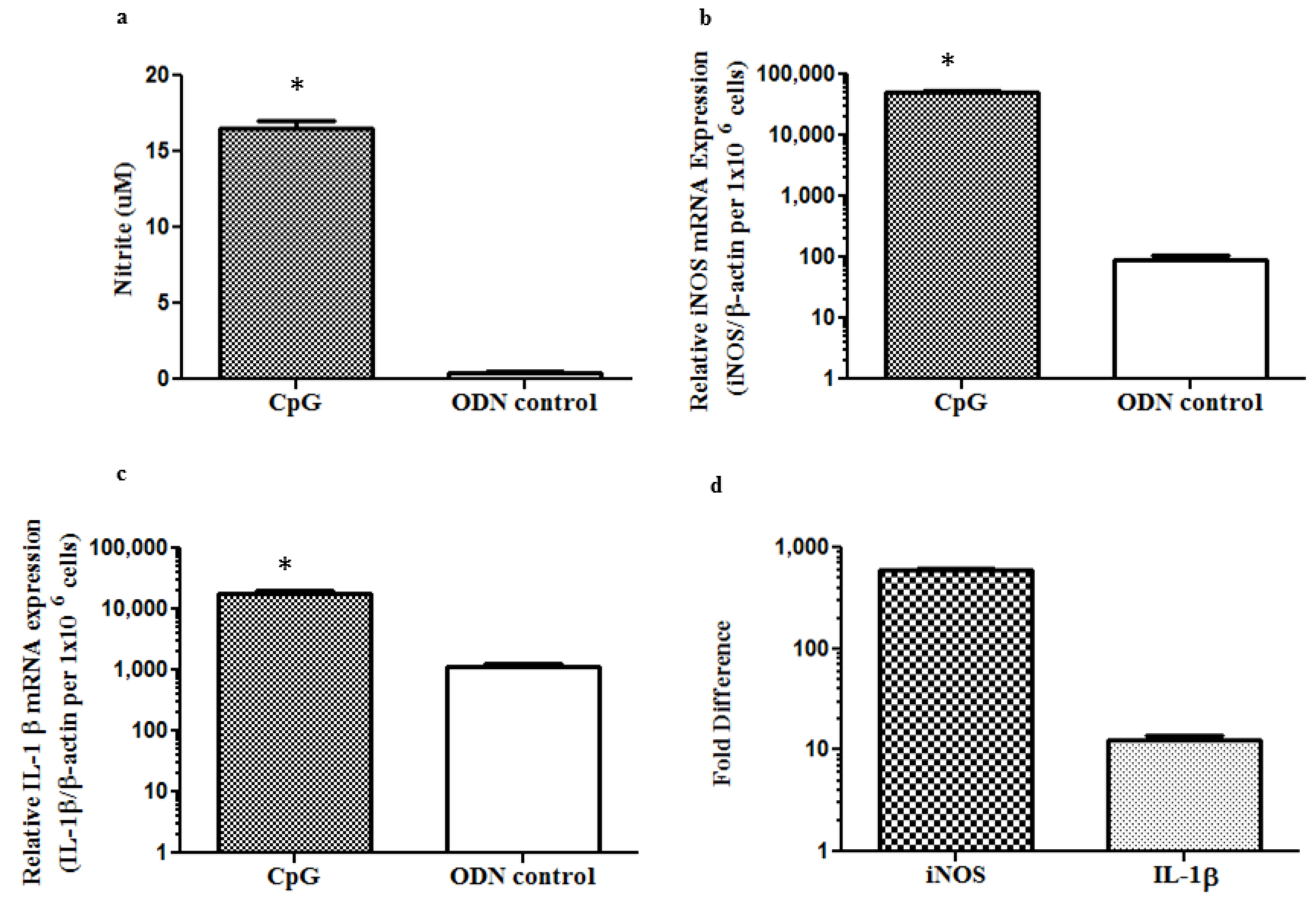
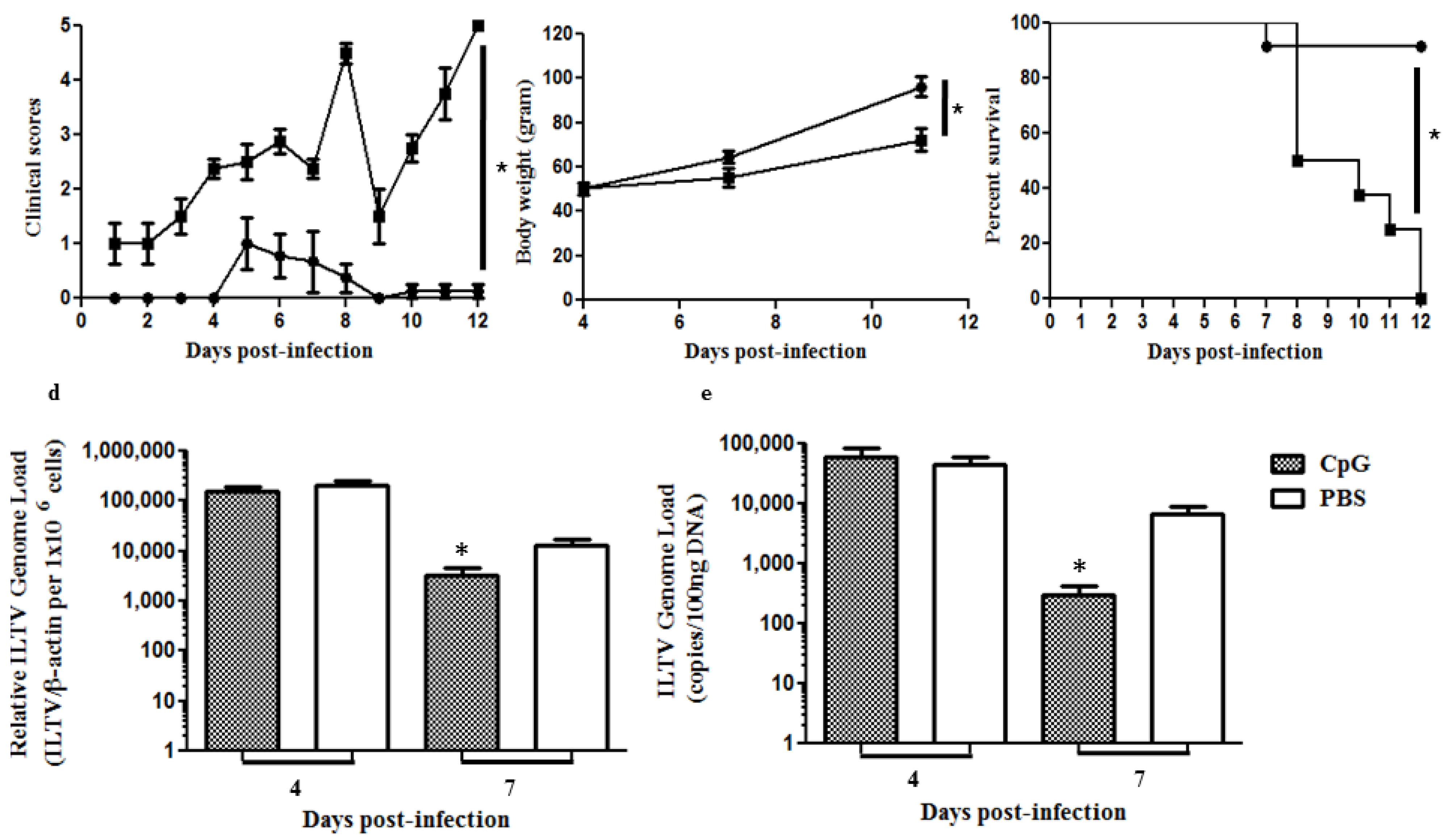
2.4. In Ovo Delivery of CpG DNA Induces Protection against ILTV Caused Morbidity and Mortality Reducing Viral Replication in the Respiratory Tract of Chickens

2.5. In Ovo Delivery of CpG DNA Does Not Affect the Hatchability of Eggs
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cells, Virus and TLR-Ligand
4.3. Cell Culture
4.4. Experimental Design
4.4.2. Stimulation of Macrophages in Vitro with CpG DNA to Determine the Expression of Pro-Inflammatory Mediators
4.4.3. In Ovo Delivered CpG mediated Expansion of Macrophage Populations in Lungs Post-Hatch
4.4.4. In Ovo Delivery of CpG DNA Mediated Protection against ILTV Caused Morbidity and Mortality in Chickens
4.4.5. Determination of Safety of in Ovo Delivery of CpG DNA
4.5. Primers
4.6. DNA and RNA Extraction
4.7. Reverse Transcription
4.8. Assay for NO Production
4.9. Flow Cytometry Technique
4.10. Immunohistochemistry Technique
4.11. Real-Time PCR Assay
4.12. Data Analyses
Acknowledgments
Author Contributions
Conflict of Interest
References
- McGeoch, D.J.; Dolan, A.; Ralph, A.C. Toward a comprehensive phylogeny for mammalian and avian herpesviruses. J. Virol. 2000, 74, 10401–10406. [Google Scholar] [CrossRef]
- McGeoch, D.J.; Rixon, F.J.; Davison, A.J. Topics in herpesvirus genomics and evolution. Virus Res. 2006, 117, 90–104. [Google Scholar] [CrossRef]
- Fahey, K.J.; Bagust, T.J.; York, J.J. Laryngotracheitis herpesvirus infection in the chicken: The role of humoral antibody in immunity to a graded challenge infection. Avian Pathol. 1983, 12, 505–514. [Google Scholar] [CrossRef]
- Crawshaw, G.J.; Boycott, B.R. Infectious laryngotracheitis in peafowl and pheasants. Avian Dis. 1982, 26, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Bagust, T.J.; Jones, R.C.; Guy, J.S. Avian infectious laryngotracheitis. Rev. Sci. Technol. 2000, 19, 483–492. [Google Scholar]
- Lee, Y.J.; Park, S.S.; Kim, S.Y.; Lee, J.Y.; Koo, H.K.; Yoon, H.I. A case of systemic polyarteritis nodosa involving bronchial artery. Sarcoidosis Vasc. Diffuse Lung Dis. 2010, 27, 164–168. [Google Scholar] [PubMed]
- Fahey, K.J.; Bagust, T.J.; York, J.J. Laryngotracheitis Herpesvirus-Infection in the Chicken—The Role of Humoral Antibody in Immunity to a Graded Challenge Infection. Avian Pathol. 1983, 12, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Okamura, H.; Taneno, A.; Yamada, S.; Takahashi, E. The Role of Cell-Mediated-Immunity in Chickens Inoculated with the Cell-Associated Vaccine of Attenuated Infectious Laryngotracheitis Virus. J. Vet. Med. Sci. 1994, 56, 1051–1055. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.C.; Lee, Y.L.; Shien, J.H.; Shieh, H.K. Rapid differentiation of vaccine strains and field isolates of infectious laryngotracheitis virus by restriction fragment length polymorphism of PCR products. J. Virol. Methods 1997, 66, 179–86. [Google Scholar] [CrossRef] [PubMed]
- Oldoni, I.; Rodriguez-Avila, A.; Riblet, S.; Garcia, M. Characterization of infectious laryngotracheitis virus (ILTV) isolates from commercial poultry by polymerase chain reaction and restriction fragment length polymorphism (PCR-RFLP). Avian Dis. 2008, 52, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Devlin, J.M.; Markham, J.F.; Noormohammadi, A.H.; Browning, G.F.; Ficorilli, N.P.; Hartley, C.A.; Markham, P.F. Phylogenetic and molecular epidemiological studies reveal evidence of multiple past recombination events between infectious laryngotracheitis viruses. PLoS ONE 2013, 8, e55121. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Markham, P.F.; Coppo, M.J.; Legione, A.R.; Markham, J.F.; Noormohammadi, A.H.; Browning, G.F.; Ficorilli, N.; Hartley, C.A.; Devlin, J.M. Attenuated vaccines can recombine to form virulent field viruses. Science 2012, 337, 188. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2001, 2, 675–680. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors—Redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.S.; Li, G. Nitric oxide production by macrophages stimulated with coccidia sporozoites, lipopolysaccharide, or interferon-γ, and its dynamic changes in SC and TK strains of chickens infected with Eimeria tenella. Avian Dis. 2004, 48, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Lynn, D.J.; Lloyd, A.T.; O’Farrelly, C. In silico identification of components of the Toll-like receptor (TLR) signaling pathway in clustered chicken expressed sequence tags (ESTs). Vet. Immunol. Immunopathol. 2003, 93, 177–184. [Google Scholar] [CrossRef]
- Philbin, V.J.; Iqbal, M.; Boyd, Y.; Goodchild, M.J.; Beal, R.K.; Bumstead, N.; Young, J.; Smith, A.L. Identification and characterization of a functional, alternatively spliced Toll-like receptor 7 (TLR7) and genomic disruption of TLR8 in chickens. Immunology 2005, 114, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.C.; Peroval, M.Y.; Hammond, J.A.; Prickett, M.D.; Young, J.R.; Smith, A.L. TLR15 is unique to avian and reptilian lineages and recognizes a yeast-derived agonist. J. Immunol. 2012, 189, 4930–4938. [Google Scholar] [CrossRef] [PubMed]
- Brownlie, R.; Zhu, J.; Allan, B.; Mutwiri, G.K.; Babiuk, L.A.; Potter, A.; Griebel, P. Chicken TLR21 acts as a functional homologue to mammalian TLR9 in the recognition of CpG oligodeoxynucleotides. Mol. Immunol. 2009, 46, 3163–3170. [Google Scholar] [CrossRef] [PubMed]
- Cormican, P.; Lloyd, A.T.; Downing, T.; Connell, S.J.; Bradley, D.; O’Farrelly, C. The avian Toll-Like receptor pathway—Subtle differences amidst general conformity. Dev. Comp. Immunol. 2009, 33, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Roach, J.C.; Glusman, G.; Rowen, L.; Kaur, A.; Purcell, M.K.; Smith, K.D.; Hood, L.E.; Aderem, A. The evolution of vertebrate Toll-like receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 9577–9582. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, H.; Schneider, K.; Ohnemus, A.; Lavric, M.; Kothlow, S.; Bauer, S.; Kaspers, B.; Staeheli, P. Chicken toll-like receptor 3 recognizes its cognate ligand when ectopically expressed in human cells. J. Interferon Cytokine Res. 2007, 27, 97–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temperley, N.D.; Berlin, S.; Paton, I.R.; Griffin, D.K.; Burt, D.W. Evolution of the chicken Toll-like receptor gene family: A story of gene gain and gene loss. BMC Genomics 2008, 9, e62. [Google Scholar] [CrossRef]
- Nishiya, T.; DeFranco, A.L. Ligand-regulated chimeric receptor approach reveals distinctive subcellular localization and signaling properties of the Toll-like receptors. J. Biol. Chem. 2004, 279, 19008–19017. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740–745. [Google Scholar]
- Krieg, A.M. Therapeutic potential of Toll-like receptor 9 activation. Nat. Rev. Drug. Discov. 2006, 5, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Verthelyi, D.; Ishii, K.J.; Gursel, M.; Takeshita, F.; Klinman, D.M. Human peripheral blood cells differentially recognize and respond to two distinct CPG motifs. J. Immunol. 2001, 166, 2372–2377. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, G.; Weeratna, R.D.; Ballas, Z.K.; Payette, P.; Blackwell, S.; Suparto, I.; Rasmussen, W.L.; Waldschmidt, M.; Sajuthi, D.; Purcell, R.H.; et al. Delineation of a CpG phosphorothioate oligodeoxynucleotide for activating primate immune responses in vitro and in vivo. J. Immunol. 2000, 164, 1617–1624. [Google Scholar]
- Gursel, M.; Verthelyi, D.; Gursel, I.; Ishii, K.J.; Klinman, D.M. Differential and competitive activation of human immune cells by distinct classes of CpG oligodeoxynucleotide. J. Leukoc. Biol. 2002, 71, 813–820. [Google Scholar] [PubMed]
- Marshall, J.D.; Fearon, K.; Abbate, C.; Subramanian, S.; Yee, P.; Gregorio, J.; Coffman, R.L.; van Nest, G. Identification of a novel CpG DNA class and motif that optimally stimulate B cell and plasmacytoid dendritic cell functions. J. Leukoc. Biol. 2003, 73, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, S.; Egeter, O.; Hausmann, S.; Lipford, G.B.; Rocken, M.; Wagner, H.; Heeg, K. CpG oligodeoxynucleotides trigger protective and curative Th1 responses in lethal murine leishmaniasis. J. Immunol. 1998, 160, 3627–3630. [Google Scholar] [PubMed]
- Cheng, C.; Bettahi, I.; Cruz-Fisher, M.I.; Pal, S.; Jain, P.; Jia, Z.; Holmgren, J.; Harandi, A.M.; de la Maza, L.M. Induction of protective immunity by vaccination against Chlamydia trachomatis using the major outer membrane protein adjuvanted with CpG oligodeoxynucleotide coupled to the nontoxic B subunit of cholera toxin. Vaccine 2009, 27, 6239–6246. [Google Scholar] [CrossRef] [PubMed]
- Nystrom-Asklin, J.; Adamsson, J.; Harandi, A.M. The adjuvant effect of CpG oligodeoxynucleotide linked to the non-toxic B subunit of cholera toxin for induction of immunity against H. pylori in mice. Scand. J. Immunol. 2008, 67, 431–440. [Google Scholar] [CrossRef]
- Oxenius, A.; Martinic, M.M.; Hengartner, H.; Klenerman, P. CpG-containing oligonucleotides are efficient adjuvants for induction of protective antiviral immune responses with T-cell peptide vaccines. J. Virol. 1999, 73, 4120–4126. [Google Scholar] [PubMed]
- Davis, H.L.; Weeratna, R.; Waldschmidt, T.J.; Tygrett, L.; Schorr, J.; Krieg, A.M. CpG DNA is a potent enhancer of specific immunity in mice immunized with recombinant hepatitis B surface antigen. J. Immunol. 1998, 160, 870–876. [Google Scholar] [PubMed]
- Belyakov, I.M.; Isakov, D.; Zhu, Q.; Dzutsev, A.; Klinman, D.; Berzofsky, J.A. Enhancement of CD8+ T cell immunity in the lung by CpG oligodeoxynucleotides increases protective efficacy of a modified vaccinia Ankara vaccine against lethal poxvirus infection even in a CD4-deficient host. J. Immunol. 2006, 177, 6336–6343. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, M.M.; DeVeer, M.J.; Edling, A.; Oates, R.K.; Simons, B.; Lindner, D.; Williams, B.R. Synergistic activation of innate immunity by double-stranded RNA and CpG DNA promotes enhanced antitumor activity. Cancer Res. 2004, 64, 5850–5860. [Google Scholar] [CrossRef] [PubMed]
- Weiner, G.J.; Liu, H.M.; Wooldridge, J.E.; Dahle, C.E.; Krieg, A.M. Immunostimulatory oligodeoxynucleotides containing the CpG motif are effective as immune adjuvants in tumor antigen immunization. Proc. Natl. Acad. Sci. USA 1997, 94, 10833–10837. [Google Scholar] [CrossRef] [PubMed]
- Tulic, M.K.; Fiset, P.O.; Christodoulopoulos, P.; Vaillancourt, P.; Desrosiers, M.; Lavigne, F.; Eiden, J.; Hamid, Q. Amb a 1-immunostimulatory oligodeoxynucleotide conjugate immunotherapy decreases the nasal inflammatory response. J. Allergy Clin. Immunol. 2004, 113, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.M. Development of TLR9 agonists for cancer therapy. J. Clin. Invest. 2007, 117, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, A.; Laigle-Donadey, F.; Zohar, S.; Capelle, L.; Behin, A.; Tibi, A.; Martin-Duverneuil, N.; Sanson, M.; Lacomblez, L.; Taillibert, S.; et al. Phase 1 trial of a CpG oligodeoxynucleotide for patients with recurrent glioblastoma. Neuro. Oncol. 2006, 8, 60–66. [Google Scholar]
- Cooper, C.L.; Davis, H.L.; Morris, M.L.; Efler, S.M.; Adhami, M.A.; Krieg, A.M.; Cameron, D.W.; Heathcote, J. CPG 7909, an immunostimulatory TLR9 agonist oligodeoxynucleotide, as adjuvant to Engerix-B HBV vaccine in healthy adults: A double-blind phase I/II study. J. Clin. Immunol. 2004, 24, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.L.; Davis, H.L.; Angel, J.B.; Morris, M.L.; Elfer, S.M.; Seguin, I.; Krieg, A.M.; Cameron, D.W. CPG 7909 adjuvant improves hepatitis B virus vaccine seroprotection in antiretroviral-treated HIV-infected adults. AIDS 2005, 19, 1473–1479. [Google Scholar] [CrossRef] [PubMed]
- Gomis, S.; Babiuk, L.; Godson, D.L.; Allan, B.; Thrush, T.; Townsend, H.; Willson, P.; Waters, E.; Hecker, R.; Potter, A. Protection of chickens against Escherichia coli infections by DNA containing CpG motifs. Infect. Immun. 2003, 71, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Gomis, S.; Babiuk, L.; Allan, B.; Willson, P.; Waters, E.; Ambrose, N.; Hecker, R.; Potter, A. Protection of neonatal chicks against a lethal challenge of Escherichia coli using DNA containing cytosine-phosphodiester-guanine motifs. Avian Dis. 2004, 48, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, A.; Allan, B.; Mutwiri, G.; Van Kessel, A.; Willson, P.; Babiuk, L.; Potter, A.; Gomis, S. Protection of neonatal broiler chicks against Salmonella Typhimurium septicemia by DNA containing CpG motifs. Avian Dis. 2008, 52, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, K.M.; He, H.; Swaggerty, C.L.; McReynolds, J.L.; Genovese, K.J.; Duke, S.E.; Nerren, J.R.; Kogut, M.H. In ovo treatment with CpG oligodeoxynucleotides decreases colonization of Salmonella enteriditis in broiler chickens. Vet. Immunol. Immunopathol. 2009, 127, 371–375. [Google Scholar] [CrossRef]
- St Paul, M.; Mallick, A.I.; Read, L.R.; Villanueva, A.I.; Parvizi, P.; Abdul-Careem, M.F.; Nagy, E.; Sharif, S. Prophylactic treatment with Toll-like receptor ligands enhances host immunity to avian influenza virus in chickens. Vaccine 2012, 30, 4524–4531. [Google Scholar] [CrossRef] [PubMed]
- Barjesteh, N.; Behboudi, S.; Brisbin, J.T.; Villanueva, A.I.; Nagy, E.; Sharif, S. TLR ligands induce antiviral responses in chicken macrophages. PLoS ONE 2014, 9, e105713. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Genovese, K.J.; Swaggerty, C.L.; Nisbet, D.J.; Kogut, M.H. In vivo priming heterophil innate immune functions and increasing resistance to Salmonella enteritidis infection in neonatal chickens by immune stimulatory CpG oligodeoxynucleotides. Vet. Immunol. Immunopathol. 2007, 117, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Potter, A.; Tikoo, S.; Gerdts, V.; Lai, K.; Babiuk, L.A.; Mutwiri, G. CpG oligodeoxynucleotides activate innate immune response that suppresses infectious bronchitis virus replication in chicken embryos. Avian Dis. 2009, 53, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Dar, A.; Tikoo, S.; Potter, A.; Babiuk, L.A.; Townsend, H.; Gerdts, V.; Mutwiri, G. CpG-ODNs induced changes in cytokine/chemokines genes expression associated with suppression of infectious bronchitis virus replication in chicken lungs. Vet. Immunol. Immunopathol. 2014, 160, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Song, J.J.; Wooming, A.; Li, X.Y.; Zhou, H.J.; Bottje, W.G.; Kong, B.W. Transcriptional profiling of host gene expression in chicken embryo lung cells infected with laryngotracheitis virus. BMC Genomics 2010, 11, e445. [Google Scholar] [CrossRef]
- Purcell, D.A. Ultrastructural Changes Produced by Infectious Laryngotracheitis Virus in Tracheal Epithelium of Fowl. Res. Vet. Sci. 1971, 12, 455–458. [Google Scholar] [PubMed]
- Keestra, A.M.; de Zoete, M.R.; Bouwman, L.I.; van Putten, J.P. Chicken TLR21 is an innate CpG DNA receptor distinct from mammalian TLR9. J. Immunol. 2010, 185, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Cheng, A.; Wang, M. Innate sensing of viruses by pattern recognition receptors in birds. Vet.Res. 2013, 44, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Simon, A.; van de Veerdonk, F.; Kullberg, B.J.; Van der Meer, J.W.; Joosten, L.A. IL-1β processing in host defense: Beyond the inflammasomes. PLoS Pathog. 2010, 6, e1000661. [Google Scholar] [CrossRef] [PubMed]
- Anders, H.J.; Banas, B.; Linde, Y.; Weller, L.; Cohen, C.D.; Kretzler, M.; Martin, S.; Vielhauer, V.; Schlondorff, D.; Grone, H.J. Bacterial CpG-DNA aggravates immune complex glomerulonephritis: Role of TLR9-mediated expression of chemokines and chemokine receptors. J. Am. Soc. Nephrol. 2003, 14, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Mathes, A.L.; Rice, L.; Affandi, A.J.; DiMarzio, M.; Rifkin, I.R.; Stifano, G.; Christmann, R.B.; Lafyatis, R. CpGB DNA activates dermal macrophages and specifically recruits inflammatory monocytes into the skin. Exp. Dermatol. 2015, 24, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Ghosh, A.K.; Singh, S.; Saha, B.; Ganguly, A.; Das, P. Unmethylated CpG motifs in the L. donovani DNA regulate TLR9-dependent delay of programmed cell death in macrophages. J. Leukoc. Biol. 2015, 97, 363–378. [Google Scholar]
- Hashimoto, Y.; Moki, T.; Takizawa, T.; Shiratsuchi, A.; Nakanishi, Y. Evidence for phagocytosis of influenza virus-infected, apoptotic cells by neutrophils and macrophages in mice. J. Immunol. 2007, 178, 2448–2457. [Google Scholar] [CrossRef] [PubMed]
- Djeraba, A.; Bernardet, N.; Dambrine, G.; Quere, P. Nitric oxide inhibits Marek’s disease virus replication but is not the single decisive factor in interferon-gamma-mediated viral inhibition. Virology 2000, 277, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Haddadi, S.; Kim, D.S.; Jasmine, H.; van der Meer, F.; Czub, M.; Abdul-Careem, M.F. Induction of Toll-like receptor 4 signaling in avian macrophages inhibits infectious laryngotracheitis virus replication in a nitric oxide dependent way. Vet. Immunol. Immunopathol. 2013, 155, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Schat, K.A.; Xing, Z. Specific and nonspecific immune responses to Marek’s disease virus. Dev. Comp. Immunol. 2000, 24, 201–221. [Google Scholar] [CrossRef] [PubMed]
- Khatri, M.; Palmquist, J.M.; Cha, R.M.; Sharma, J.M. Infection and activation of bursal macrophages by virulent infectious bursal disease virus. Virus Res. 2005, 113, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Ramos, H.J.; Lanteri, M.C.; Blahnik, G.; Negash, A.; Suthar, M.S.; Brassil, M.M.; Sodhi, K.; Treuting, P.M.; Busch, M.P.; Norris, P.J.; et al. IL-1beta signaling promotes CNS-intrinsic immune control of West Nile virus infection. PLoS Pathog. 2012, 8, e1003039. [Google Scholar]
- Cai, H.; Kuang, Z.; Huang, K.; Shi, J.; Zhao, X.; Chu, P.; Huang, C.; Ming, F.; Xia, F.; Yang, J.; et al. CpG oligodeoxynucleotide protect neonatal piglets from challenge with the enterotoxigenic E. coli. Vet. Immunol. Immunopathol. 2014, 161, 66–76. [Google Scholar]
- Krieg, A.M. CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef] [PubMed]
- Calnek, B.W.; Fahey, K.J.; Bagust, T.J. In vitro infection studies with infectious laryngotracheitis virus. Avian Dis. 1986, 30, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.M.; Burmester, B.R. Resistance to Marek's disease at hatching in chickens vaccinated as embryos with the turkey herpesvirus. Avian Dis. 1982, 26, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Wigley, P.; Hulme, S.D.; Bumstead, N.; Barrow, P.A. In vivo and in vitro studies of genetic resistance to systemic salmonellosis in the chicken encoded by the SAL1 locus. Microbes Infect. 2002, 4, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Okamura, M.; Lillehoj, H.S.; Raybourne, R.B.; Babu, U.S.; Heckert, R.A. Cell-mediated immune responses to a killed Salmonella enteritidis vaccine: Lymphocyte proliferation, T-cell changes and interleukin-6 (IL-6), IL-1, IL-2, and IFN-gamma production. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Haddadi, S.; Thapa, S.; Kameka, A.M.; Hui, J.; Czub, M.; Nagy, E.; Muench, G.; Abdul-Careem, M.F. Toll-like receptor 2 ligand, lipoteichoic acid is inhibitory against infectious laryngotracheitis virus infection in vitro and in vivo. Dev. Comp. Immunol. 2015, 48, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Careem, M.F.; Hunter, B.D.; Sarson, A.J.; Mayameei, A.; Zhou, H.; Sharif, S. Marek’s disease virus-induced transient paralysis is associated with cytokine gene expression in the nervous system. Viral Immunol. 2006, 19, 167–176. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thapa, S.; Cader, M.S.A.; Murugananthan, K.; Nagy, E.; Sharif, S.; Czub, M.; Abdul-Careem, M.F. In Ovo Delivery of CpG DNA Reduces Avian Infectious Laryngotracheitis Virus Induced Mortality and Morbidity. Viruses 2015, 7, 1832-1852. https://doi.org/10.3390/v7041832
Thapa S, Cader MSA, Murugananthan K, Nagy E, Sharif S, Czub M, Abdul-Careem MF. In Ovo Delivery of CpG DNA Reduces Avian Infectious Laryngotracheitis Virus Induced Mortality and Morbidity. Viruses. 2015; 7(4):1832-1852. https://doi.org/10.3390/v7041832
Chicago/Turabian StyleThapa, Simrika, Mohamed Sarjoon Abdul Cader, Kalamathy Murugananthan, Eva Nagy, Shayan Sharif, Markus Czub, and Mohamed Faizal Abdul-Careem. 2015. "In Ovo Delivery of CpG DNA Reduces Avian Infectious Laryngotracheitis Virus Induced Mortality and Morbidity" Viruses 7, no. 4: 1832-1852. https://doi.org/10.3390/v7041832
APA StyleThapa, S., Cader, M. S. A., Murugananthan, K., Nagy, E., Sharif, S., Czub, M., & Abdul-Careem, M. F. (2015). In Ovo Delivery of CpG DNA Reduces Avian Infectious Laryngotracheitis Virus Induced Mortality and Morbidity. Viruses, 7(4), 1832-1852. https://doi.org/10.3390/v7041832