
A Current Overview of the Papaya meleira virus, an Unusual Plant Virus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
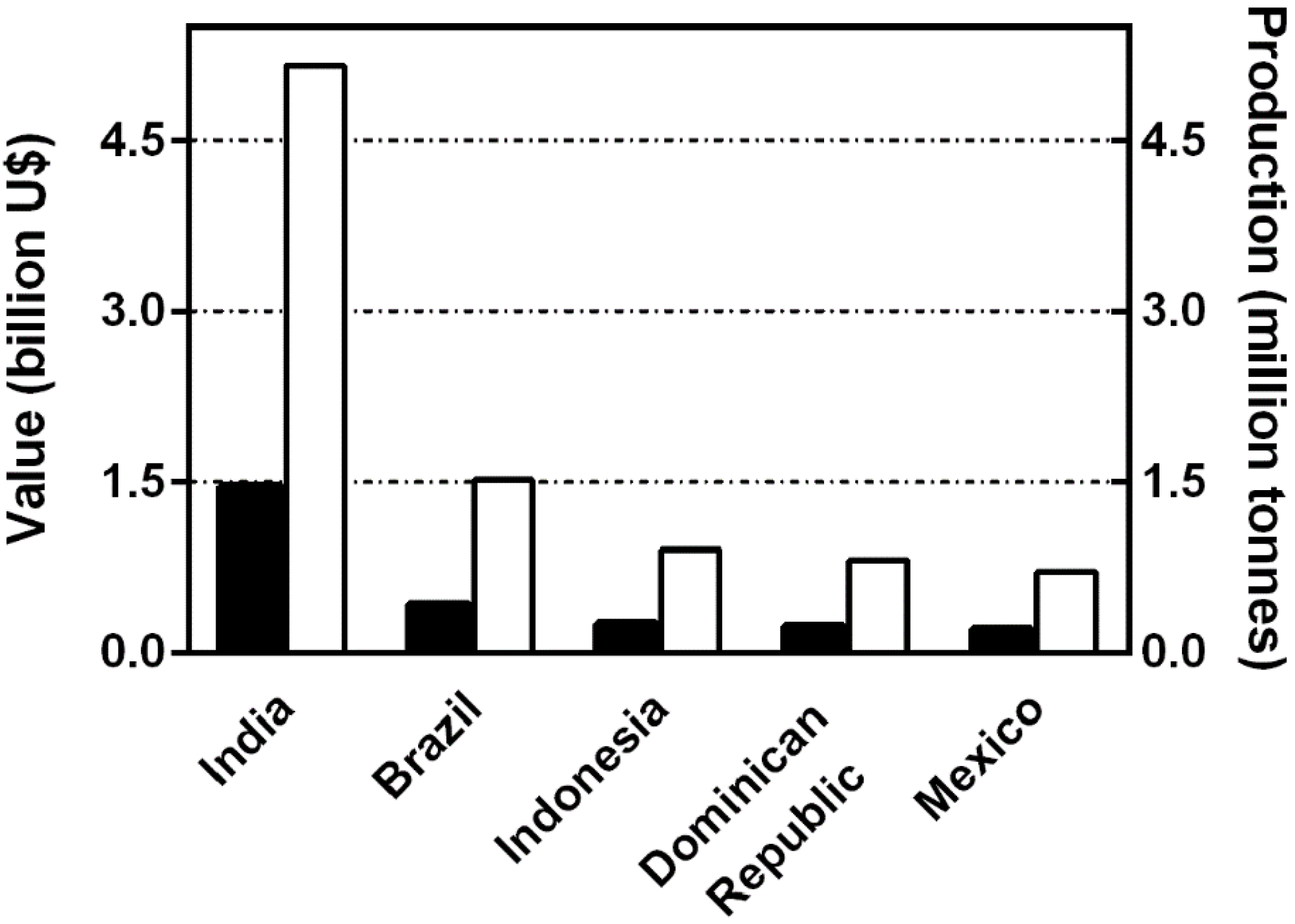
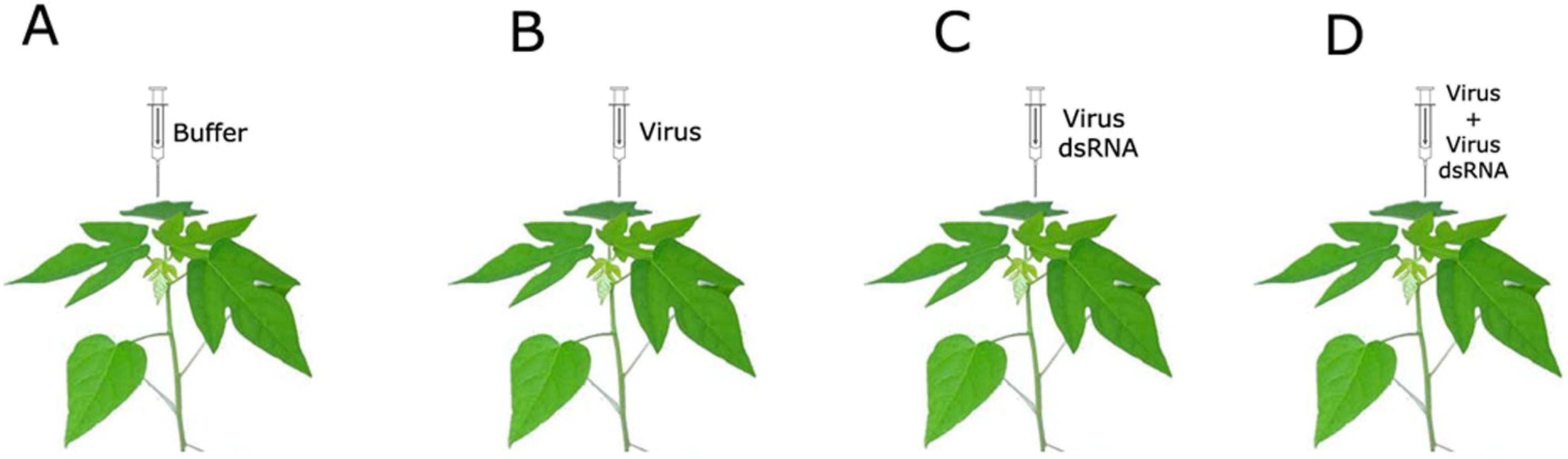
:1. Introduction

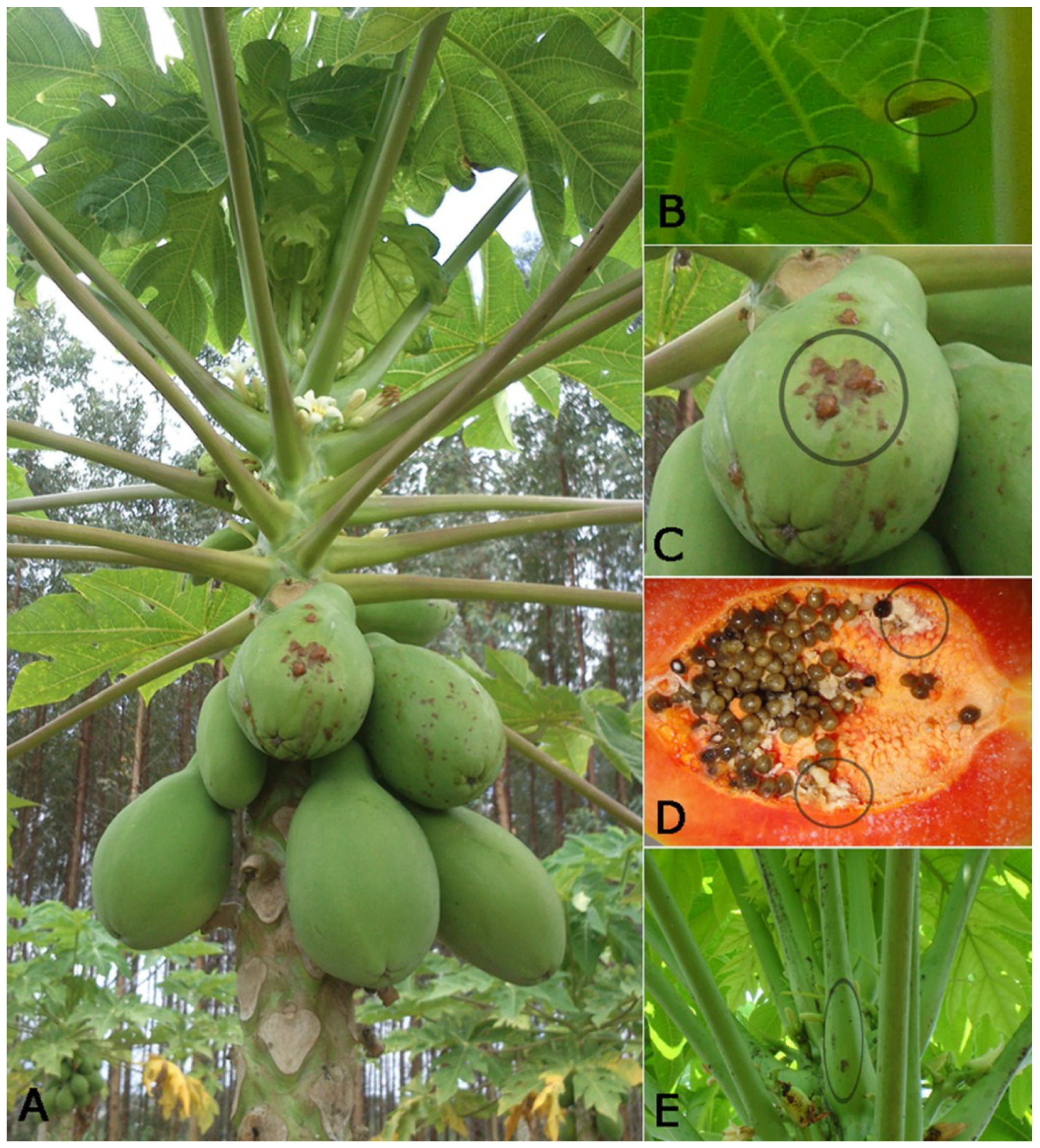
2. Papaya Meleira Virus
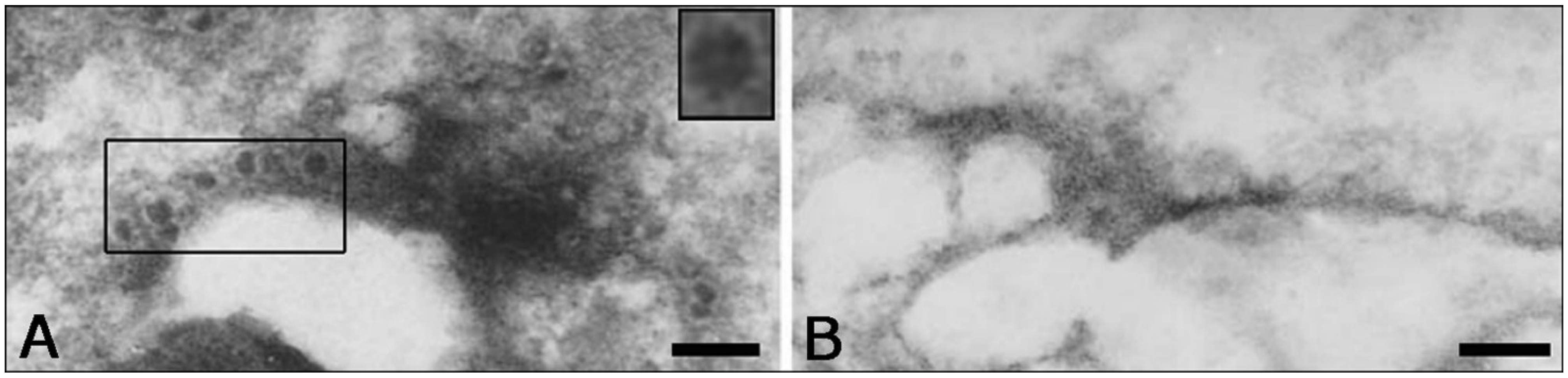
3. Etiology


4. Epidemiology

5. Detection Methods
6. Disease Management
7. Papaya and PMeV Interaction
8. Biotechnological Strategies to Papaya meleira virus Control

9. Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAOSTAT. FAOSTAT Production. Available online: http://faostat.fao.org/site/339/default.aspx (accessed on 19 December 2014).
- SIAP. Sistema de Información Agrícola y Pesquera (SIAP). Available online: http://www.siap.gob.mx/cierre-de-la-produccion-agricola-por-cultivo/ (accessed on 19 December 2014).
- Azad, M.A.K.; Amin, L.; Sidik, N.M. Gene technology for Papaya ringspot virus disease management. Sci. World J. 2014. [Google Scholar] [CrossRef]
- Bau, H.-J.; Kung, Y.-J.; Raja, J.; Chan, S.-J.; Chen, K.-C.; Chen, Y.-K.; Wu, H.-W.; Yeh, S.-D. Potential threat of a new pathotype of Papaya leaf distortion mosaic virus infecting transgenic papaya resistant to Papaya ringspot virus. Phytopathology 2008, 98, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Daltro, C.B.; PereiraII, Á.J.; Cascardo, R.S.; Alfenas-Zerbini, P.; Bezerra-Junior, J.E.A.; Lima, J.A.A.; Zerbini, F.M.; Andrade, E.C. Genetic variability of Papaya lethal yellowing virus isolates from Ceará and Rio Grande do Norte States, Brazil. Trop. Plant Pathol. 2012, 37. [Google Scholar] [CrossRef]
- Noa-Carrazana, J.; González-de-León, D.; Ruiz-Castro, B.; Piñero, D.; Silva-Rosales, L. Distribution of Papaya ringspot virus and Papaya mosaic virus in papaya plants (Carica papaya) in Mexico. Plant Dis. 2006, 90, 1004–1011. [Google Scholar] [CrossRef]
- Ventura, J.A.; Costa, H.; da Silva Tatagiba, J. Papaya diseases and integrated control. In Diseases of Fruits and Vegetables: Volume II; Springer: Berlin/Heidelberg, Germany, 2004; pp. 201–268. [Google Scholar]
- Teliz, D.; Mora, G.; Nieto, D.; Gonsalves, D.; Garcia, E.; Matheis, L.; Avila, C. La mancha anular del papayo en México. Rev. Mexicana Fitopatol. 1991, 9, 64–68. [Google Scholar]
- Ventura, J.A.; Costa, H.; Tatagiba, J.S.; Andrade, J.S.; Martins, D.S. Meleira do mamoeiro: Etiologia, sintomas e epidemiologia. In Papaya Brasil: Qualidade do Mamão Para o Mercado Interno; Martins, D.S., Ed.; Incaper: Vitória, Brazil, 2003; pp. 267–276. [Google Scholar]
- Rodrigues, C.H.; Ventura, J.A.; Maffia, L.A. Distribuição e transmissão da meleira em pomares de mamão no Espírito Santo. Fitopatol. Bras. 1989, 14, 118. [Google Scholar]
- Perez-Brito, D.; Tapia-Tussell, R.; Cortes-Velazquez, A.; Quijano-Ramayo, A.; Nexticapan-Garcez, A.; Martín-Mex, R. First report of Papaya meleira virus (PMeV) in Mexico. Afr. J. Biotechnol. 2012, 11, 13564–13570. [Google Scholar] [CrossRef]
- Daltro, C.B.; Abreu, E.F.M.; Aragão, F.J.L.; Andrade, E.C. Genetic diversity studies of Papaya meleira virus. Trop. Plant Pathol. 2014, 39, 104–108. [Google Scholar] [CrossRef]
- Araújo, M.M.M.; Tavares, E.T.; Silva, F.R.; Marinho, V.L.A.; Júnior, M.T.S. Molecular detection of Papaya meleira virus in the latex of Carica papaya by RT-PCR. J. Virol. Methods 2007, 146, 305–310. [Google Scholar] [CrossRef]
- Kitajima, E.W.; Rodrigues, C.H.; Silveira, J.S.; Alves, F.; Ventura, J.A.; Aragao, F.J.L.; Oliveira, L.H.R. Association of isometric virus like particles, restricted to laticifers, with “meleira” (“sticky disease”) of papaya (Carica papaya). Fitopatol. Bras. 1993, 18, 118–122. [Google Scholar]
- Maciel-Zambolim, E.; Kunieda-Alonso, S.; Matsuoka, K.; de Carvalho, M.; Zerbini, F. Purification and some properties of Papaya meleira virus, a novel virus infecting papayas in Brazil. Plant Pathol. 2003, 52, 389–394. [Google Scholar] [CrossRef]
- Nakagawa, J.T.Y.; Suzukama, Y. Exudação de Látex Pelo Mamoeiro. Estudo de Ocorrência em Teixeira de Freitas, BA; Congresso Brasileiro de Fruticultura: Brasil, 1987; pp. 555–559. [Google Scholar]
- Akiba, F. Bactérias pleomórficas, Gram negativas, e de crescimento lento em meio de cultura, isoladas do sistema vascular de diversas espécies de plantas apresentando sintomas de declínio. Fitopatol. Bras. 1989, 14, 110–111. [Google Scholar]
- Fauquet, C.M.; Fargette, D. International Committee on Taxonomy of Viruses and the 3142 unassigned species. Virol. J. 2005, 2, e64. [Google Scholar] [CrossRef]
- Rodrigues, S.P.; da Cunha, M.; Ventura, J.A.; Fernandes, P.M.B. Effects of the Papaya meleira virus on papaya latex structure and composition. Plant Cell Rep. 2009, 28, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.P.; Andrade, J.S.; Ventura, J.A.; Lindsey, G.G.; Fernandes, P.M.B. Papaya meleira virus is neither transmitted by infection at wound sites nor by the whitefly Trialeurodes variabilis. J. Plant Pathol. 2009, 91, 87–91. [Google Scholar]
- Magaña-Alvarez, A. Evaluación de la transmisión de Papaya Meleira Virus (PMeV) por Semillas. Maestría; Centro de Investigación Científica de Yucatán A.C.: Mexico, Mexico, 2013. [Google Scholar]
- Cosmi, F.C. Epidemiologia da Meleira do Mamoeiro no Norte do estado do Espírito Santo; Mestrado, UFES: Alegre, Espírito Santo, Brazil, 2010. [Google Scholar]
- Tapia-Tussell, R.; Magaña-Alvarez, A.; Cortes-Velazquez, A.; Itza-Kuk, G.; Nexticapan-Garcez, A.; Quijano-Ramayo, A.; Martin-Mex, R.; Perez-Brito, D. Seed transmission of Papaya meleira virus in papaya (Carica papaya) cv. Maradol. Plant Pathol. 2015, 64, 272–275. [Google Scholar] [CrossRef]
- Abreu, P.; Piccin, J.G.; Rodrigues, S.P.; Buss, D.S.; Ventura, J.A.; Fernandes, P. Molecular diagnosis of Papaya meleira virus (PMeV) from leaf samples of Carica papaya L. using conventional and real-time RT-PCR. J. Virol. Methods 2012, 180, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Forster, R.L.; Seifers, D.L.; Strausbaugh, C.A.; Jensen, S.G.; Ball, E.M.; Harvey, T.L. Seed transmission of the High Plains virus in sweet corn. Plant Dis. 2001, 85, 696–699. [Google Scholar] [CrossRef]
- Mikel, M.A.; Darcy, C.J.; Ford, R.E. Seed transmission of Maize dwarf mosaic virus in sweet corn. J. Phytopathol. 1984, 110, 185–191. [Google Scholar] [CrossRef]
- Zaitlin, M.; Palukaitis, P. Advances in understanding plant viruses and virus diseases. Annu. Rev. Phytopathol. 2000, 38, 117–143. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.A.; Laranjeira, F.F.; Nascimento, A.S.; Habibe, T.C. Distribuição espacial da meleira do mamoeiro em zonas de trópico úmido e trópico semi-árido. Fitopatol. Bras. 2004, 29, 276–281. [Google Scholar] [CrossRef]
- Vidal, C.A.; Nascimento, A.S.; Habibe, T.C. Transmissão do vírus da meleira do mamoeiro por insetos. In Papaya Brasil: Qualidade do Mamão Para o Mercado Interno; Martins, D.S., Ed.; Incaper: Vitória, Brazil, 2003; pp. 612–615. [Google Scholar]
- Picaço, M.C.; Moura, M.F.; Moreira, M.D. Biologia, identificação e manejo de moscas-brancas em fruteiras. In Manejo Integrado, Produção Integrada, Fruteiras Tropicais, Doenças e Pragas; Laercio, Z., Ed.; UVS Pulbisher: Viçosa, Brazil, 2003; pp. 243–284. [Google Scholar]
- Andrade, J.d.S.; Ventura, J.A.; Rodrigues, S.P.; Fernandes, P.M.B.; Tatagiba, J.D.S.; Costa, H. Avaliação de diferentes métodos de inoculação da meleira em plantas jovens de mamão. Fitopatol. Bras. 2003, 28, 288. [Google Scholar] [CrossRef]
- Martins, D.D.; Ventura, J.A.; Lima, R.D.A.; Culik, M.P.; Costa, H.; Ferreira, P.S.F. Interaction between Papaya meleira virus (PMeV) infection of papaya plants and mediterranean fruit fly infestation of fruits. Crop. Prot. 2012, 36, 7–10. [Google Scholar] [CrossRef]
- Rodrigues, S.P.G.O.P.; Andrade, J.S.; Ventura, J.A.; Fernandes, P.M.B. Simplified molecular method for the diagnosis of Papaya meleira virus in papaya latex and tissues. Summa Phytopathol. 2005, 31, 281–283. [Google Scholar]
- López-Ochoa, L.A.; Moreno-Valenzuela, O.A.; Zamudio-Moreno, E. Secuencia parcial y metodo de diagnóstico por RT-PCR del virus de la meleira de la papaya. Mx Patent 2012000081A; filed 16 Dec 2011, and issue June 2013,
- Habibe, T.C.; Dantas, J.L.L.; do Nascimento, A.S.; Dantas, A.C.V.L. Suscetibilidade de Genótipos de Mamoeiro (Carica Papaya L.) ao Vírus da Meleira, Sob Condições de Trópico Semi-Árido; Embrapa Mandioca e Fruticultura: Cruz Das Almas, Brazil, 2003. [Google Scholar]
- Dos S. Martins, D. Papaya Brasil: Manejo, Qualidade e Mercado do Mamão; Incaper: Vitória, Brazil, 2007. [Google Scholar]
- El Moussaoui, A.; Nijs, M.; Paul, C.; Wintjens, R.; Vincentelli, J.; Azarkan, M.; Looze, Y. Revisiting the enzymes stored in the laticifers of Carica papaya in the context of their possible participation in the plant defence mechanism. Cell Mol. Life Sci. 2001, 58, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L.M.; Hermosilla, J.; Salas, C.E. A biochemical comparison between latex from Carica candamarcensis and C. Papaya. Braz. J. Med. Biol. Res. 1994, 27, 2831–2842. [Google Scholar] [PubMed]
- Moutim, V.; Silva, L.G.; Lopes, M.T.P.; Fernandes, G.W.; Salas, C.E. Spontaneous processing of peptides during coagulation of latex from Carica papaya. Plant Sci. 1999, 142, 115–121. [Google Scholar] [CrossRef]
- Rodrigues, S.P.; Ventura, J.A.; Aguilar, C.; Nakayasu, E.S.; Choi, H.; Sobreira, T.J.P.; Nohara, L.L.; Wermelinger, L.S.; Almeida, I.C.; Zingali, R.B.; et al. Label-free quantitative proteomics reveals differentially regulated proteins in the latex of sticky diseased Carica papaya L. plants. J. Proteomics 2012, 75, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Konno, K. Plant latex and other exudates as plant defense systems: Roles of various defense chemicals and proteins contained therein. Phytochemistry 2011, 72, 1510–1530. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hirayama, C.; Nakamura, M.; Tateishi, K.; Tamura, Y.; Hattori, M.; Kohno, K. Papain protects papaya trees from herbivorous insects: Role of cysteine proteases in latex. Plant J. 2004, 37, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Azarkan, M.; Dibiani, R.; Baulard, C.; Baeyens-Volant, D. Effects of mechanical wounding on Carica papaya cysteine endopeptidases accumulation and activity. Int. J. Biol. Macromol. 2006, 38, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Azarkan, M.; el Moussaoui, A.; van Wuytswinkel, D.; Dehon, G.; Looze, Y. Fractionation and purification of the enzymes stored in the latex of Carica papaya. J. Chromatogr. B 2003, 790, 229–238. [Google Scholar] [CrossRef]
- Solomon, M.; Belenghi, B.; Delledonne, M.; Menachem, E.; Levine, A. The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 1999, 11, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.T.; Yao, N. The role and regulation of programmed cell death in plant-pathogen interactions. Cell Microbiol. 2004, 6, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [PubMed]
- Buss, D.S.; Dias, G.B.; Santos, M.P.; Ventura, J.A.; Fernandes, P.M.B. Oxidative stress defence response of Carica papaya challenged by nitric oxide, Papaya meleira virus and Saccharomyces cerevisiae. Open Nitric Oxide J. 2011, 3, 55–64. [Google Scholar] [CrossRef]
- Rodrigues, S.P.; Ventura, J.A.; Aguilar, C.; Nakayasu, E.S.; Almeida, I.C.; Fernandes, P.; Zingali, R.B. Proteomic analysis of papaya (Carica papaya L.) displaying typical sticky disease symptoms. Proteomics 2011, 11, 2592–2602. [Google Scholar] [CrossRef] [PubMed]
- Citovsky, V.; Zaltsman, A.; Kozlovsky, S.V.; Gafni, Y.; Krichevsky, A. Proteasomal degradation in plant-pathogen interactions. Semin. Cell Dev. Biol. 2009, 20, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Dielen, A.S.; Badaoui, S.; Candresse, T.; German-Retana, S. The ubiquitin/26S proteasome system in plant-pathogen interactions: A never-ending hide-and-seek game. Mol. Plant Pathol. 2010, 11, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.; Peeters, N.; Rivas, S. Ubiquitination during plant immune signaling. Plant Physiol. 2012, 160, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Alcaide-Loridan, C.; Jupin, I. Ubiquitin and plant viruses, let’s play together! Plant Physiol. 2012, 160, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, M.K.; Ploegh, H.L. Ubiquitination, ubiquitin-like modifiers, and deubiquitination in viral infection. Cell Host Microbe 2009, 5, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Randow, F.; Lehner, P.J. Viral avoidance and exploitation of the ubiquitin system. Nat. Cell Biol. 2009, 11, U527–U535. [Google Scholar] [CrossRef]
- Park, W.; Li, J.J.; Song, R.T.; Messing, J.; Chen, X.M. CARPEL FACTORY, a Dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana. Curr. Biol. 2002, 12, 1484–1495. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Weinstein, E.G.; Rhoades, M.W.; Bartel, B.; Bartel, D.P. MicroRNAs in plants. Gene Dev. 2002, 16, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Aryal, R.; Yang, X.Z.; Yu, Q.Y.; Sunkar, R.; Li, L.; Ming, R. Asymmetric purine-pyrimidine distribution in cellular small RNA population of papaya. BMC Genomics 2012, 13, 682. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Li, Y.; He, H.; Wang, F.; Yu, D.Q. Identification of miRNAs and miRNA-mediated regulatory pathways in Carica papaya. Planta 2013, 238, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Abreu, P.M.V.; Gaspar, C.G.; Buss, D.S.; Ventura, J.A.; Ferreira, P.C.G.; Fernandes, P.M.B. Carica papaya microRNAs are responsive to Papaya meleira virus infection. PLoS ONE 2014, 9, e103401. [Google Scholar] [CrossRef] [PubMed]
- Tagami, Y.; Inaba, N.; Kutsuna, N.; Kurihara, Y.; Watanabe, Y. Specific enrichment of miRNAs in Arabidopsis thaliana infected with Tobacco mosaic virus. DNA Res. 2007, 14, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Bazzini, A.A.; Hopp, H.E.; Beachy, R.N.; Asurmendi, S. Infection and coaccumulation of Tobacco mosaic virus proteins alter microRNA levels, correlating with symptom and plant development. Proc. Natl. Acad. Sci. USA 2007, 104, 12157–12162. [Google Scholar] [CrossRef] [PubMed]
- Gonsalves, D.; Vegas, A.; Prasartsee, V.; Drew, R.A.; Suzuki, J.Y.; Tripathi, S. Developing papaya to control Papaya ringspot virus by transgenic resistance, intergeneric hybridization, and tolerance breeding. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006; Volume 26, pp. 35–73. [Google Scholar]
- Scorza, R.; Callahan, A.; Levy, L.; Damsteegt, V.; Webb, K.; Ravelonandro, M. Post-transcriptional gene silencing in plum pox virus resistant transgenic European plum containing the plum pox potyvirus coat protein gene. Transgenic Res. 2001, 10, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Tricoli, D.M.; Carney, K.J.; Russell, P.F.; Mcmaster, J.R.; Groff, D.W.; Hadden, K.C.; Himmel, P.T.; Hubbard, J.P.; Boeshore, M.L.; Quemada, H.D. Field evaluation of transgenic squash containing single or multiple virus coat protein gene constructs for resistance to Cucumber Mosaic Virus. Biotechnology 1995, 13, 1458–1465. [Google Scholar] [CrossRef]
- Tenllado, F.; Martinez-Garcia, B.; Vargas, M.; Diaz-Ruiz, J.R. Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections. BMC Biotechnol. 2003, 3, e3. [Google Scholar] [CrossRef] [Green Version]
- Tenllado, F.D.-R.J.R. Double-stranded RNA-mediated interference with plant virus infection. J. Virol. 2001, 75, 12288–12297. [Google Scholar] [CrossRef] [PubMed]
- Nicaise, V. Crop immunity against viruses: Outcomes and future challenges. Front. Plant Sci. 2014, 5, e660. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Alazem, M.; Lin, K.Y.; Lin, N.S. The abscisic acid pathway has multifaceted effects on the accumulation of Bamboo mosaic virus. Mol. Plant Microbe Interact 2014, 27, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, L.; Li, D.; Wang, F.; Yu, D. WRKY8 transcription factor functions in the TMV-cg defense response by mediating both abscisic acid and ethylene signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, E1963–E1971. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abreu, P.M.V.; Antunes, T.F.S.; Magaña-Álvarez, A.; Pérez-Brito, D.; Tapia-Tussell, R.; Ventura, J.A.; Fernandes, A.A.R.; Fernandes, P.M.B. A Current Overview of the Papaya meleira virus, an Unusual Plant Virus. Viruses 2015, 7, 1853-1870. https://doi.org/10.3390/v7041853
Abreu PMV, Antunes TFS, Magaña-Álvarez A, Pérez-Brito D, Tapia-Tussell R, Ventura JA, Fernandes AAR, Fernandes PMB. A Current Overview of the Papaya meleira virus, an Unusual Plant Virus. Viruses. 2015; 7(4):1853-1870. https://doi.org/10.3390/v7041853
Chicago/Turabian StyleAbreu, Paolla M. V., Tathiana F. S. Antunes, Anuar Magaña-Álvarez, Daisy Pérez-Brito, Raúl Tapia-Tussell, José A. Ventura, Antonio A. R. Fernandes, and Patricia M. B. Fernandes. 2015. "A Current Overview of the Papaya meleira virus, an Unusual Plant Virus" Viruses 7, no. 4: 1853-1870. https://doi.org/10.3390/v7041853
APA StyleAbreu, P. M. V., Antunes, T. F. S., Magaña-Álvarez, A., Pérez-Brito, D., Tapia-Tussell, R., Ventura, J. A., Fernandes, A. A. R., & Fernandes, P. M. B. (2015). A Current Overview of the Papaya meleira virus, an Unusual Plant Virus. Viruses, 7(4), 1853-1870. https://doi.org/10.3390/v7041853