
Dynamics of Apis mellifera Filamentous Virus (AmFV) Infections in Honey Bees and Relationships with Other Parasites

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Honey Bee Samples
2.2. DNA Extraction and PCR Analysis
2.3. Statistical Analyses
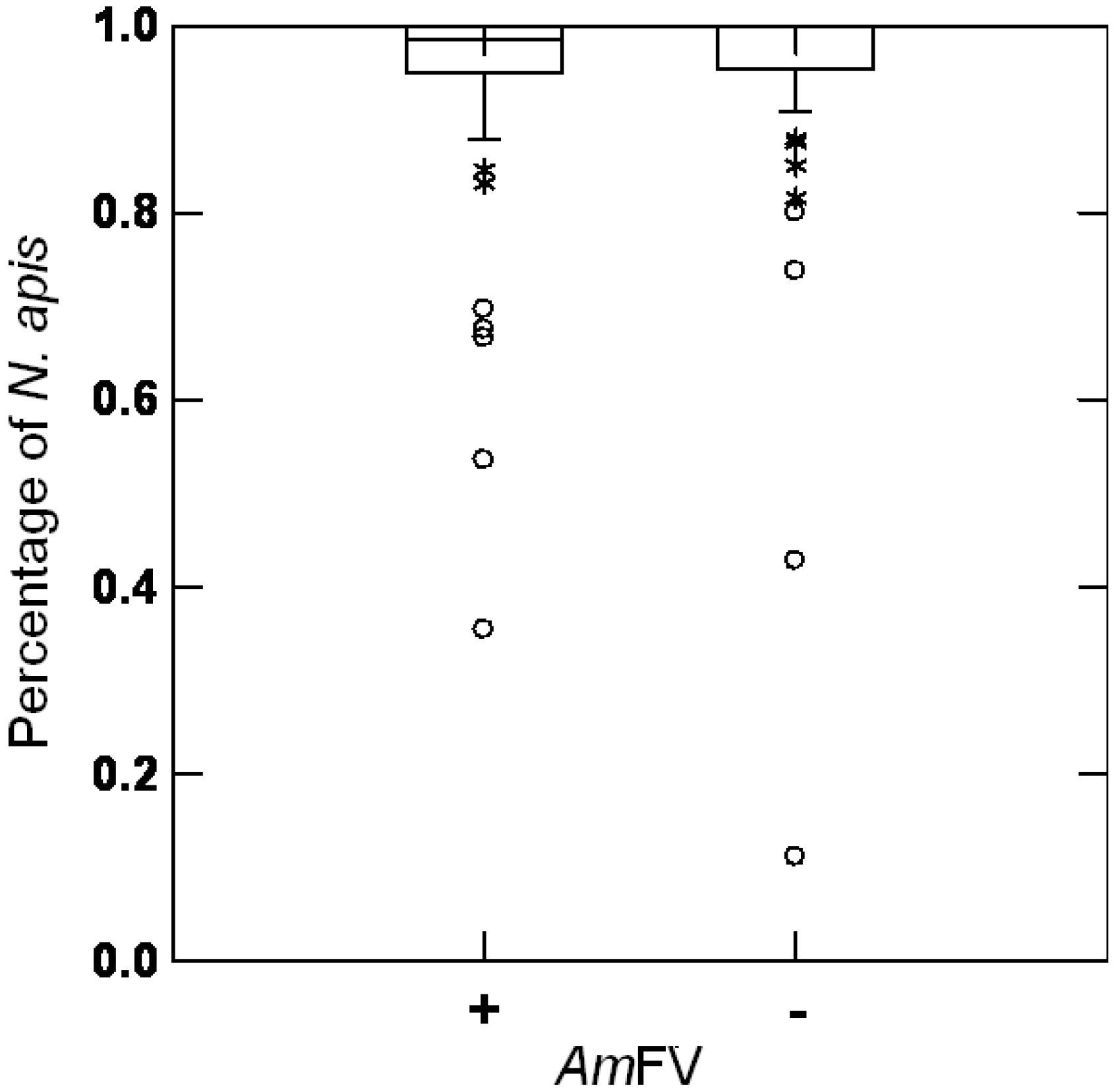
3. Results
3.1. Kinetics of Honey Bee Gut Colonization by Parasites

{kind=link}
| GUTS | AmFV | N. ceranae | Trypanosomes | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Weeks p.e. | 0 | 1 | 2 | 3 | WB | 0 | 1 | 2 | 3 | WB | 0 | 1 | 2 | 3 | WB |
| Colony A | + | + | + | + | + | ||||||||||
| + | + | + | + | ||||||||||||
| + | n | + | + | n | + | + | n | ||||||||
| + | + | + | + | + | + | + | |||||||||
| + | + | + | + | + | + | + | + | + | |||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | + | + | ||||||||
| Colony B | + | + | + | + | + | + | + | ||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | + | |||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | + | |||||||||
| + | + | + | + | + | |||||||||||
| + | + | + | + | + | |||||||||||
| + | + | + | + | + | |||||||||||
| Colony C | + | + | + | + | + | + | + | ||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | ||||||||||||
| + | + | + | + | + | + | + | + | + | + | ||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | ||||||||||
| pos. (%) | 0 | 75 | 65 | 63 | 96 | 0 | 17 | 30 | 71 | 83 | 0 | 0 | 30 | 42 | 54 |
| BODIES | AmFV | N. ceranae | Trypanosomes | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Weeks p.e. | 0 | 1 | 2 | 3 | WB | 0 | 1 | 2 | 3 | WB | 0 | 1 | 2 | 3 | WB |
| Colony A | + | + | + | + | |||||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | + | |||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | + | |||||||||
| + | + | + | + | + | + | ||||||||||
| + | + | + | + | + | + | + | + | ||||||||
| Colony B | + | + | + | + | + | + | + | ||||||||
| + | + | + | + | + | + | + | + | + | + | ||||||
| + | + | + | + | + | + | + | |||||||||
| + | + | + | + | + | + | + | + | ||||||||
| + | + | + | + | + | + | + | |||||||||
| + | + | + | + | + | + | + | + | + | |||||||
| + | + | + | + | + | + | + | + | + | |||||||
| + | + | + | + | + | + | + | + | ||||||||
| Colony C | + | + | + | + | + | + | + | + | + | ||||||
| + | + | + | + | + | + | + | + | + | |||||||
| + | + | + | + | + | + | + | + | + | |||||||
| + | + | + | + | + | + | + | + | + | + | ||||||
| + | + | + | + | + | + | + | + | + | |||||||
| + | + | + | + | + | + | + | + | ||||||||
| + | + | + | + | + | + | + | + | ||||||||
| + | + | + | + | + | |||||||||||
| pos. (%) | 0 | 100 | 79 | 87 | 71 | 0 | 75 | 37 | 100 | 87 | 0 | 12 | 33 | 58 | 17 |
3.2. Relationships between AmFV and Honey Bee Parasites at the Colony Level

4. Discussion
AmFV Distribution in Honey Bee Colonies
5. Conclusions
Acknowledgments
Author Contributions
Supplementary
Conflicts of Interest
References
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef] [Green Version]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Potts, S.G.; Schaffers, A.P.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators an insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; Vanengelsdorp, D.; Pettis, J.S. High Levels of Miticides and Agrochemicals in North American Apiaries: Implications for Honey Bee Health. PLoS ONE 2010, 5, 1–19. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Drivers and impacts. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, 96–119. [Google Scholar] [CrossRef] [PubMed]
- Lounsberry, Z.; Spiewok, S.; Pernal, S.F.; Sonstegard, T.S.; Hood, W.M.; Pettis, J.; Neumann, P.; Evans, J.D. Worldwide Diaspora of Aethina tumida (Coleoptera: Nitidulidae), a Nest Parasite of Honey Bees. Ann. Entomol. Soc. Am. 2010, 103, 671–677. [Google Scholar] [CrossRef]
- Johnson, R.M.; Ellis, M.D.; Mullin, C.A.; Frazier, M. Pesticides and honey bee toxicity—USA. Apidologie 2010, 41, 312–331. [Google Scholar] [CrossRef]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed]
- Vidau, C.; Diogon, M.; Aufauvre, J.; Fontbonne, R.; Viguès, B.; Brunet, J.L.; Texier, C.; Biron, D.G.; Blot, N.; El Alaoui, H.; et al. Exposure to sublethal doses of fipronil and thiacloprid highly increases mortality of honeybees previously infected by Nosema ceranae. PLoS ONE 2011, 6, e21550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.Y.; Smart, M.D.; Anelli, C.M.; Sheppard, W.S. Honey bees (Apis mellifera) reared in brood combs containing high levels of pesticide residues exhibit increased susceptibility to Nosema (Microsporidia) infection. J. Invertebr. Pathol. 2012, 109, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Pettis, J.S.; vanEngelsdorp, D.; Johnson, J.; Dively, G. Pesticide exposure in honey bees results in increased levels of the gut pathogen Nosema. Naturwissenschaften 2012, 99, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Aufauvre, J.; Biron, D.G.; Vidau, C.; Fontbonne, R.; Roudel, M.; Diogon, M.; Viguès, B.; Belzunces, L.P.; Delbac, F.; Blot, N. Parasite-insecticide interactions: A case study of Nosema ceranae and fipronil synergy on honeybee. Sci. Rep. 2012, 2, 326. [Google Scholar] [CrossRef] [PubMed]
- Doublet, V.; Labarussias, M.; de Miranda, J.R.; Moritz, R.F.; Paxton, R.J. Bees under stress: Sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 2015, 17, 969–983. [Google Scholar] [CrossRef] [PubMed]
- Tanada, Y.; Kaya, H.K. Insect Pathology; Academic Press: San Diego, CA, USA, 1993; pp. 147–153. [Google Scholar]
- De Miranda, J.; Chen, Y.; Ribière, M.; Gauthier, L. Varroa and Viruses. In Varroa—Still a Problem in the 21st Century; Carreck, N., Ed.; IBRA: Worcester, UK, 2011. [Google Scholar]
- Clark, T.B. A filamentous virus of the honey bee. J. Invertebr. Pathol. 1978, 32, 332–340. [Google Scholar] [CrossRef]
- Gauthier, L.; Cornman, S.; Hartmann, U.; Cousserans, F.; Evans, J.D.; de Miranda, J.R.; Neumann, P. The Apis mellifera filamentous virus genome. Submitted.
- Sitaropoulou, N.; Neophytou, E.P.; Thomopoulos, G.N. Structure of the Nucleocapsid of A Filamentous Virus of the Honey Bee (Apis mellifera). J. Invertebr. Pathol. 1989, 53, 354–357. [Google Scholar] [CrossRef]
- Bailey, L.; Carpenter, J.M.; Woods, R.D. Properties of A Filamentous Virus of the Honey Bee (Apis mellifera). Virology 1981, 114, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ravoet, J.; de Smet, L.; Meeus, I.; Smagghe, G.; Wenseleers, T.; de Graaf, D.C. Widespread occurrence of honey bee pathogens in solitary bees. J. Invertebr. Pathol. 2014, 122, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Clark, T.B. Another virus in the honey bees. Am. Bee J. 1977, 117, 340–341. [Google Scholar]
- Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 103, 73–79. [Google Scholar] [CrossRef]
- Cornman, R.S.; Chen, Y.P.; Schatz, M.C.; Street, C.; Zhao, Y.; Desany, B.; Egholm, M.; Hutchison, S.; Pettis, J.S.; Lipkin, W.I.; et al. Genomic Analyses of the Microsporidian Nosema ceranae, an Emergent Pathogen of Honey Bees. PLoS Pathog. 2009, 5, e1000466. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.S.; Bauchan, G.R.; Murphy, C.A.; Ravoet, J.; de Graaf, D.C.; Evans, J.D. Characterization of Two Species of Trypanosomatidae from the Honey Bee Apis mellifera: Crithidia mellificae Langridge and McGhee, and Lotmaria passim n. gen., n. sp. J. Eukaryot. Microbiol. 2015. [Google Scholar] [CrossRef]
- Runckel, C.; DeRisi, J.; Flenniken, M.L. A Draft Genome of the Honey Bee Trypanosomatid Parasite Crithidia mellificae. PLoS ONE 2014, 9, e95057. [Google Scholar] [CrossRef] [PubMed]
- Bailey, L.; Ball, B.V.; Perry, J.N. Association of viruses with two protozoal pathogens of the honey bee. Ann. Appl. Biol. 1983, 103, 13–20. [Google Scholar] [CrossRef]
- Bailey, L.; Ball, B.V. Honey Bee Pathology; Academic Press: London, UK, 1991. [Google Scholar]
- Chen, Y.P.; Siede, R. Honey bee viruses. Adv. Virus Res. 2007, 70, 33–80. [Google Scholar] [PubMed]
- Zander, E. Tierische Parasiten als Krankheitserreger bei der Biene. Leipz. Bienen Ztg. 1909, 24, 147–150. [Google Scholar]
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chinh, T.X.; Puerta, F.; Ruz, J.M.; Kryger, P.; et al. Widespread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera. J. Invertebr. Pathol. 2007, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Huang, Z.Y. Nosema ceranae, a newly identified pathogen of Apis mellifera in the USA and Asia. Apidologie 2010, 41, 364–374. [Google Scholar] [CrossRef]
- Forsgren, E.; Fries, I. Comparative virulence of Nosema ceranae and Nosema apis in individual European honey bees. Vet. Parasitol. 2010, 170, 212–217. [Google Scholar] [CrossRef] [PubMed]
- VanEngelsdorp, D.; Hayes, J.; Underwood, R.M.; Pettis, J. A Survey of Honey Bee Colony Losses in the US, Fall 2007 to Spring 2008. PLoS ONE 2008, 3, e4071. [Google Scholar] [CrossRef] [PubMed]
- Paxton, R.J. Does infection by Nosema ceranae cause “Colony Collapse Disorder” in honey bees (Apis mellifera)? J. Apic. Res. 2010, 49, 80–84. [Google Scholar] [CrossRef]
- Genersch, E.; von der Ohe, W.; Kaatz, H.; Schroeder, A.; Otten, C.; Büchler, R.; Berg, S.; Ritter, W.; Mühlen, W.; Gisder, S.; et al. The German bee monitoring project: A long term study to understand periodically high winter losses of honey bee colonies. Apidologie 2010, 41, 332–352. [Google Scholar] [CrossRef]
- Stevanovic, J.; Stanimirovic, Z.; Genersch, E.; Kovacevic, S.R.; Ljubenkovic, J.; Radakovic, M.; Aleksic, N. Dominance of Nosema ceranae in honey bees in the Balkan countries in the absence of symptoms of colony collapse disorder. Apidologie 2011, 42, 49–58. [Google Scholar] [CrossRef]
- Higes, M.; Martin-Hernandez, R.; Botias, C.; Bailon, E.G.; Gonzalez-Porto, A.V.; Barrios, L.; del Nozal, M.J.; Bernal, J.L.; Jimenez, J.J.; Palencia, P.G.; et al. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Runckel, C.; Flenniken, M.L.; Engel, J.C.; Ruby, J.G.; Ganem, D.; Andino, R.; Derisi, J.L. Temporal Analysis of the Honey Bee Microbiome Reveals Four Novel Viruses and Seasonal Prevalence of Known Viruses, Nosema, and Crithidia. PLoS ONE 2011, 6, e20656. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Sheehan, T.H.; Eckholm, B.J.; Mott, B.M.; DeGrandi-Hoffman, G. An emerging paradigm of colony health: Microbial balance of the honey bee and hive (Apis mellifera). Insect. Soc. 2011, 58, 431–444. [Google Scholar] [CrossRef]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef] [PubMed]
- Forsgren, E; Fries, I. Temporal study of Nosema ssp in a cold climate. Environ. Microbiol. Rep. 2013, 5, 72–78. [Google Scholar]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.B.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucl. Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, A.L.; Rinderer, T.E.; Beaman, L.D.; Danka, R.G. Genetic detection and quantification of Nosema apis and N. ceranae in the honey bee. J. Invertebr. Pathol. 2010, 103, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, A.P.; Mackert, A.; Cristino, A.D.; Simoes, Z.L.P. Validation of reference genes for gene expression studies in the honey bee, Apis mellifera, by quantitative real-time RT-PCR. Apidologie 2008, 39, 372–385. [Google Scholar] [CrossRef]
- Gauthier, L.; Tentcheva, D.; Tournaire, M.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Viral load estimation in asymptomatic honey bee colonies using the quantitative RT-PCR technique. Apidologie 2007, 38, 426–437. [Google Scholar] [CrossRef]
- Powell, J.E.; Martinson, V.G.; Urban-Mead, K.; Moran, N.A. Routes of acquisition of the gut microbiota of Apis mellifera. Appl. Environ. Microbiol. 2014, 80, 7378–7387. [Google Scholar] [CrossRef]
- Engel, P.; Martinson, V.G.; Moran, N.A. Functional diversity within the simple gut microbiota of the honey bee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef] [PubMed]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; vanEngelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [Google Scholar] [CrossRef] [PubMed]
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumann, P. Predictive markers of honey bee colony collapse. PLoS One 2012, 7, e32151. [Google Scholar] [CrossRef] [PubMed]
- Retschnig, G.; Williams, G.R.; Odemer, R.; Boltin, J.; di Poto, C.; Mehmann, M.M.; Retschnig, P.; Winiger, P.; Rosenkranz, P.; Neumann, P. Effects, but no interactions, of ubiquitous pesticide and parasite stressors on honey bee (Apis mellifera) lifespan and behaviour in a colony environment. Environ. Microbiol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.R.; Shutler, D.; Little, C.M.; Burgher-MacLellan, K.L.; Rogers, R.E.L. The microsporidian Nosema ceranae, the antibiotic Fumagilin-B (R), and western honey bee (Apis mellifera) colony strength. Apidologie 2011, 42, 15–22. [Google Scholar] [CrossRef]
- Wallace, F.C. The trypanosomatid parasites of insects and arachnids. Exp. Parasitol. 1966, 18, 124–193. [Google Scholar] [CrossRef] [PubMed]
- Langridge, D.F.; McGhee, R.B. Crithidia mellificae n. sp. an acidophilic trypanosomatid of honey bee Apis mellifera. J. Protozool. 1967, 14, 485–487. [Google Scholar] [CrossRef] [PubMed]
- Ravoet, J.; Maharramov, J.; Meeus, I.; de Smet, L.; Wenseleers, T.; Smagghe, G.; de Graaf, D.C. Comprehensive bee pathogen screening in Belgium reveals Crithidia mellificae as a new contributory factor to winter mortality. PLoS ONE 2013, 8, e72443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shykoff, J.A.; Schmid-Hempel, P. Parasites delay worker reproduction in bumblebees: Consequences for eusociality. Behav. Ecol. 1991, 2, 242–248. [Google Scholar] [CrossRef]
- Schluns, H.; Sadd, B.M.; Schmid-Hempel, P.; Crozier, R.H. Infection with the trypanosome Crithidia bombi and expression of immune-related genes in the bumblebee Bombus terrestris. Dev. Comp. Immunol. 2010, 34, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Hempel, R.; Schmid-Hempel, P. Colony performance and immunocompetence of a social insect, Bombus terrestris, in poor and variable environments. Funct. Ecol. 1998, 12, 22–30. [Google Scholar] [CrossRef]
- Locke, B.; Forsgren, E.; Fries, I.; de Miranda, J.R. Acaricide treatment affects viral dynamics in Varroa destructor-infested honey bee colonies via both host physiology and mite control. Appl. Environ. Microbiol. 2012, 78, 227–235. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hartmann, U.; Forsgren, E.; Charrière, J.-D.; Neumann, P.; Gauthier, L. Dynamics of Apis mellifera Filamentous Virus (AmFV) Infections in Honey Bees and Relationships with Other Parasites. Viruses 2015, 7, 2654-2667. https://doi.org/10.3390/v7052654
Hartmann U, Forsgren E, Charrière J-D, Neumann P, Gauthier L. Dynamics of Apis mellifera Filamentous Virus (AmFV) Infections in Honey Bees and Relationships with Other Parasites. Viruses. 2015; 7(5):2654-2667. https://doi.org/10.3390/v7052654
Chicago/Turabian StyleHartmann, Ulrike, Eva Forsgren, Jean-Daniel Charrière, Peter Neumann, and Laurent Gauthier. 2015. "Dynamics of Apis mellifera Filamentous Virus (AmFV) Infections in Honey Bees and Relationships with Other Parasites" Viruses 7, no. 5: 2654-2667. https://doi.org/10.3390/v7052654
APA StyleHartmann, U., Forsgren, E., Charrière, J. -D., Neumann, P., & Gauthier, L. (2015). Dynamics of Apis mellifera Filamentous Virus (AmFV) Infections in Honey Bees and Relationships with Other Parasites. Viruses, 7(5), 2654-2667. https://doi.org/10.3390/v7052654