
The Effect of Oral Administration of dsRNA on Viral Replication and Mortality in Bombus terrestris




Abstract
:1. Introduction
2. Materials and Methods
2.1. Viral Stock
2.2. IAPV Infectivity and Mortality
2.3. dsRNA Administration and Effects on Infectivity and Mortality of IAPV Infected Bees
2.4. Extraction and Virus Quantification

{kind=link}
{kind=link}
{kind=link}
| Target | Use | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Reference |
|---|---|---|---|---|
| IAPV | PCR | CGAACTTGGTGACTTGAAGG | GCATCAGTCGTCTTCCAGGT | [30] |
| PPIA | RT-qPCR | TCGTAATGGAGTTGAGGAGTGA | CTTGGCACATGAAGTTTGGAAT | [31] |
| IAPV | RT-qPCR | CCATGCCTGGCGATTCAC | CTGAATAATACTGTGCGTATC | [31] |
| dsVP586 | dsRNA | ACCTGGAAGACGATTGATGC | CTGCCCACTTCCAAACAACT | This study |
| dsVP443 | dsRNA | TATAGATGCCGCTCCATGTG | CTGCCCACTTCCAAACAACT | This study |
| dsVP293 | dsRNA | ACCTGGAAGACGATTGATGC | GTGGGTTTGACGGGTATCAC | This study |
2.5. dsRNA Synthesis
3. Results
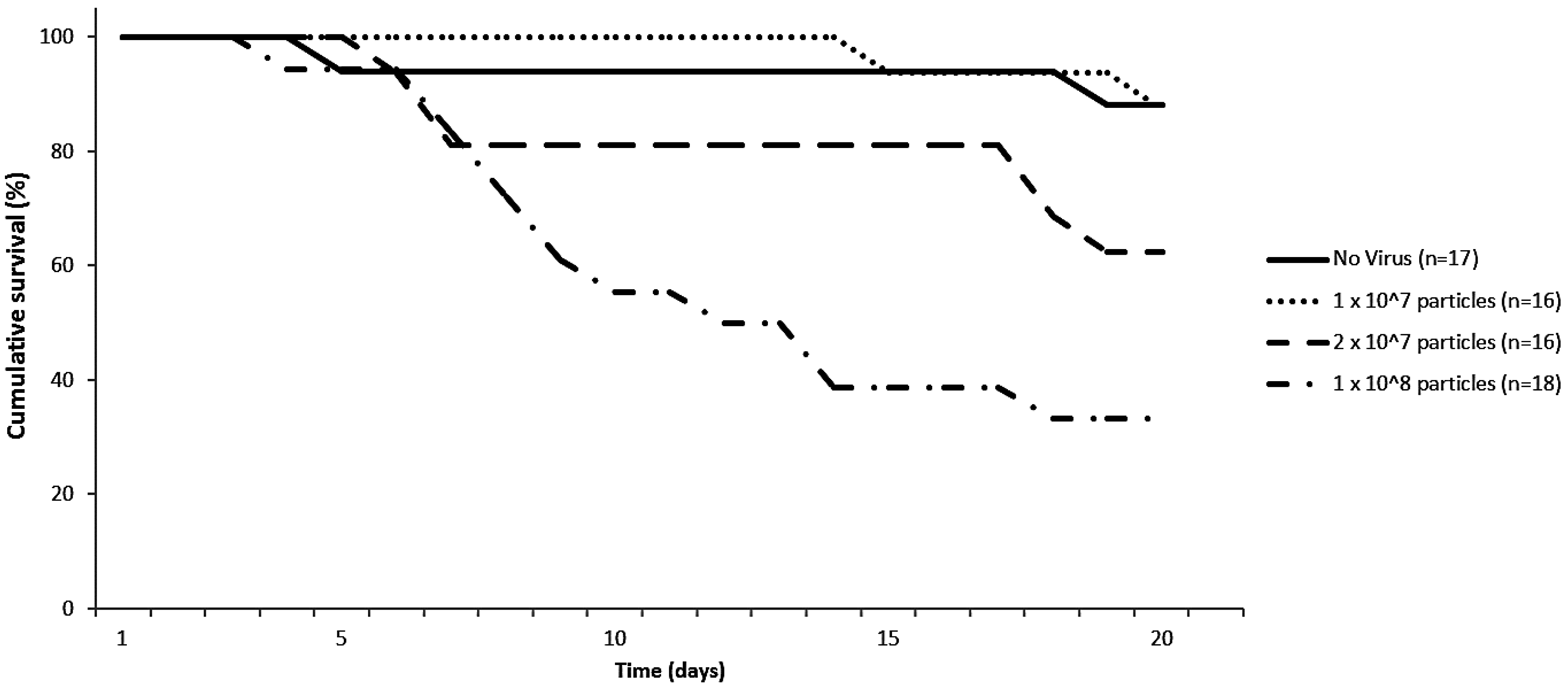
3.1. Infectivity of IAPV in Bumblebees and Its Effect on Survival

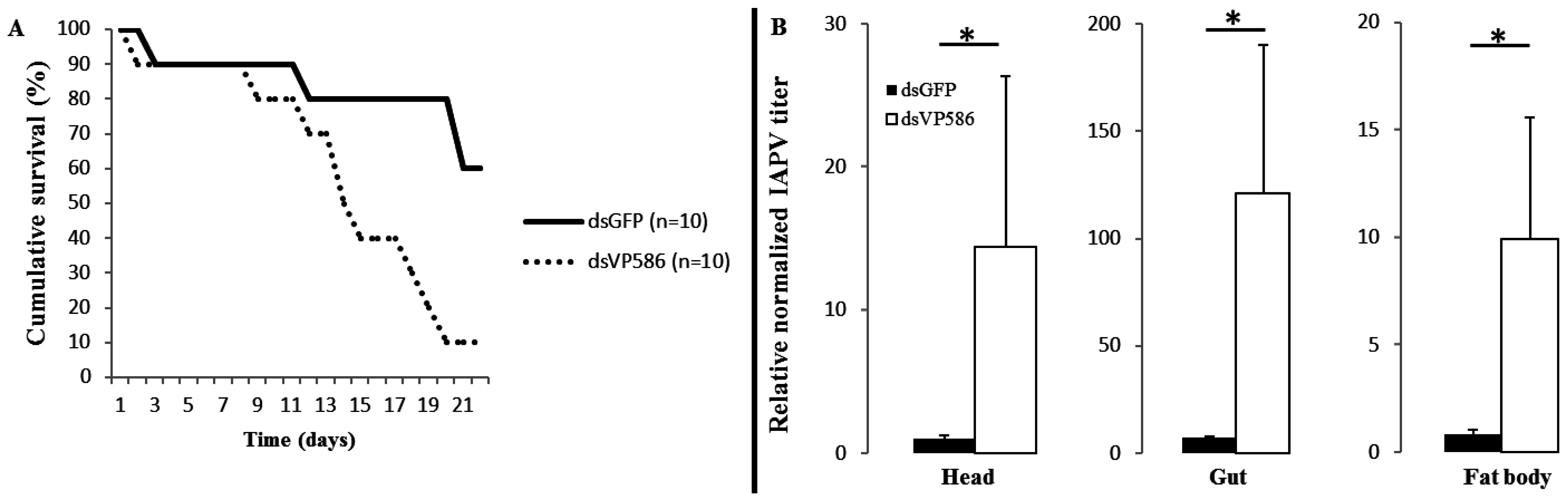
3.2. Effect of Oral Application of dsRNA on IAPV Infectivity, Relative Virus Titer and Bumblebee Survival

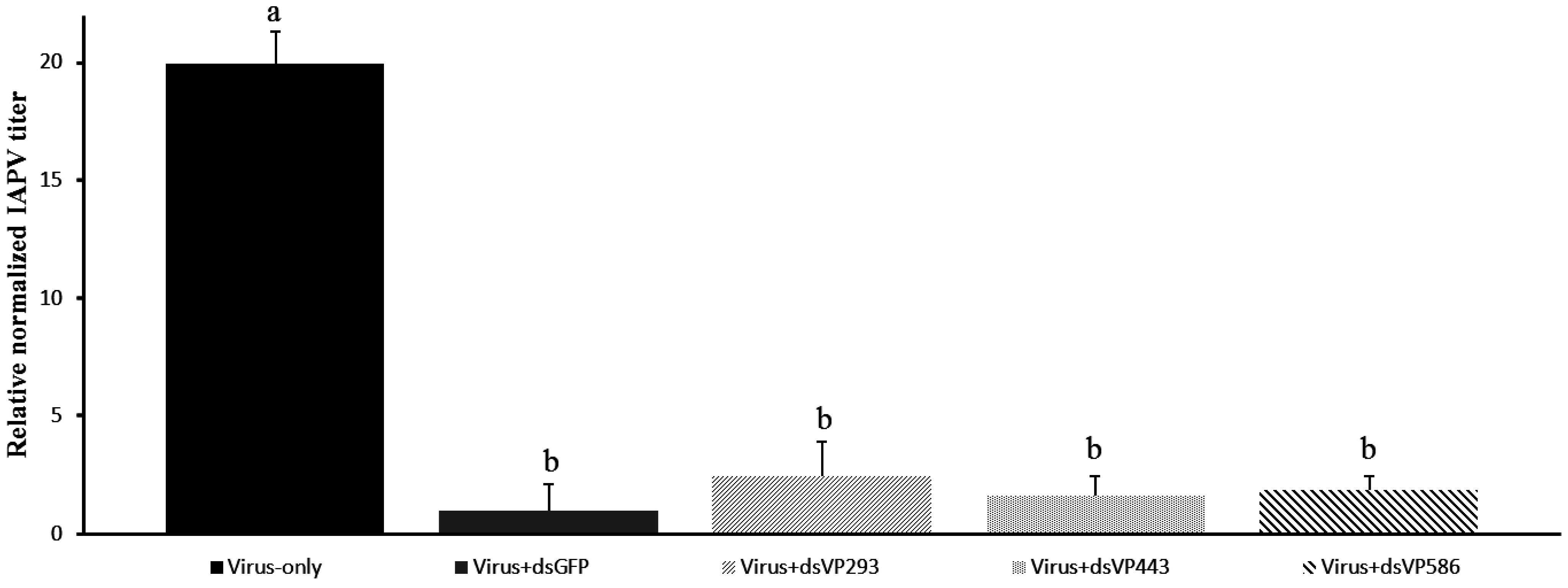
3.3. Confirmation of Lower Relative Virus Titers after Non-Specific and Specific dsRNA Treatment

4. Discussion
4.1. Pathology and Tissue Tropism of IAPV in Bumblebees
4.2. dsRNA As Possible Treatment to Prevent Spillback Principles
4.3. Does Non-Specific dsRNA Works as A PAMP in Bumblebees?
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Velthuis, H.H.W.; van Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef]
- Chen, Y.P.; Pettis, J.S.; Collins, A.; Feldlaufer, M.F. Prevalence and transmission of honeybee viruses. Appl. Environ. Microbiol. 2006, 72, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Ribière, M.; Ball, B.V.; Aubert, M. Natural History And Geographical Distribution Of Honey Bee Viruses. In Virology and the Honey Bee; Aubert, M., Ball, B., Fries, I., Moritz, R., Milani, N., Bernardinelli, I., Eds.; EC Publications: Luxembourg, Luxembourg, 2008; pp. 15–84. [Google Scholar]
- De Miranda, J.R.; Cordoni, G.; Budge, G. The Acute bee paralysis virus-Kashmir bee virus-Israeli acute paralysis virus complex. J. Invertebr. Pathol. 2010, 103, S30–S47. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Levitt, A.L.; Rajotte, E.G.; Holmes, E.C.; Ostiguy, N.; Vanengelsdorp, D.; Lipkin, W.I.; Depamphilis, C.W.; Toth, A.L.; Cox-Foster, D.L. RNA viruses in hymenopteran pollinators: Evidence of inter-taxa virus transmission via pollen and potential impact on non-Apis hymenopteran species. PLoS ONE 2010, 5, e14357. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.P.; Fürst, M.A.; Caspar, J.; Theodorou, P.; Brown, M.J.F.; Paxton, R.J. A sting in the spit: Widespread cross-infection of multiple RNA viruses across wild and managed bees. J. Anim. Ecol. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
- Meeus, I.; Brown, M.J.F.; de Graaf, D.C.; Smagghe, G. Effects of invasive parasites on bumble bee declines. Conserv. Biol. 2011, 25, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Murray, T.E.; Coffey, M.F.; Kehoe, E.; Horgan, F.G. Pathogen prevalence in commercially reared bumble bees and evidence of spillover in conspecific populations. Biol. Conserv. 2013, 159, 269–276. [Google Scholar] [CrossRef]
- Graystock, P.; Goulson, D.; Hughes, W.O.H. The relationship between managed bees and the prevalence of parasites in bumblebees. Peer J. Prepr. 2014, 2, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Hempel, R.; Eckhardt, M.; Goulson, D.; Heinzmann, D.; Lange, C.; Plischuk, S.; Escudero, L.R.; Salathé, R.; Scriven, J.J.; Schmid-Hempel, P. The invasion of southern South America by imported bumblebees and associated parasites. J. Anim. Ecol. 2014, 83, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Arbetman, M.P.; Meeus, I.; Morales, C.L.; Aizen, M.A.; Smagghe, G. Alien parasite hitchhikes to Patagonia on invasive bumblebee. Biol. Invasions 2012, 15, 489–494. [Google Scholar] [CrossRef]
- Maharramov, J.; Meeus, I.; Maebe, K.; Arbetman, M.; Morales, C.; Graystock, P.; Hughes, W.O.H.; Plischuk, S.; Lange, C.E.; de Graaf, D.C.; et al. Genetic variability of the neogregarine Apicystis bombi, an etiological agent of an emergent bumblebee disease. PLoS ONE 2013, 8, e81475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fürst, M.A; McMahon, D.P.; Osborne, J.L.; Paxton, R.J.; Brown, M.J.F. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 2014, 506, 364–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. IAPV, a bee-affecting virus associated with colony collapse disorder can be silenced by dsRNA ingestion. Insect Mol. Biol. 2009, 18, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Hunter, W.; Ellis, J.; Vanengelsdorp, D.; Hayes, J.; Westervelt, D.; Glick, E.; Williams, M.; Sela, I.; Maori, E.; Pettis, J.; et al. Large-scale field application of RNAi technology reducing Israeli acute paralysis virus disease in honey bees (Apis mellifera, hymenoptera: Apidae). PLoS Pathog. 2010, 6, e1001160. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Y.; Yan, X.; Han, R. Prevention of chinese sacbrood virus infection in apis cerana using rna interference. Curr. Microbiol. 2010, 61, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.D.; Eu, Y.J.; Whyard, S.; Currie, R.W. Reduction in deformed wing virus infection in larval and adult honey bees (Apis mellifera L.) by double-stranded RNA ingestion. Insect Mol. Biol. 2012, 21, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Paldi, N.; Glick, E.; Oliva, M.; Zilberberg, Y.; Aubin, L.; Pettis, J.; Chen, Y.; Evans, J.D. Effective gene silencing in a microsporidian parasite associated with honeybee (Apis mellifera) colony declines. Appl. Environ. Microbiol. 2010, 76, 5960–5964. [Google Scholar] [CrossRef] [PubMed]
- Sadd, B.M.; Barribeau, S.M.; Bloch, G.; de Graaf, D.C.; Dearden, P.; Elsik, C.G.; Gadau, J.; Grimmelikhuijzen, C.J.; Hasselmann, M.; Lozier, J.D.; et al. The genomes of two key bumblebee species with primitive eusocial organization. Genome Biol. 2015, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshwal, S.; Mallon, E.B. Antimicrobial peptides play a functional role in bumblebee anti-trypanosome defense. Dev. Comp. Immunol. 2013, 42, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Piot, N.; Meeus, I.; Smagghe, G. Injection of Israeli acute paralysis virus-specific dsRNA reduces virus replication and mortality in Bombus terrestris. Submitted.
- Wang, P.H.; Weng, S.P.; He, J.G. Nucleic acid-induced antiviral immunity in invertebrates: An evolutionary perspective. Dev. Comp. Immunol. 2014, 48, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Flenniken, M.L.; Andino, R. Non-specific dsRNA-mediated antiviral response in the honey bee. PLoS ONE 2013, 8, e77263. [Google Scholar] [CrossRef] [PubMed]
- Robalino, J.; Bartlett, T.C.; Chapman, R.W.; Gross, P.S.; Browdy, C.L.; Warr, G.W. Double-stranded RNA and antiviral immunity in marine shrimp: Inducible host mechanisms and evidence for the evolution of viral counter-responses. Dev. Comp. Immunol. 2007, 31, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Meeus, I.; Mosallanejad, H.; Niu, J.; de Graaf, D.C.; Wäckers, F.; Smagghe, G. Gamma irradiation of pollen and eradication of Israeli acute paralysis virus. J. Invertebr. Pathol. 2014, 121, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Strauss, A.; White, A.; Boots, M. Invading with biological weapons: The importance of disease-mediated invasions. Funct. Ecol. 2012, 26, 1249–1261. [Google Scholar] [CrossRef]
- Meeus, I.; de Miranda, J.R.; de Graaf, D.C.; Wäckers, F.; Smagghe, G. Effect of oral infection with Kashmir bee virus and Israeli acute paralysis virus on bumblebee (Bombus terrestris) reproductive success. J. Invertebr. Pathol. 2014, 121, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Mommaerts, V.; Sterk, G.; Smagghe, G. Hazards and uptake of chitin synthesis inhibitors in bumblebees Bombus terrestris. Pest Manag. Sci. 2006, 62, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Cappelle, K.; de Miranda, J.R.; Smagghe, G.; Meeus, I. Analysis of reference gene stability after Israeli acute paralysis virus infection in bumblebees Bombus terrestris. J. Invertebr. Pathol. 2014, 115, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Lavi, S.; Mozes-Koch, R.; Gantman, Y.; Peretz, Y.; Edelbaum, O.; Tanne, E.; Sela, I. Isolation and characterization of Israeli acute paralysis virus, a dicistrovirus affecting honeybees in Israel: Evidence for diversity due to intra- and inter-species recombination. J. Gen. Virol. 2007, 88, 3428–3438. [Google Scholar] [CrossRef] [PubMed]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Manley, R.; Boots, M.; Wilfert, L. Emerging viral disease risk to pollinating insects: Ecological , evolutionary and anthropogenic factors. J. Appl. Ecol. 2015, 52, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Levitt, A.L.; Singh, R.; Cox-Foster, D.L.; Rajotte, E.; Hoover, K.; Ostiguy, N.; Holmes, E.C. Cross-species transmission of honey bee viruses in associated arthropods. Virus Res. 2013, 176, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Pettis, J.S.; Corona, M.; Chen, W.P.; Li, C.J.; Spivak, M.; Visscher, P.K.; DeGrandi-Hoffman, G.; Boncristiani, H.; Zhao, Y.; et al. Israeli acute paralysis virus: Epidemiology, pathogenesis and implications for honey bee health. PLoS Pathog. 2014, 10, e1004261. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Ribière, M.; Lallemand, P.; Iscache, A.L.; Schurr, F.; Celle, O.; Blanchard, P.; Olivier, V.; Faucon, J.P. Spread of infectious chronic bee paralysis virus by honeybee (Apis mellifera L.) feces. Appl. Environ. Microbiol. 2007, 73, 7711–7716. [Google Scholar] [CrossRef] [PubMed]
- Otterstatter, M.C.; Thomson, J.D. Does pathogen spillover from commercially reared bumble bees threaten wild pollinators? PLoS ONE 2008, 3, e2771. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Rivkin, H.; Slabezki, Y.; Chejanovsky, N. Dynamics of the presence of israeli acute paralysis virus in honey bee colonies with colony collapse disorder. Viruses 2014, 6, 2012–2027. [Google Scholar] [CrossRef] [PubMed]
- Ribière, M.; Drajnudel, P.; Faucon, J.-P. The collapse of bee colonies: The CCD case (“Colony collapse disorder”) and the IAPV virus (Israeli acute paralysis virus). Virology 2008, 12, 319–322. [Google Scholar]
- Colla, S.R.; Otterstatter, M.C.; Gegear, R.J.; Thomson, J.D. Plight of the bumble bee: Pathogen spillover from commercial to wild populations. Biol. Conserv. 2006, 129, 461–467. [Google Scholar] [CrossRef]
- Graystock, P.; Yates, K.; Evison, S.E.F.; Darvill, B.; Goulson, D.; Hughes, W.O.H. The Trojan hives: Pollinator pathogens, imported and distributed in bumblebee colonies. J. Appl. Ecol. 2013, 50, 1207–1215. [Google Scholar] [CrossRef]
- Jones, C.M.; Brown, M.J.F. Parasites and genetic diversity in an invasive bumblebee. J. Anim. Ecol. 2014, 83, 1428–1440. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.M.; Brown, M.J.F.; Oldroyd, B.P. Inbred and outbred honey bees (Apis mellifera) have similar innate immune responses. Insectes Soc. 2012, 60, 97–102. [Google Scholar] [CrossRef]
- Moret, Y.; Schmid-Hempel, P. Survival for immunity: The price of immune system activation for bumblebee workers. Science 2000, 290, 1166–1168. [Google Scholar] [CrossRef] [PubMed]
- Schlüns, H.; Sadd, B.M.; Schmid-Hempel, P.; Crozier, R.H. Infection with the trypanosome Crithidia bombi and expression of immune-related genes in the bumblebee Bombus terrestris. Dev. Comp. Immunol. 2010, 34, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Wilfert, L.; Gadau, J.; Baer, B.; Schmid-Hempel, P. Natural variation in the genetic architecture of a host-parasite interaction in the bumblebee Bombus terrestris. Mol. Ecol. 2007, 16, 1327–1339. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piot, N.; Snoeck, S.; Vanlede, M.; Smagghe, G.; Meeus, I. The Effect of Oral Administration of dsRNA on Viral Replication and Mortality in Bombus terrestris. Viruses 2015, 7, 3172-3185. https://doi.org/10.3390/v7062765
Piot N, Snoeck S, Vanlede M, Smagghe G, Meeus I. The Effect of Oral Administration of dsRNA on Viral Replication and Mortality in Bombus terrestris. Viruses. 2015; 7(6):3172-3185. https://doi.org/10.3390/v7062765
Chicago/Turabian StylePiot, Niels, Simon Snoeck, Maarten Vanlede, Guy Smagghe, and Ivan Meeus. 2015. "The Effect of Oral Administration of dsRNA on Viral Replication and Mortality in Bombus terrestris" Viruses 7, no. 6: 3172-3185. https://doi.org/10.3390/v7062765
APA StylePiot, N., Snoeck, S., Vanlede, M., Smagghe, G., & Meeus, I. (2015). The Effect of Oral Administration of dsRNA on Viral Replication and Mortality in Bombus terrestris. Viruses, 7(6), 3172-3185. https://doi.org/10.3390/v7062765