
The Use of Convalescent Sera in Immune-Electron Microscopy to Detect Non-Suspected/New Viral Agents


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Convalescent Sera
Determination of the Optimal Dilution of Sera
2.2. IAEM Method
2.3. Controls
3. Results and Discussion
3.1. Porcine Rotavirus (PoRV), Porcine Circovirus (PCV2), Porcine Torovirus (PoToV) and Porcine Epidemic Diarrhea Coronavirus (PEDV) in Pigs with Enteritis
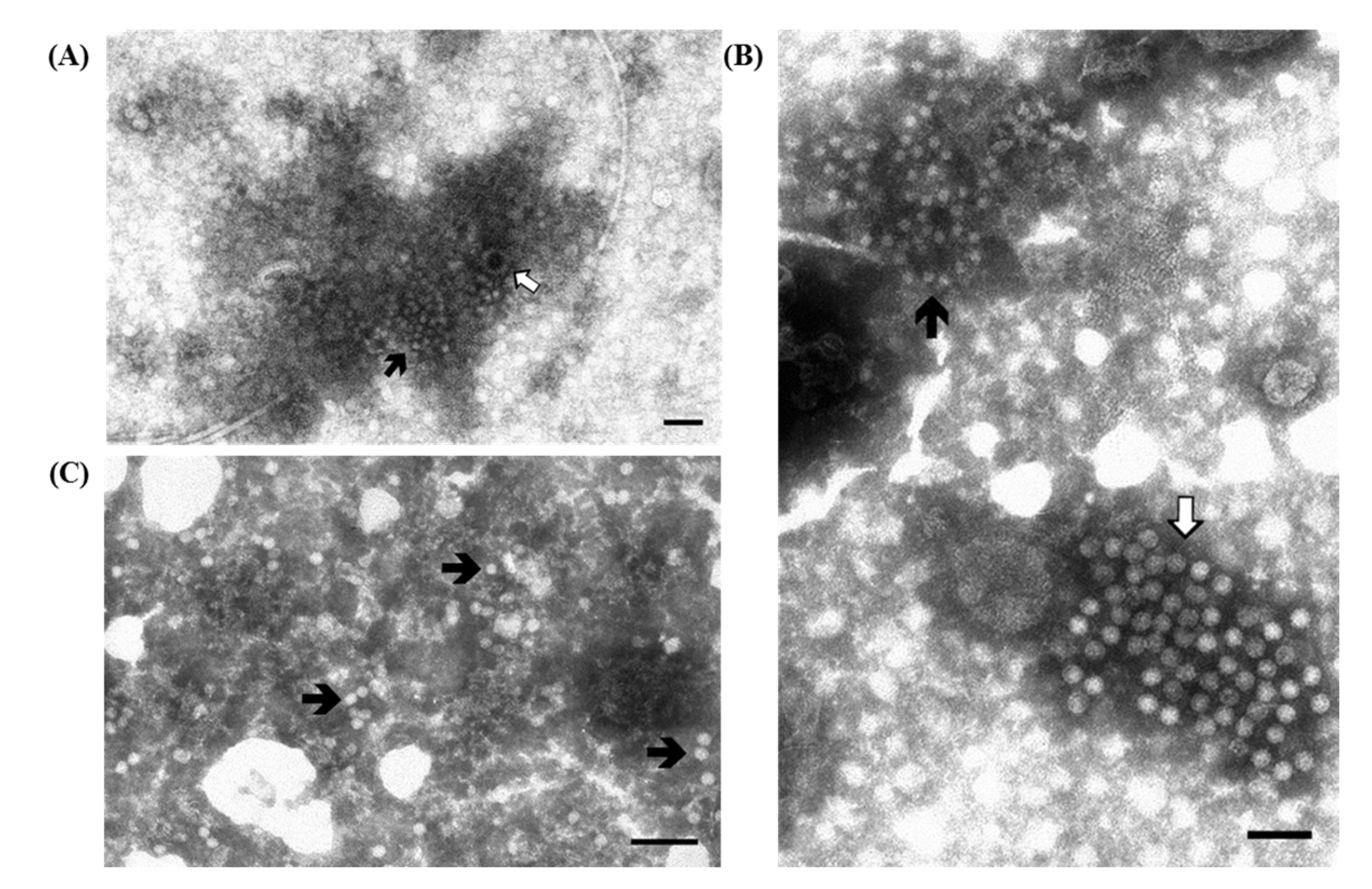
3.1.1. Porcine Rotavirus (PoRV)
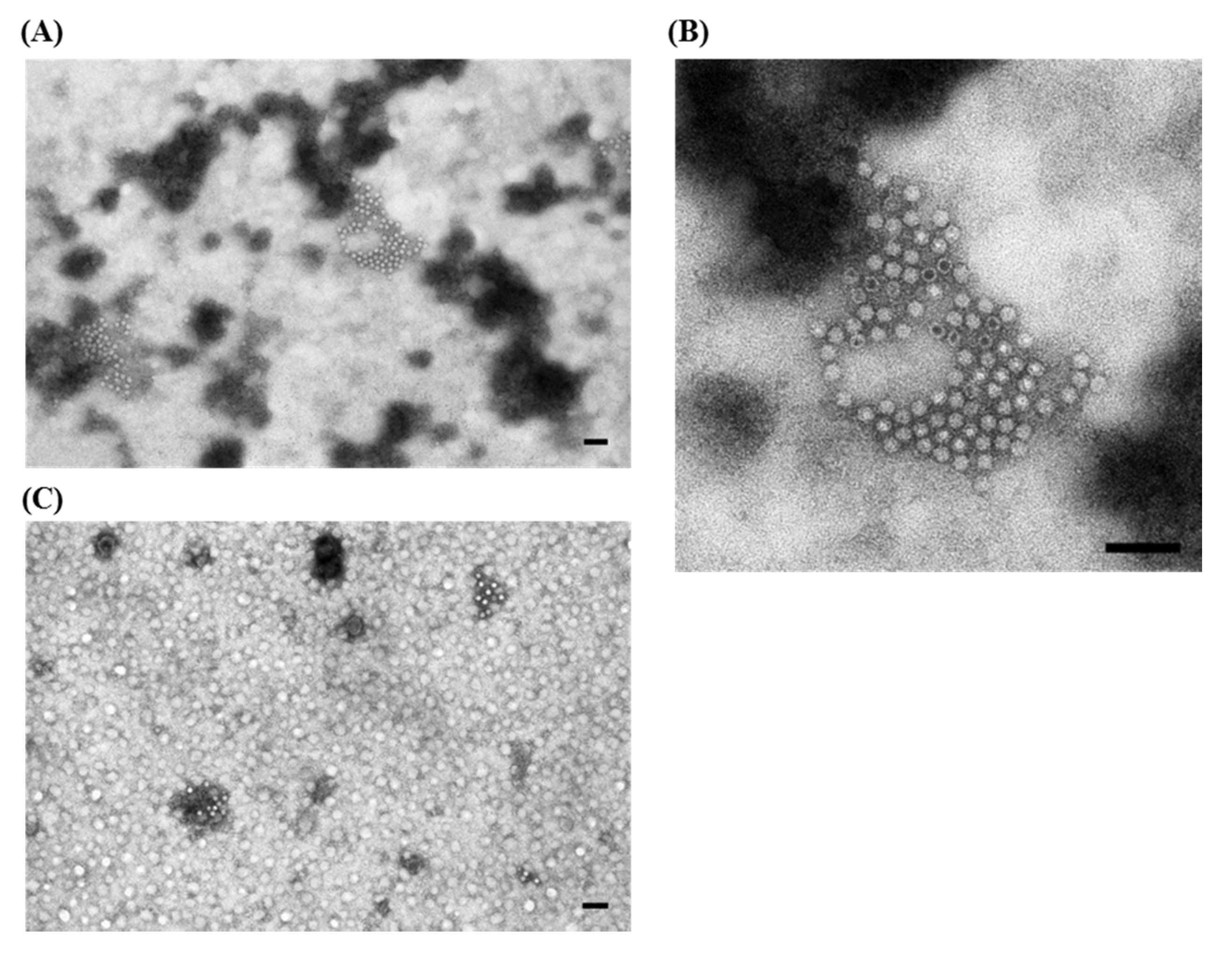
 ) are visible; (D,E) Higher magnifications of (C) showing respectively one group of rotavirus and one group of small round fecal virus/enterovirus-like virus. IAEM preparation with convalescent sera diluted 1:80. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm; (F) IAEM negative control. Dispersed rotavirus particles. TEM Philips CM10, 80 kV. Bar = 100 nm.
) are visible; (D,E) Higher magnifications of (C) showing respectively one group of rotavirus and one group of small round fecal virus/enterovirus-like virus. IAEM preparation with convalescent sera diluted 1:80. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm; (F) IAEM negative control. Dispersed rotavirus particles. TEM Philips CM10, 80 kV. Bar = 100 nm.
) are visible; (D,E) Higher magnifications of (C) showing respectively one group of rotavirus and one group of small round fecal virus/enterovirus-like virus. IAEM preparation with convalescent sera diluted 1:80. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm; (F) IAEM negative control. Dispersed rotavirus particles. TEM Philips CM10, 80 kV. Bar = 100 nm.
) are visible; (D,E) Higher magnifications of (C) showing respectively one group of rotavirus and one group of small round fecal virus/enterovirus-like virus. IAEM preparation with convalescent sera diluted 1:80. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm; (F) IAEM negative control. Dispersed rotavirus particles. TEM Philips CM10, 80 kV. Bar = 100 nm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | Total | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. exams | 716 | 788 | 622 | 384 | 746 | 333 | 3589 | |||||||
| No. negative | 407 | 56.8 | 399 | 50.6 | 412 | 66.2 | 288 | 75.0 | 489 | 65.5 | 220 | 66.1 | 2215 | 61.7 |
| No. positive | 309 | 43.2 | 389 | 49.4 | 210 | 33.8 | 96 | 25.0 | 257 | 34,5 | 113 | 33.9 | 1374 | 38.3 |
| Adenovirus | 3 | 0.4 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 0.3 | 4 | 0.1 |
| Calicivirus | 3 | 0.4 | 2 | 0.3 | 6 | 1.0 | 0 | 0.0 | 5 | 0.7 | 3 | 0.9 | 19 | 0.5 |
| Coronavirus | 152 | 21.2 | 224 | 28.4 | 71 | 11.4 | 38 | 9.9 | 152 | 20.4 | 33 | 9.9 | 670 | 18.7 |
| Circovirus | 54 | 7.5 | 107 | 13.6 | 0 | 0.0 | 0 | 0.0 | 2 | 0.3 | 3 | 0.9 | 166 | 4.6 |
| Enterovirus | 16 | 2.2 | 2 | 0.3 | 0 | 0.0 | 0 | 0.0 | 15 | 2.0 | 19 | 5.7 | 52 | 1.4 |
| Parvovirus | 2 | 0.3 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 2 | 0.1 |
| Rotavirus | 59 | 8.2 | 99 | 12.6 | 123 | 19.8 | 40 | 10.4 | 56 | 7.5 | 55 | 16.5 | 432 | 12.0 |
| Torovirus | 4 | 0.6 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 4 | 0.1 |
| Not classified | 155 | 21.6 | 245 | 31.1 | 43 | 6.9 | 31 | 8.1 | 32 | 4.3 | 12 | 3.6 | 518 | 14.4 |
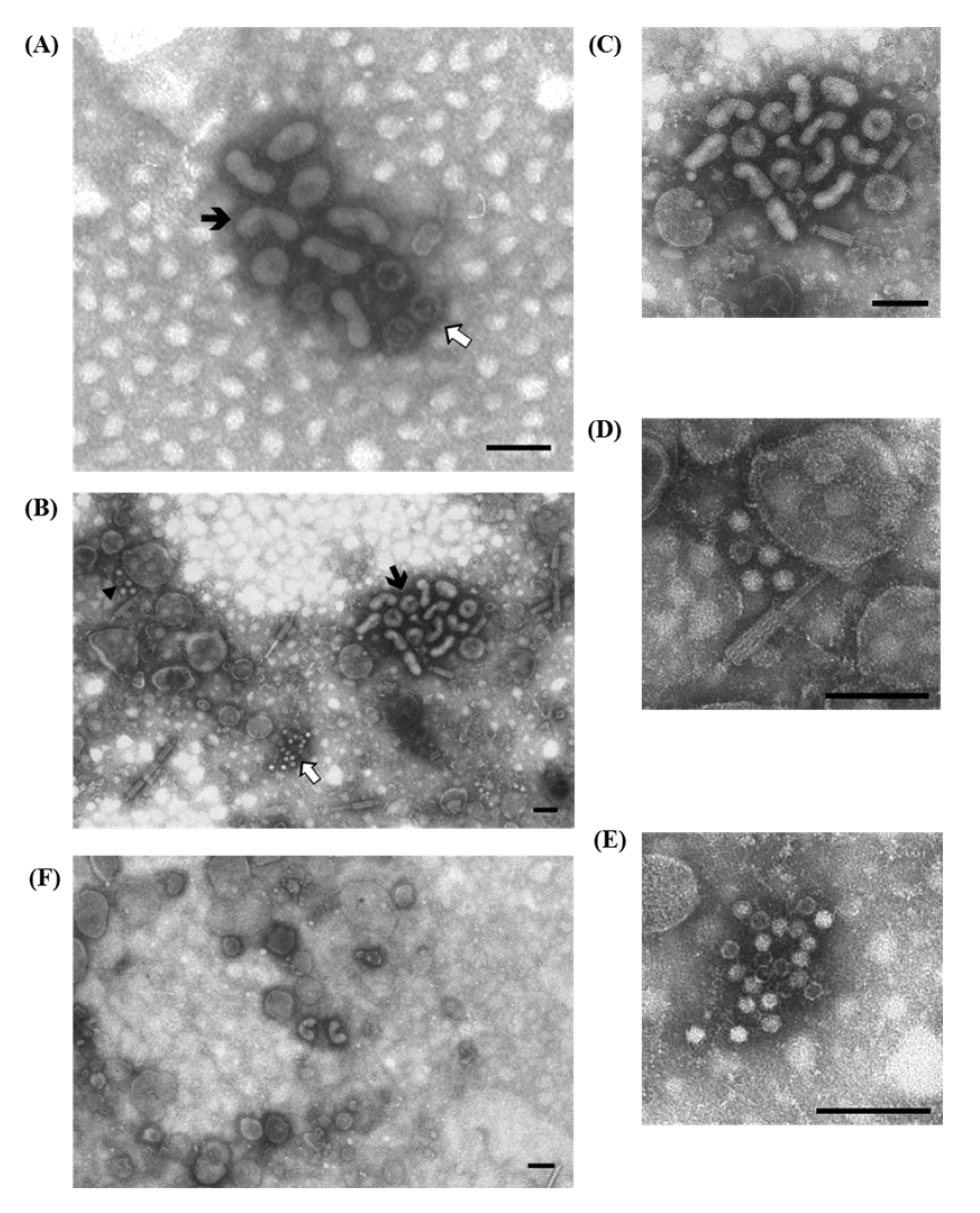
3.1.2. Porcine Circovirus Type 2 (PCV2)
) are grouped together with (A) a few partially destroyed and empty rotavirus particles (⇧) and (B) a large group of enterovirus-like particles (⇧). Immuno-aggregation electron microscopy preparation with convalescent sera diluted 1:40; (C) IAEM negative control. Several circovirus particles ( ) are visible, singularly distributed or in small groups. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm.
) are grouped together with (A) a few partially destroyed and empty rotavirus particles (⇧) and (B) a large group of enterovirus-like particles (⇧). Immuno-aggregation electron microscopy preparation with convalescent sera diluted 1:40; (C) IAEM negative control. Several circovirus particles ( ) are visible, singularly distributed or in small groups. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm.
| Viral Agent | No. PCV2 Pos. | % |
|---|---|---|
| Coronavirus/PED | 27 | 60.0 |
| Enterovirus-Like | 26 | 57.8 |
| Rotavirus | 4 | 8.9 |
| Others (parvovirus, calicivirus, adenovirus) | 7 | 15.6 |
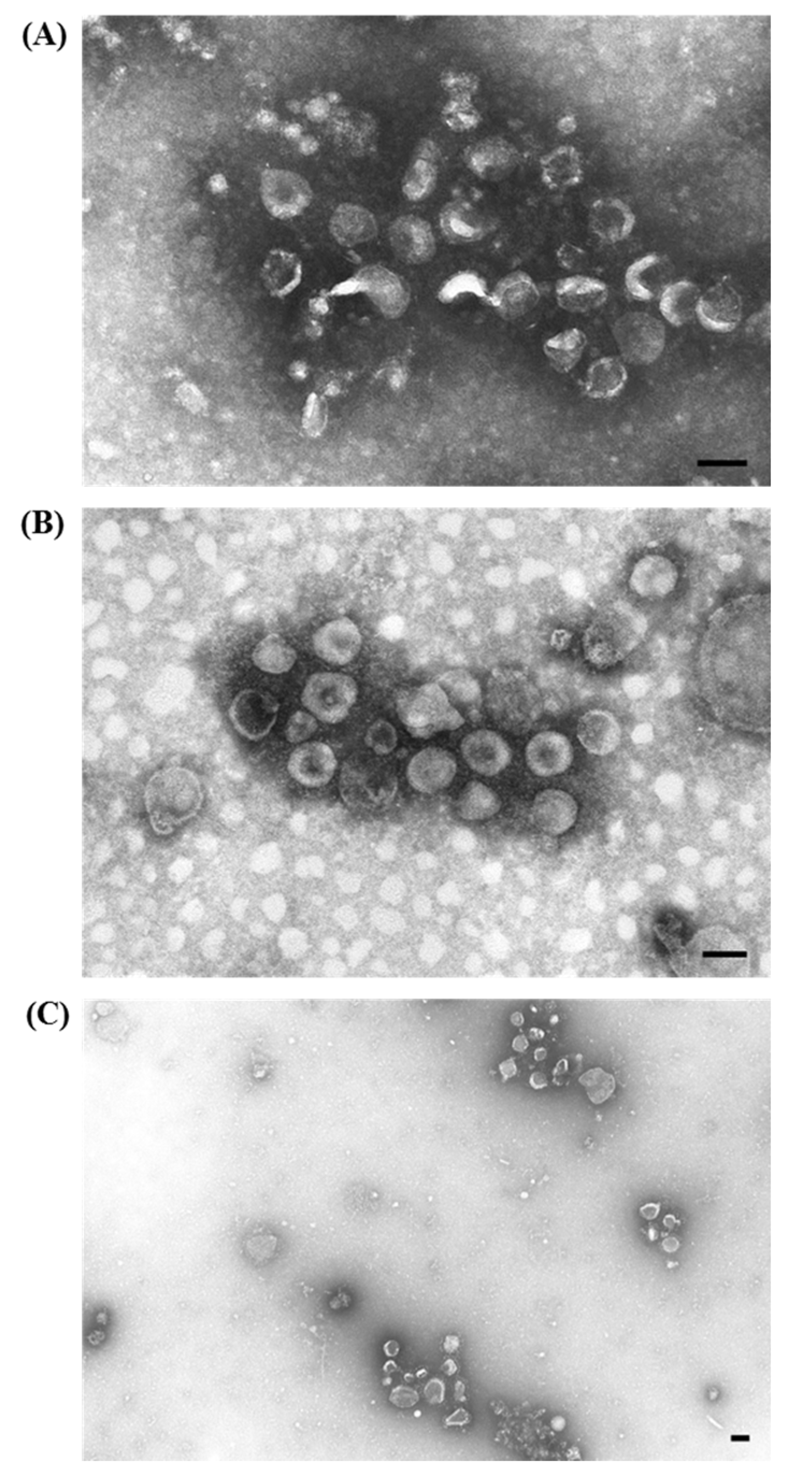
3.1.3. Porcine Torovirus (PToV)
3.1.4 Porcine Epidemic Diarrhea Virus (PEDV)
) are grouped together with (A) rotavirus particles (⇧) and (B) groups of either enterovirus-like (▲) or circovirus (⇧) particles; (C) Higher magnification of (B) showing the aggregate of pleomorphic torovirus particles with a fringed surface in which short peplomers are immersed in an halo of antibodies; (D) Higher magnification of the small aggregate in the upper left of (B) composed by four full and one empty enterovirus-like particles close to a rod shaped phage-tail; (E) Higher magnification of (B) showing the group of empty and full circovirus particles; (F) IAEM negative control: two kidney-shaped torovirus particles are visible in the middle together with several isolated fringed particles. Immuno-aggregation electron microscopy preparation with convalescent sera diluted 1:80. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm.
) are grouped together with (A) rotavirus particles (⇧) and (B) groups of either enterovirus-like (▲) or circovirus (⇧) particles; (C) Higher magnification of (B) showing the aggregate of pleomorphic torovirus particles with a fringed surface in which short peplomers are immersed in an halo of antibodies; (D) Higher magnification of the small aggregate in the upper left of (B) composed by four full and one empty enterovirus-like particles close to a rod shaped phage-tail; (E) Higher magnification of (B) showing the group of empty and full circovirus particles; (F) IAEM negative control: two kidney-shaped torovirus particles are visible in the middle together with several isolated fringed particles. Immuno-aggregation electron microscopy preparation with convalescent sera diluted 1:80. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm.

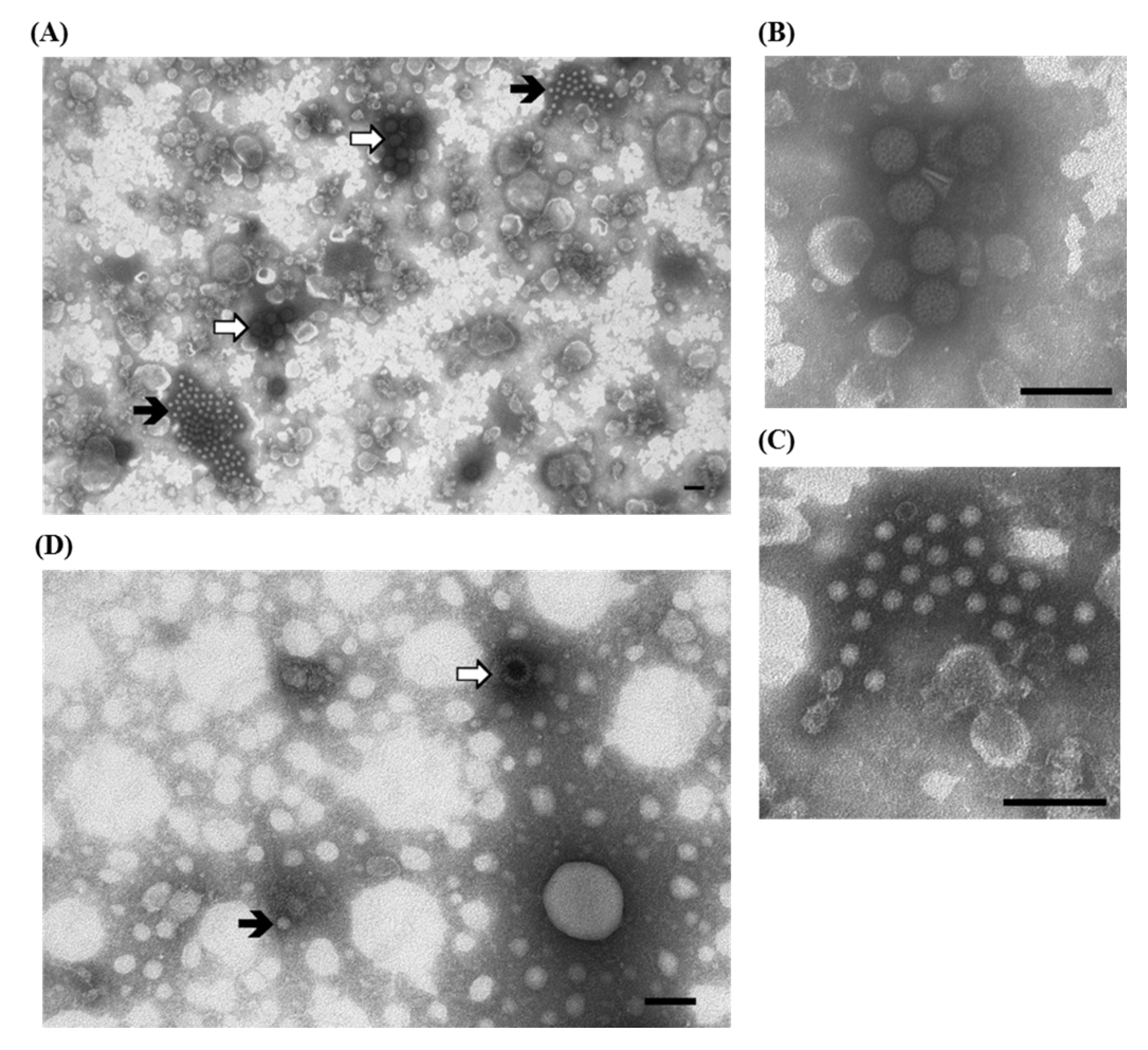
3.2. Rotavirus and Astrovirus in Young Turkeys with Enteritis
), surrounded by a fuzzy halo of antibodies are visible; (B) Higher magnification of rotavirus particles: the presence of a halo of antibodies does not mask the typical single—and double-shelled morphology of rotaviruses; (C) Higher magnification of astrovirus particles: the surface of aggregated virions, even if not always distinctively showing the 5–6 star-shaped pattern, do not have a completely homogeneous or featureless surface, which is highly indicative of astroviruses; (D) IAEM negative control: one isolated rotavirus ( ) and one astrovirus-like particle (⇧) are visible. Immuno-aggregation electron microscopy (IAEM) preparation with convalescent sera diluted 1:40. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm.
), surrounded by a fuzzy halo of antibodies are visible; (B) Higher magnification of rotavirus particles: the presence of a halo of antibodies does not mask the typical single—and double-shelled morphology of rotaviruses; (C) Higher magnification of astrovirus particles: the surface of aggregated virions, even if not always distinctively showing the 5–6 star-shaped pattern, do not have a completely homogeneous or featureless surface, which is highly indicative of astroviruses; (D) IAEM negative control: one isolated rotavirus ( ) and one astrovirus-like particle (⇧) are visible. Immuno-aggregation electron microscopy (IAEM) preparation with convalescent sera diluted 1:40. Negative staining (2% sodium phosphotungstate). TEM Philips CM10, 80 kV. Bar = 100 nm.
| Year | No. | Negative | Rotavirus | Astrovirus | Enterovirus-Like Virus | Parvovirus-Like Virus | Adenovirus | RSVLP | Coronavirus | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | ||
| 1994 | 14 | 12 | 85.7 | 1 | 7.1 | 0 | 0.0 | 1 | 7.1 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 1995 | 11 | 1 | 9.1 | 3 | 27.3 | 4 | 36.4 | 3 | 27.3 | 2 | 18.2 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 1996 | 7 | 1 | 14.3 | 0 | 0.0 | 3 | 42.9 | 3 | 42.9 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 1997 | 26 | 17 | 65.4 | 7 | 26.9 | 2 | 7.7 | 2 | 7.7 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 1998 | 49 | 28 | 57.1 | 10 | 20.4 | 16 | 32.7 | 3 | 6.1 | 1 | 2.0 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 1999 | 48 | 23 | 47.9 | 11 | 22.9 | 13 | 27.1 | 2 | 4.2 | 0 | 0.0 | 2 | 4.2 | 0 | 0.0 | 4 | 8.3 |
| 2000 | 41 | 19 | 46.3 | 14 | 34.1 | 10 | 24.4 | 0 | 0.0 | 4 | 9.8 | 1 | 2.4 | 2 | 4.9 | 1 | 2.4 |
| 2001 | 64 | 26 | 40.6 | 22 | 34.4 | 14 | 21.9 | 12 | 18.8 | 2 | 3.1 | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 |
| 2002 | 77 | 46 | 59.7 | 11 | 14.3 | 17 | 22.1 | 0 | 0.0 | 6 | 7.8 | 2 | 2.6 | 1 | 1.3 | 1 | 1.3 |
| 2003 | 118 | 50 | 42.4 | 43 | 36.4 | 32 | 27.1 | 6 | 5.1 | 6 | 5.1 | 3 | 2.5 | 0 | 0.0 | 4 | 3.4 |
| 2004 | 26 | 11 | 42.3 | 6 | 23.1 | 13 | 50.0 | 1 | 3.8 | 0 | 0.0 | 1 | 3.8 | 0 | 0.0 | 0 | 0.0 |
| Total | 481 | 234 | 48.6 | 128 | 26.6 | 124 | 25.8 | 33 | 6.9 | 21 | 4.4 | 9 | 1.9 | 3 | 0.6 | 10 | 2.1 |
| Type of Association | 1994 | 1995 | 1996 | 1997 | 1998 | 1999 | 2000 | 2001 | 2002 | 2003 | 2004 | Tot. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rotavirus + Astrovirus | 1 | 7 | 7 | 6 | 10 | 4 | 17 | 4 | 56 | |||
| Rotavirus + Astrovirus + Enterovirus-like | 1 | 1 | ||||||||||
| Rotavirus + Enterovirus-like | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | ||||
| Rotavirus + Parvovirus-like | 1 | 1 | 2 | 4 | ||||||||
| Rotavirus + Coronavirus | 1 | 1 | ||||||||||
| Astrovirus + Parvovirus-like | 1 | 1 | 2 | |||||||||
| Astrovirus + Adenovirus | 1 | 1 | 2 | |||||||||
| Astrovirus + Coronavirus | 1 | 1 | ||||||||||
| Enterovirus-like + Parvovirus-like | 1 | 1 | 2 | |||||||||
| Adenovirus + Parvovirus-like | 1 | 1 | 2 | |||||||||
| Total | 0 | 2 | 0 | 2 | 9 | 7 | 9 | 12 | 7 | 25 | 6 | 79 |
3.3. Parvovirus-Like Particles in Pheasants

3.4. Lagoviruses: Rabbit Hemorrhagic Disease Virus (RHDV) of Rabbits and European Brown Hare Syndrome Virus (EBHSV) of Hares



4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gentile, M.; Gelderblom, H.R. Rapid viral diagnosis: Role of electron microscopy. New Microbiol. 2005, 28, 1–12. [Google Scholar] [PubMed]
- Biel, S.; Gelderblom, H.R. Diagnostic Electron Microscopy is still timely and rewarding method. J. Clin. Virol. 1999, 13, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, H.R.; Renz, H.; Özel, M. Negative staining in diagnostic virology. Micron Microsc. Acta 1991, 22, 435–447. [Google Scholar] [CrossRef]
- Hazelton, P.R.; Gelderblom, H.R. Electron microscopy for rapid diagnosis of infectious agents in emergent situations. Emerg. Infect. Dis. 2003, 9, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Hopley, J.F.A.; Doane, F.W. Development of a sensitive Protein A-Gold immunoelectronmicroscopy method for detecting viral antigen in fluid specimens. J. Virol. Methods 1985, 12, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsberg, E. Demonstration of calicivirus in human faeces by immunosorbent and immunogold-labelling electron microscopy methods. J. Virol. Methods 1986, 14, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Spencer, R.R. The Zone Phenomenon in Agglutination Tests. J. Infect. Dis. 1930, 46, 138–147. [Google Scholar] [CrossRef]
- Estes, M.K.; Kapikian, A.Z. Rotaviruses. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, M.A., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott, Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 1917–1974. [Google Scholar]
- Saif, L.J.; Bohl, E.H.; Theil, K.W.; Cross, R.F.; House, J.A. Rotavirus-like, calicivirus-like, and 23-nm virus-like particles associated with diarrhea in young pigs. J. Clin. Microbiol. 1980, 12, 105–111. [Google Scholar] [PubMed]
- Bridger, J.C. Detection by electron microscopy of caliciviruses, astroviruses and rotavirus-like particles in the faeces of piglets with diarrhea. Vet. Rec. 1980, 107, 532–533. [Google Scholar] [PubMed]
- Martella, V.; Bányai, K.; Lorusso, E.; Bellacicco, A.L.; Decaro, N.; Camero, M.; Bozzo, G.; Moschidou, P.; Arista, S.; Pezzotti, G.; et al. Prevalence of group C rotaviruses in weaning and post-weaning pigs with enteritis. Vet. Microbiol. 2007, 123, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Lorusso, E.; Banyai, K.; Decaro, N.; Corrente, M.; Elia, G.; Cavalli, A.; Radogna, A.; Costantini, V.; Saif, L.J.; et al. Identification of a porcine calicivirus related genetically to human sapoviruses. J. Clin. Microbiol. 2008, 46, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Bridger, J.C. Novel rotaviruses in animals and men. In Novel Diarrhoea Viruses; Bock, G., Whelan, J., Eds.; John Wiley & Sons LTD: Chichester, UK, 1987; CIBA Foundation Symposium 128, London; 15–17 July 1986; pp. 5–15.
- Lavazza, A.; Tittarelli, C.; Cerioli, M.; Luppi, A.; Alborali, G.; Cordioli, P. Electronmicroscopic detection of viral agents in diarrhoeic pigs during the period 2002–2007 in Northern Italy. In Proceedings of the 20th IPVS International Pig Veterinary Science Congress, Waterkloof, Durban, South Africa, 22–26 June 2008; p. 180.
- Sozzi, E.; Barbieri, I.; Lavazza, A.; Lelli, D.; Moreno, A.; Canelli, E.; Bugnetti, M.; Cordioli, P. Molecular Characterization and Phylogenetic Analysis of VP1 of Porcine Enteric Picornaviruses Isolates in Italy. Transbound. Emerg. Dis. 2010, 57, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Bányai, K.; Ciarlet, M.; Iturriza-Gómara, M.; Lorusso, E.; De Grazia, S.; Arista, S.; Decaro, N.; Elia, G.; Cavalli, A.; et al. Relationships among porcine and human Pp[6] rotaviruses: Evidence that the different human P[6] lineages have originated from multiple interspecies transmission events. Virology 2006, 344, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Ciarlet, M.; Banyai, K.; Lorusso, E.; Arista, S.; Lavazza, A.; Pezzotti, G.; Decaro, N.; Cavalli, A.; Lucente, M.S.; et al. Identification of Group A Porcine Rotavirus Strains Bearing a Novel VP4 (P) Genotype in Italian Swine Herds. J. Clin. Microbiol. 2007, 45, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Cerioli, M.; Bozzoni, G.; Lavazza, A. Identificazione di circovirus suino tipo 2 in associazione ad altri agenti patogeni. In Società Italiana di Patologia ed Allevamento dei Suini (SIPAS), Proceedings of the 30th Annual Meeting, “Atti della Società Italiana di Patologia ed Allevamento dei Suini”, Salsomaggiore Terme, Parma, Italia, 25–26 March 2004; pp. 493–500.
- Sala, G.; Rigola, S.; Alborali, G.L.; Brocchi, E.; Cordioli, P. Development of monoclonal antibodies based ELISAS for the detection of antibodies against porcine Circovirus type 1 and type 2. In Veterinary Virology in the New Millennium, Proceedings of the 5th International Congress of the European Society for Veterinary Virology, Brescia, Italy, 27–30 August 2000; Brocchi, E., Lavazza, A., Eds.; Istituto Zooprofilattico Sperimentale della Lombardia e dell’ Emilia Romagna e Fondazione Iniziative Zooprofilattiche e Zootecniche: Brescia, Italy, 2000; pp. 253–254. [Google Scholar]
- Cerioli, M.; Alborali, G.L.; Salogni, C.; Zanoni, M.G.; Pacciarini, M.L.; Lavazza, A. Use of Immunoelectronmicroscopy in Comparison with Immunofluorescence and PCR for the Detection of PMCV-2 from Piglets Clinically Affected by PMWS. In “PRRS, Swine Influenza”, Proceedings of the 4th International Symposium on Emerging and Re-emerging Pig Disease, Rome, Italy, 29 June–2 July 2003; Martelli, P., Cavirani, S., Lavazza, A., Eds.; Faculty of Veterinary medicine: Parma, Italy, 2003; pp. 224–225. [Google Scholar]
- Lavazza, A.; Candotti, P.; Perini, S.; Vezzali, L. Electron Microscopic Identification of Torovirus-like particles in post-weaned piglets with enteritis. In Proceedings of the 14th International Pig Veterinary Society, Bologna, Italy, 7–10 July 1996; Vignola, G., Monetti, P.G., Eds.; Scientific Committee of the 14th IPVS Congress: Bologna, Italy, 1996; p. 131. [Google Scholar]
- Smits, S.L.; Lavazza, A.; Matiz, K.; Horzinek, M.C.; Koopmans, M.P.; De-Groot, R.J. Phylogenetic and evolutionary relationships among torovirus field variants: Evidence for multiple intertypic recombination events. J. Virol. 2003, 77, 9567–9577. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, J.; Jimenez, M.; Luque, J.; Rejas, M.T.; Lavazza, A.; Rodriguez, D. Molecular characterization of a new PToV strain. Evolutionary implications. Virus Res. 2009, 143, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Martelli, P.; Lavazza, A.; Nigrelli, A.D.; Merialdi, G.; Alborali, L.G.; Pensaert, M.B. An epidemic of diarrhoea caused by Porcine Epidemic Diarrhoea virus in Italy. Vet. Rec. 2008, 162, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Sozzi, E.; Luppi, A.; Lelli, D.; Moreno Martin, A.; Canelli, E.; Brocchi, E.; Lavazza, A.; Cordioli, P. Comparison of enzyme-linked immunosorbent assay and RT-PCR for the detection of porcine epidemic diarrhoea virus. Res. Vet. Sci. 2010, 88, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Sozzi, E.; Alborali, L.G.; Luppi, A.; Lelli, D.; Moreno, A.; Cordioli, P.; Brocchi, E. ELISA con anticorpi monoclonali per la diagnostica sierologica della diarrea epidemica suina (PED). In Società Italiana di Patologia ed Allevamento dei Suini (SIPAS), Proceedings of the 33th Annual Meeting, “Atti della Società Italiana di Patologia ed Allevamento dei Suini”, Modena, Italy, 29–30 March 2007; pp. 415–420.
- Kim, S.Y.; Song, D.S.; Park, B.K. Differential detection of transmissible gastroenteritis virus and porcine epidemic diarrhoea virus by duplex RT-PCR. J. Vet. Diagn. Investig. 2001, 13, 516–520. [Google Scholar] [CrossRef]
- Sozzi, E.; Papetti, A.; Lelli, D.; Boniotti, B.; Moreno, A.; Brocchi, E.; Alborali, L.; Lavazza, A.; Cordioli, P. Diagnosis and investigations on PED in Northern Italy. In Proceedings of the 8th Annual Meeting of EPIZONE, “Primed for tomorrow”, Copenhagen, Denmark, 23–25 September 2014; p. 1.
- Estes, M.K.; Palmer, E.L.; Obijeski, J.F. Rotaviruses: A review. Curr. Top. Microbiol. 1983, 105, 123–184. [Google Scholar]
- McNulty, M.S.; Todd, D.; Allan, G.M.; McFerran, J.B.; Greene, J.A. Epidemiology of rotavirus infection in broiler chickens: Recognition of four serogroups. Arch. Virol. 1984, 81, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Saif, Y.M.; Theil, K.W. Enteric viruses in diarrhoeic turkey poults. Avian Dis. 1985, 29, 798–811. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.L.; Saif, Y.M.; Theil, K.W. Enteric Viral Infections of Turkey Poults: Incidence of Infection. Avian Dis. 1987, 31, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Murgia, M.V.; Battilani, M.; Cecchinato, M.; Catelli, E.; Crudi, M.; Vinco, L.J.; Cerioli, M.; Tosi, G.; Lavazza, A. Preliminary data on the molecular characterization of rotavirus detected in diarrhoeic turkey poults in Italy. In Proceedings of the 5th International Symposium on Turkey Diseases, Berlin, Germany, 16–19 June 2004; Hafez, H.M., Pomeroy, B., Eds.; Deutsche Veterinärmedizinische Gesellschaft: Giessen, Germany, 2004; pp. 211–219. [Google Scholar]
- Doane, F.W.; Anderson, N. Electron Microscopy in Diagnostic Virology: A Practical Guide and Atlas; Cambridge University Press: Cambridge, UK, 1987; pp. 100–102. [Google Scholar]
- Palmer, E.L.; Martin, M.L. Electron Microscopy in Viral Diagnosis; CRC Press Inc.: Boca Raton, FL, USA, 1988; p. 55. [Google Scholar]
- Lavazza, A.; Pascucci, S.; Gelmetti, D. Rod-shaped virus-like particles in intestinal contents of three avian species. Vet. Rec. 1990, 126, 581. [Google Scholar] [PubMed]
- Canelli, E.; Cordioli, P.; Barbieri, I.; Catella, A.; Pennelli, D.; Ceruti, R.; Moreno, A.; Lavazza, A. Astroviruses as causative agents of poultry enteritis: Genetic characterization and longitudinal studies on field conditions. Avian Dis. 2012, 56, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Otto, P.H.; Ahmed, M.U.; Hotzel, H.; Machnowska, P.; Reetz, J.; Roth, B.; Trojnar, E.; Johne, R. Detection of avian rotaviruses of groups A, D, F and G in diseased chickens and turkeys from Europe and Bangladesh. Vet. Microbiol. 2012, 156, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Gelmetti, D.; Fabbi, M.; Sironi, G.; Grilli, G.; Lavazza, A. Identification of parvovirus-like particles associated with three outbreaks of mortality in young pheasants (Phasianus colchicus). J. Vet. Diagn. Investig. 1996, 8, 108–112. [Google Scholar] [CrossRef]
- Todd, D.; Westob, J.; Ball, N.W.; Borghmans, B.J.; Gelmini, L.; Lavazza, A. Nucleotide sequence based identification of a novel circovirus of canaries. Avian Pathol. 2001, 30, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, J.; van der Loo, W.; Le Pendu, J.; Esteves, J.P. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, e43. [Google Scholar] [CrossRef]
- Marcato, P.S.; Benazzi, C.; Vecchi, G.; Galeotti, M.; Della Salda, L.; Sarli, G.; Lucidi, P. Clinical and pathological features of viral haemorrhagic disease of rabbits and the European brown hare syndrome. Rev. Sci. Tech. OIE 1991, 10, 371–392. [Google Scholar]
- Lavazza, A.; Vecchi, G. Osservazioni su alcuni episodi di mortalità nella lepre: Evidenziazione al microscopio elettronico di una particella virale: Nota preliminare. Sel. Vet. 1989, 30, 461–468. [Google Scholar]
- Capucci, L.; Scicluna, M.T.; Lavazza, A.; Brocchi, E. Purificazione e caratterizzazione dell’ agente eziologico della malattia emorragica virale del coniglio. Sel. Vet. 1990, 31, 301–312. [Google Scholar]
- Capucci, L.; Scicluna, M.T.; Lavazza, A. Diagnosis of viral haemorrhagic disease of rabbits and the European brown hare syndrome. Rev. Sci. Tech. OIE 1991, 10, 347–370. [Google Scholar]
- Poli, A.; Nigro, M.; Gallazzi, D.; Sironi, G.; Lavazza, A.; Gelmetti, D. Acute hepatosis in the European Brown Hare (Lepus europaeus) in Italy. J. Wildl. Dis. 1991, 27, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Lavazza, A.; Capucci, L. How Many Caliciviruses are there in Rabbits? A Review on RHDV and Correlated Viruses. In Lagomorph Biology; Alves, P.C., Ferrand, N., Hacklaender, K., Eds.; Springer: Berlin, Germany, 2007; pp. 263–278. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavazza, A.; Tittarelli, C.; Cerioli, M. The Use of Convalescent Sera in Immune-Electron Microscopy to Detect Non-Suspected/New Viral Agents. Viruses 2015, 7, 2683-2703. https://doi.org/10.3390/v7052683
Lavazza A, Tittarelli C, Cerioli M. The Use of Convalescent Sera in Immune-Electron Microscopy to Detect Non-Suspected/New Viral Agents. Viruses. 2015; 7(5):2683-2703. https://doi.org/10.3390/v7052683
Chicago/Turabian StyleLavazza, Antonio, Cristiana Tittarelli, and Monica Cerioli. 2015. "The Use of Convalescent Sera in Immune-Electron Microscopy to Detect Non-Suspected/New Viral Agents" Viruses 7, no. 5: 2683-2703. https://doi.org/10.3390/v7052683
APA StyleLavazza, A., Tittarelli, C., & Cerioli, M. (2015). The Use of Convalescent Sera in Immune-Electron Microscopy to Detect Non-Suspected/New Viral Agents. Viruses, 7(5), 2683-2703. https://doi.org/10.3390/v7052683