
Characterisation of Structural Proteins from Chronic Bee Paralysis Virus (CBPV) Using Mass Spectrometry

Abstract
:1. Introduction

2. Materials and Methods
2.1. Virus Purification
2.2. In-Gel Digestion for Protein Identification
2.3. Nano-HPLC MALDI-TOF/TOF Analysis (Peptide Separation and Fractionation and Mass Spectrometry)
2.4. Western Blot Analysis
2.5. Computer-Assisted Sequence Analysis
3. Results
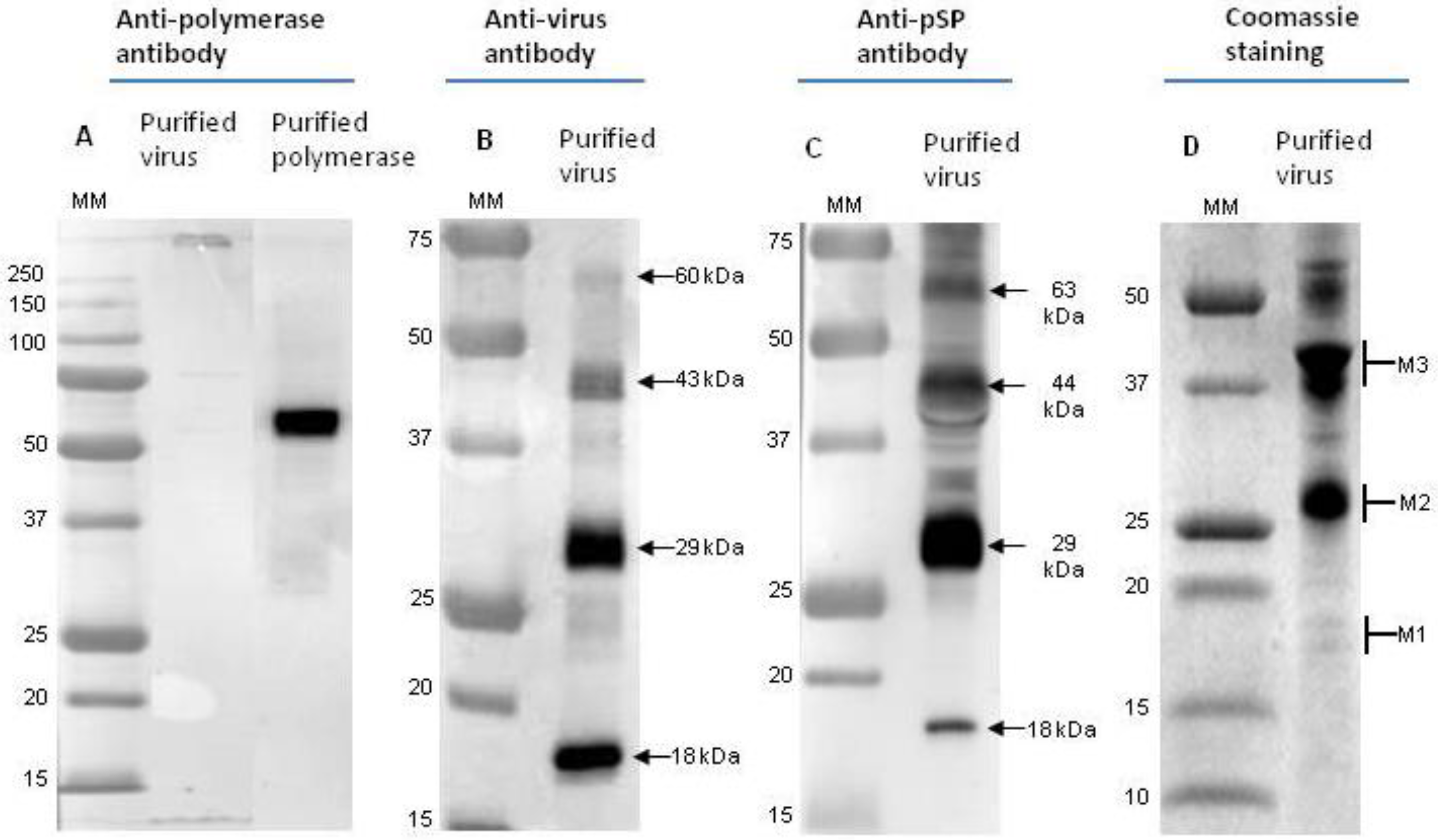
3.1. Western Blot Analysis of Virions
3.2. Detection of pSP and hSP Peptides by Mass Spectrometry


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Approximate Molecular Weight (kDa) | Viral Protein | Number of Peptides (Confidence of Identity > 95%) | Sequence of Identified Peptides |
|---|---|---|---|---|
| M1 | 18 | pSP | 1 | FIGDFITEHPEQTIGAVAVSAAVLSSAR |
| hSP | 2 | QIPIANFNEFLIK, LLEGPDEWLLVTAR | ||
| M2 | 29 | pSP | 4 | FIGDFITEHPEQTIGAVAVSAAVLSSAR, LQTVNALK, SILDR, SIVVTLGQK |
| M3 | 44 | pSP | 3 | FIGDFITEHPEQTIGAVAVSAAVLSSAR, LQTVNALK, SIVVTLGQK |
| hSP | 7 | QIPIANFNEFLIK, LFHYYPPLQLR, IDTQATLSELR, LLEGPDEWLLVTAR, FFPLQPLAR, ADNPDSLLEVLPVLVSADIK, YDWLGSGGSYCLVQPDR |

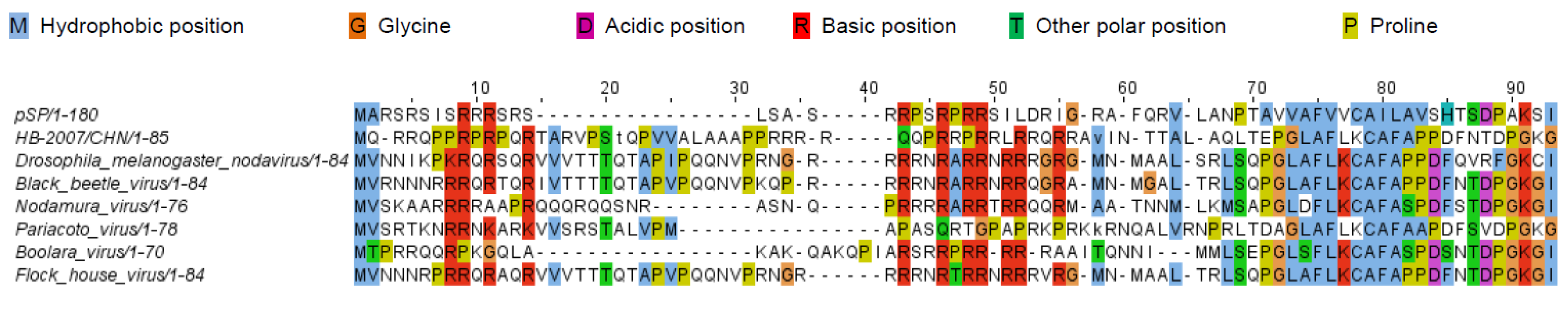
3.3. Analysis of the Amino Acid Sequences of pSP and hSP

4. Discussion

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ball, B.V.; Bailey, L. Viruses. In Honey Bee Pests, Predators, and Diseases, 3rd ed.; Morse, R.A., Flottum, K., Eds.; A.I. Root Company: Medina, OH, USA, 1997; pp. 11–32. [Google Scholar]
- Olivier, V.; Massou, I.; Celle, O.; Blanchard, P.; Schurr, F.; Ribière, M.; Gauthier, M. In situ hybridization assays for localization of the chronic bee paralysis virus in the honey bee (Apis mellifera) brain. J. Virol. Methods 2008, 153, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Bailey, L.; Gibbs, A.J.; Woods, R.D. The purification and properties of chronic bee-paralysis virus. J. Gen. Virol. 1968, 2, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Olivier, V.; Blanchard, P.; Chaouch, S.; Lallemand, P.; Schurr, F.; Celle, O.; Dubois, E.; Tordo, N.; Thiéry, R.; Houlgatte, R.; et al. Molecular characterisation and phylogenetic analysis of Chronic bee paralysis virus, a honey bee virus. Virus Res. 2008, 132, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Ribiere, M.; Olivier, V.; Blanchard, P. Chronic bee paralysis: A disease and a virus like no other? J. Invertebr. Pathol. 2010, 103, S120–S131. [Google Scholar] [CrossRef] [PubMed]
- Bailey, L. Viruses attacking the honey bee. In Advances in Virus Research; Lauffer, M.A., Bang, F.B., Maramorosch, K., Smith, K.M., Eds.; Academic Press: New York, NY, USA, 1976; Volume 20, pp. 271–304. [Google Scholar]
- Blanchard, P.; Schurr, F.; Olivier, V.; Celle, O.; Antúnez, K.; Bakonyi, T.; Berthoud, H.; Haubruge, E.; Higes, M.; Kasprzak, S.; et al. Phylogenetic analysis of the RNA-dependant RNA polymerase (RdRp) and a predicted structural protein (pSP) of the chronic bee paralysis virus (CBPV) isolated from various geographical regions. Virus Res. 2009, 144, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Runckel, C.; Flenniken, M.L.; Engel, J.C.; Ruby, J.G.; Ganem, D.; Andino, R.; DeRisi, J.L. Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, Nosema, and Crithidia. PLoS ONE 2011, 6, e20656. [Google Scholar] [CrossRef] [PubMed]
- Schuster, S.; Zirkel, F.; van Cleef, K.W.R.; Drosten, C.; van Rij, R.P.; Junglen, S. A unique nodavirus with novel features: Mosinovirus expresses two subgenomic RNAs, a capsid gene of unknown origin, and a suppressor of the antiviral RNA interference pathway. J. Virol. 2014, 88, 13447–13459. [Google Scholar] [CrossRef] [PubMed]
- Kuchibhatla, D.B.; Sherman, W.A.; Chung, B.Y.W.; Cook, S.; Schneider, G.; Eisenhaber, B.; Karlin, D.G. Powerful sequence similarity search methods and in-depth manual analyses can identify remote homologs in many apparently “Orphan” viral proteins. J. Virol. 2014, 88, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Kattenhorn, L.M.; Mills, R.; Wagner, M.; Lomsadze, A.; Makeev, V.; Borodovsky, M.; Ploegh, H.L.; Kessler, B.M. Identification of proteins associated with murine cytomegalovirus virions. J. Virol. 2004, 78, 11187–11197. [Google Scholar] [CrossRef] [PubMed]
- Renesto, P.; Abergel, C.; Decloquement, P.; Moinier, D.; Azza, S.; Ogata, H.; Fourquet, P.; Gorvel, J.P.; Claverie, J.M. Mimivirus giant particles incorporate a large fraction of anonymous and unique gene products. J. Virol. 2006, 80, 11678–11685. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Deng, F.; Hou, D.; Zhao, Y.; Guo, L.; Wang, H.; Hu, Z. Proteomics of the Autographa californica nucleopolyhedrovirus budded virions. J. Virol. 2010, 84, 7233–7242. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Ruan, H.Q.; Jiang, X.S.; Zhou, H.; Shi, L.; Zhang, L.; Sheng, Q.H.; Tu, Q.; Xia, Q.C.; Wu, J.R. Proteomic analysis of SARS associated coronavirus using two-dimensional liquid chromatography mass spectrometry and one-dimensional sodium dodecyl sulfate-polyacrylamide gel electrophoresis followed by mass spectrometric analysis. J. Proteome Res. 2004, 3, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Chertova, E.; Chertov, O.; Coren, L.V.; Roser, J.D.; Trubey, C.M.; Bess, J.W., Jr.; Sowder, R.C.; Barsov, E.; Hood, B.L.; Fisher, R.J.; et al. Proteomic and biochemical analysis of purified human immunodeficiency virus type 1 produced from infected monocyte-derived macrophages. J. Virol. 2006, 80, 9039–9052. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, P.; Ribière, M.; Celle, O.; Lallemand, P.; Schurr, F.; Olivier, V.; Iscache, A.L.; Faucon, J.P. Evaluation of a real-time two-step RT-PCR assay for quantitation of Chronic bee paralysis virus (CBPV) genome in experimentally-infected bee tissues and in life stages of a symptomatic colony. J. Virol. Methods 2007, 141, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, P.; Regnault, J.; Schurr, F.; Dubois, E.; Ribière, M. Intra-laboratory validation of chronic bee paralysis virus quantitation using an accredited standardised real-time quantitative RT-PCR method. J. Virol. Methods 2012, 180, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Bakonyi, T.; Grabensteiner, E.; Kolodziejek, J.; Rusvai, M.; Topolska, G.; Ritter, W.; Nowotny, N. Phylogenetic analysis of acute bee paralysis virus strains. Appl. Environ. Microbiol. 2002, 68, 6446–6450. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Grabensteiner, E.; Ritter, W.; Carter, M.J.; Davison, S.; Pechhacker, H.; Kolodziejek, J.; Boecking, O.; Derakhshifar, I.; Moosbeckhofer, R.; Licek, E.; et al. Sacbrood virus of the honeybee (Apis mellifera): rapid identification and phylogenetic analysis using reverse transcription-PCR. Clin. Diagn. Lab. Immunol. 2001, 8, 93–104. [Google Scholar] [PubMed]
- Lowry, O.H.; Rosebrough, A.L.; Farr, N.J.; Randall, R.J. Protein measurement with the Folin-Phenol reagents. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Lantez, V.; Dalle, K.; Charrel, R.; Baronti, C.; Canard, B.; Coutard, B. Comparative production analysis of three phlebovirus nucleoproteins under denaturing or non-denaturing conditions for crystallographic studies. PLoS Negl. Trop. Dis. 2011, 5, e936. [Google Scholar] [CrossRef] [PubMed]
- Ribiere, M.; Faucon, J.P.; Pépin, M. Detection of chronic bee paralysis virus infection: Application to a field survey. Apidologie 2000, 31, 567–577. [Google Scholar] [CrossRef]
- ProtParam. Available online: http://web.expasy.org/protparam (accessed on 20 March 2015).
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- PROTSCALE. Available online: http://web.expasy.org/protscale/ (accessed on 20 March 2015).
- Jones, D.T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Söndig, J. Protein homology detection by HMM-HMM comparison. Bioinformatics 2005, 21, 951–960. [Google Scholar]
- Kochentov, A.V.; Palyanov, A.; Titov, I.; Grigorovich, D.; Sarai, A.; Kolchanov, N.A. AUG_hairpin: Prediction of a downstream secondary structure influencing the recognition of a translation start site. BMC Bioinform. 2007, 8, e318. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2-A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Chung, B.Y.-W.; Bass, D.; Moureau, G.; Tang, S.; McAlister, E.; Culverwell, C.L.; Glücksman, A.; Wang, H.; Brown, T.D.K.; et al. Novel virus discovery and genome reconstruction from field RNA samples reveals highly divergent viruses in dipteran hosts. PLoS ONE 2013, 8, e80720. [Google Scholar] [CrossRef] [PubMed]
- Fang, N.X.; Frazer, I.H.; Zhou, J.; Fernando, G.J. Post translational modifications of recombinant human papillomavirus type 6b major capsid protein. Virus Res. 1999, 60, 113–121. [Google Scholar] [CrossRef]
- Geigenmuller, U.; Ginzton, N.H.; Matsui, S.M. Studies on intracellular processing of the capsid protein of human astrovirus serotype 1 in infected cells. J. Gen. Virol. 2002, 83, 1691–1695. [Google Scholar] [PubMed]
- Soldevila, A.I.; Huang, S.; Ghabrial, S.A. Assembly of the Hv190S totivirus capsid is independent of posttranslational modification of the capsid protein. Virology 1998, 251, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Knejzlík, Z.; Strohalm, M.; Sedlácková, L.; Kodícek, M.; Sakalian, M.; Ruml, T. Isolation and characterization of the Mason-Pfizer monkey virus p12 protein. Virology 2004, 324, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Curry, S.; Fry, E.; Blakemore, W.; Abu-Ghazaleh, R.; Jackson, T.; King, A.; Lea, S.; Newman, J.; Stuart, D. Dissecting the roles of VP0 cleavage and RNA packaging in picornavirus capsid stabilization: The structure of empty capsids of foot-and-mouth disease virus. J. Virol. 1997, 71, 9743–9752. [Google Scholar] [PubMed]
- Basavappa, R.; Syed, R.; Flore, O.; Icenogle, J.P.; Filman, D.J.; Hogle, J.M. Role and mechanism of the maturation cleavage of VP0 in poliovirus assembly: Structure of the empty capsid assembly intermediate at 2.9 Å resolution. Protein Sci. 1994, 3, 1651–1669. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, T.M.; Rueckert, R.R. Assembly-dependent maturation cleavage in provirions of a small icosahedral insect ribovirus. J. Virol. 1988, 62, 3399–3406. [Google Scholar] [PubMed]
- Kozak, M. Pushing the limits of the scanning mechanism for initiation of translation. Gene 2002, 299, 1–34. [Google Scholar] [CrossRef]
- Clyde, K.; Harris, E. RNA secondary structure in the coding region of dengue virus type 2 directs translation start codon selection and is required for viral replication. J. Virol. 2006, 80, 2170–2182. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.; Frankel, A.D. Structural variety of arginine-rich RNA-binding peptides. Proc. Natl. Acad. Sci. USA 1995, 92, 5282–5286. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.L.; Grantham, G.L. Molecular studies on bromovirus capsid protein. II. Functional analysis of the amino-terminal arginine-rich motif and its role in encapsidation, movement, and pathology. Virology 1996, 226, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.G.; Grantham, G.L.; Rao, A.L. Molecular studies on bromovirus capsid protein. Virology 2000, 270, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Venter, P.A.; Marshall, D.; Schneemann, A. Dual roles for an arginine-rich motif in specific genome recognition and localization of viral coat protein to RNA replication sites in flock house virus-infected cells. J. Virol. 2009, 83, 2872–2882. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Sit, T.L.; Kim, K.H.; Lommel, S.A. The Red clover necrotic mosaic virus capsid protein N-terminal lysine-rich motif is a determinant of symptomatology and virion accumulation. Mol. Plant Pathol. 2012, 13, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Johnson, J.E. Activation, exposure and penetration of virally encoded, membrane-active polypeptides during non-enveloped virus entry. Curr. Protein Pept. Sci. 2008, 9, 16–27. [Google Scholar] [PubMed]
- Chevin, A.; Schurr, F.; Blanchard, P.; Thiéry, R.; Ribière, M. Experimental infection of the honeybee (Apis mellifera L.) with chronic bee paralysis virus (CBPV): Infectivity of naked CBPV RNAs. Virus Res. 2012, 167, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Youssef, I.; Schurr, F.; Goulet, A.; Cougoule, N.; Ribière-Chabert, M.; Darbon, M.; Thiéry, R.; Dubois, E. RNA 1 and RNA 2 genomic segments of Chronic Bee Paralysis Virus (CBPV) are sufficient to recover an infectious virus. J. Immun. Res. Submitted.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chevin, A.; Coutard, B.; Blanchard, P.; Dabert-Gay, A.-S.; Ribière-Chabert, M.; Thiéry, R. Characterisation of Structural Proteins from Chronic Bee Paralysis Virus (CBPV) Using Mass Spectrometry. Viruses 2015, 7, 3329-3344. https://doi.org/10.3390/v7062774
Chevin A, Coutard B, Blanchard P, Dabert-Gay A-S, Ribière-Chabert M, Thiéry R. Characterisation of Structural Proteins from Chronic Bee Paralysis Virus (CBPV) Using Mass Spectrometry. Viruses. 2015; 7(6):3329-3344. https://doi.org/10.3390/v7062774
Chicago/Turabian StyleChevin, Aurore, Bruno Coutard, Philippe Blanchard, Anne-Sophie Dabert-Gay, Magali Ribière-Chabert, and Richard Thiéry. 2015. "Characterisation of Structural Proteins from Chronic Bee Paralysis Virus (CBPV) Using Mass Spectrometry" Viruses 7, no. 6: 3329-3344. https://doi.org/10.3390/v7062774
APA StyleChevin, A., Coutard, B., Blanchard, P., Dabert-Gay, A. -S., Ribière-Chabert, M., & Thiéry, R. (2015). Characterisation of Structural Proteins from Chronic Bee Paralysis Virus (CBPV) Using Mass Spectrometry. Viruses, 7(6), 3329-3344. https://doi.org/10.3390/v7062774