
Plant Virus–Insect Vector Interactions: Current and Potential Future Research Directions




{kind=link}
Abstract
:1. Introduction—Insects as Plant Virus Vectors
2. Types of Interactions of Plant Viruses with the Insect Vector
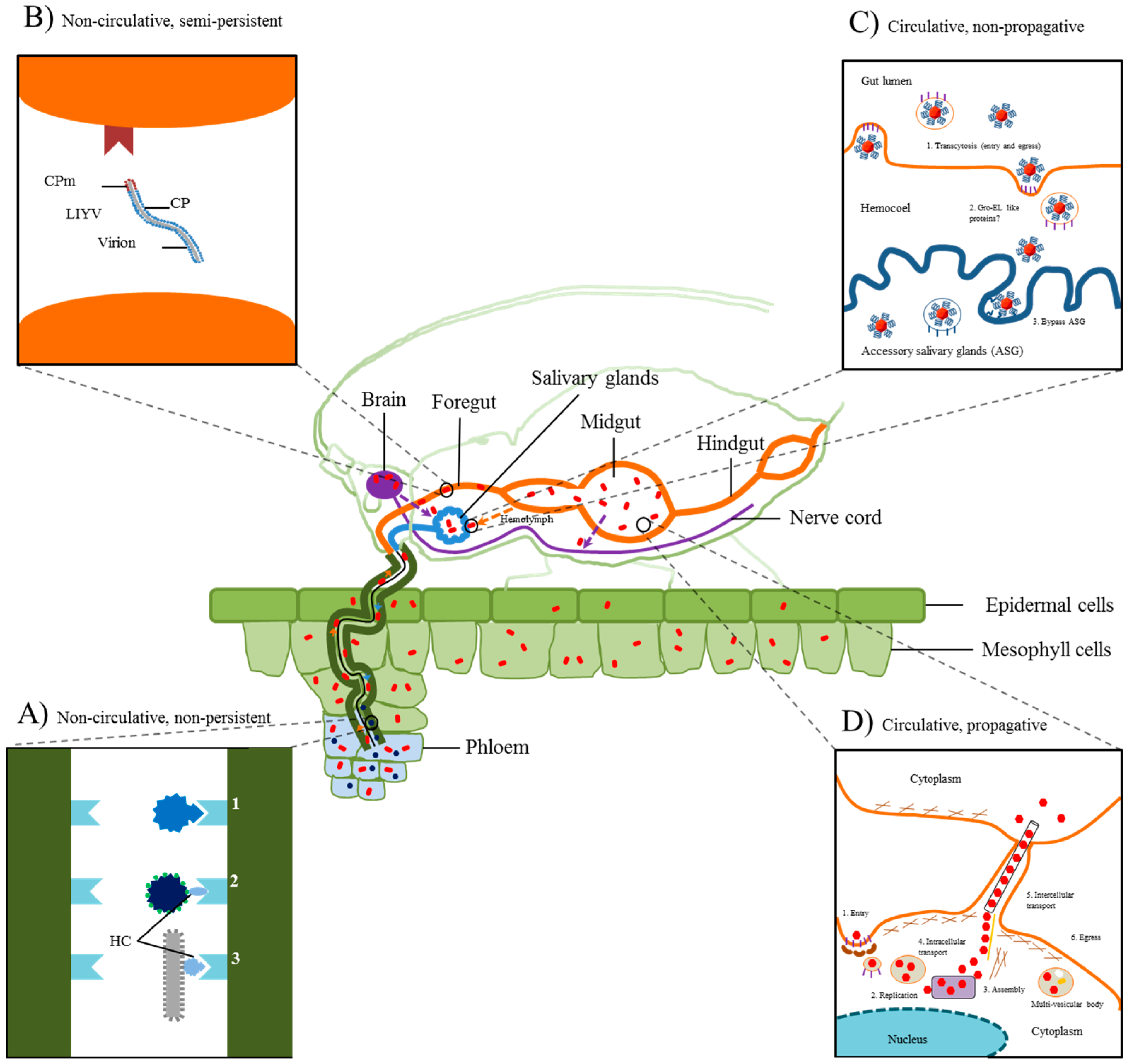
2.1. Non-Persistent Transmission
2.2. Non-Circulative, Semi-Persistent Transmission
2.3. Circulative, Non-Propagative Transmission
2.4. Circulative, Propagative Transmission
3. Critical Steps in Virus–Insect Interactions as Potential Control Targets
3.1. Stylet-Borne Viruses
3.2. Persistent Viruses
3.3. Plant Virus Effects on Vector Behaviour
4. What Can We Learn from Research on Virus Interactions with Vectors of Animal Viruses?
5. Significant Technological Advances in Vertebrate Virus—Insect Vector Control
6. Experimental Approaches to Facilitate Further Insights and Translational Research
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Bragard, C.; Caciagli, P.; Lemaire, O.; Lopez-Moya, J.J.; MacFarlane, S.; Peters, D.; Susi, P.; Torrance, L. Status and prospects of plant virus control through interference with vector transmission. Annu. Rev. Phytopathol. 2013, 51, 177–201. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Ammar, E.D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [PubMed]
- Ammar, E.D.; Tsai, C.W.; Whitfield, A.E.; Redinbaugh, M.G.; Hogenhout, S.A. Cellular and molecular aspects of rhabdovirus interactions with insect and plant hosts. Annu. Rev. Entomol. 2009, 54, 447–468. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.C.K.; Falk, B.W. Virus-vector interactions mediating nonpersistent and semipersistent transmission of plant viruses. Annu. Rev. Phytopathol. 2006, 44, 183–212. [Google Scholar] [CrossRef] [PubMed]
- Blanc, S.; Drucker, M.; Uzest, M. Localizing viruses in their insect vectors. Annu. Rev. Phytopathol. 2014, 52, 403–425. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, S.; Michalakis, Y.; van Munster, M.; Blanc, S. Plant feeding by insect vectors can affect life cycle, population genetics and evolution of plant viruses. Funct. Ecol. 2013, 27, 610–622. [Google Scholar] [CrossRef]
- Pirone, T.P.; Megahed, E. Aphid transmissibility of some purified viruses and viral RNAs. Virology 1966, 30, 631–637. [Google Scholar] [CrossRef]
- Chen, B.; Francki, R.I.B. Cucumovirus transmission by the aphid Myzus persicae is determined solely by the viral coat protein. J. Gen. Virol. 1990, 71, 939–944. [Google Scholar] [CrossRef]
- Liu, S.J.; He, X.H.; Park, G.; Josefsson, C.; Perry, K.L. A conserved capsid protein surface domain of Cucumber mosaic virus is essential for efficient aphid vector transmission. J. Virol. 2002, 76, 9756–9762. [Google Scholar] [CrossRef] [PubMed]
- Bak, A.; Gargani, D.; Macia, J.L.; Malouvet, E.; Vernerey, M.S.; Blanc, S.; Drucker, M. Virus factories of cauliflower mosaic virus are virion reservoirs that engage actively in vector transmission. J. Virol. 2013, 87, 12207–12215. [Google Scholar] [CrossRef] [PubMed]
- Leh, V.; Jacquot, E.; Geldreich, A.; Hermann, T.; Leclerc, D.; Cerutti, M.; Yot, P.; Keller, M.; Blanc, S. Aphid transmission of cauliflower mosaic virus requires the viral PIII protein. EMBO J. 1999, 18, 7077–7085. [Google Scholar] [CrossRef] [PubMed]
- Hoh, F.; Uzest, M.; Drucker, M.; Plisson-Chastang, C.; Bron, P.; Blanc, S.; Dumas, C. Structural insights into the molecular mechanisms of cauliflower mosaic virus transmission by its insect vector. J. Virol. 2010, 84, 4706–4713. [Google Scholar] [CrossRef] [PubMed]
- Plisson, C.; Uzest, M.; Drucker, M.; Froissart, M.; Dumas, C.; Conway, J.; Thomas, D.; Blanc, S.; Bron, P. Structure of the mature P3-virus particle complex of cauliflower mosaic virus revealed by cryo-electron microscopy. J. Mol. Biol. 2005, 346, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Martiniere, A.; Bak, A.; Macia, J.L.; Lautredou, N.; Gargani, D.; Doumayrou, J.; Garzo, E.; Moreno, A.; Fereres, A.; Blanc, S. A virus responds instantly to the presence of the vector on the host and forms transmission morphs. eLife 2013, 2, e00183. [Google Scholar] [CrossRef] [PubMed]
- Uzest, M.; Gargani, D.; Dombrovsky, A.; Cazevieille, C.; Cot, D.; Blanc, S. The “acrostyle”: A newly described anatomical structure in aphid stylets. Arthropod Struct. Dev. 2010, 39, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Uzest, M.; Gargani, D.; Drucker, M.; Hebrard, E.; Garzo, E.; Candresse, T.; Fereres, A.; Blanc, S. A protein key to plant virus transmission at the tip of the insect vector stylet. Proc. Natl. Acad. Sci. USA 2007, 104, 17959–17964. [Google Scholar] [CrossRef] [PubMed]
- Govier, D.A.; Kassanis, B. Virus-induced component of plant sap needed when aphids acquire potato virus Y from purified preparations. Virology 1974, 61, 420–426. [Google Scholar] [CrossRef]
- Wang, R.Y.; Ammar, E.D.; Thornbury, D.W.; LopezMoya, J.J.; Pirone, T.P. Loss of potyvirus transmissibility and helper-component activity correlate with non-retention of virions in aphid stylets. J. Gen. Virol. 1996, 77, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Blanc, S.; Ammar, E.D.; Garcia-Lampasona, S.; Dolja, V.V.; Llave, C.; Baker, J.; Pirone, T.P. Mutations in the potyvirus helper component protein: Effects on interactions with virions and aphid stylets. J. Gen. Virol. 1998, 79, 3119–3122. [Google Scholar] [CrossRef] [PubMed]
- Dombrovsky, A.; Gollop, N.; Chen, S.B.; Chejanovsky, N.; Raccah, B. In vitro association between the helper component-proteinase of zucchini yellow mosaic virus and cuticle proteins of Myzus persicae. J. Gen. Virol. 2007, 88, 1602–1610. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.K.; Kang, S.H.; Seo, B.Y.; Jung, J.K.; Kim, K.H. Mutational analysis of interaction between coat protein and helper component-proteinase of Soybean mosaic virus involved in aphid transmission. Mol. Plant Pathol. 2010, 11, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Kontra, L.; Burgyan, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziebell, H.; Murphy, A.M.; Groen, S.C.; Tungadi, T.; Westwood, J.H.; Lewsey, M.G.; Moulin, M.; Kleczkowski, A.; Smith, A.G.; Stevens, M.; et al. Cucumber mosaic virus and its 2b RNA silencing suppressor modify plant-aphid interactions in tobacco. Sci. Rep. 2011, 1, 187. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.C.K.; Zhou, J.S. Insect vector-plant virus interactions associated with non-circulative, semi-persistent transmission: Current perspectives and future challenges. Curr. Opin. Virol. 2015, 15, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.Y.S.; Walker, G.P.; Carter, D.; Ng, J.C.K. A virus capsid component mediates virion retention and transmission by its insect vector. Proc. Natl. Acad. Sci. USA 2011, 108, 16777–16782. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.R.; Medina, V.; Tian, T.Y.; Turina, M.; Falk, B.W.; Ng, J.C.K. A mutation in the Lettuce infectious yellows virus minor coat protein disrupts whitefly transmission but not in planta systemic movement. J. Virol. 2010, 84, 12165–12173. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, P.V.; Kliot, A.; Ghanim, M.; Cilia, M. Is there a role for symbiotic bacteria in plant virus transmission by insects? Curr. Opin. Insect Sci. 2015, 8, 69–78. [Google Scholar] [CrossRef]
- Gray, S.; Cilia, M.; Ghanim, M. Circulative, “nonpropagative” virus transmission: An orchestra of virus-, insect-, and plant-derived instruments. Adv. Virus Res. 2014, 89, 141–199. [Google Scholar] [PubMed]
- Garret, A.; Kerlan, C.; Thomas, D. Ultrastructural study of acquisition and retention of potato leafroll luteovirus in the alimentary canal of its aphid vector, Myzus persicae Sulz. Arch. Virol. 1996, 141, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Gildow, F.E. Evidence for receptor-mediated endocytosis regulating luteovirus acquisition by aphids. Phytopathol. 1993, 83, 270–277. [Google Scholar] [CrossRef]
- Reinbold, C.; Gildow, F.E.; Herrbach, E.; Ziegler-Graff, V.; Goncalves, M.C.; van den Heuvel, J.F.J.M.; Brault, V. Studies on the role of the minor capsid protein in transport of Beet western yellows virus through Myzus persicae. J. Gen. Virol. 2001, 82, 1995–2007. [Google Scholar] [CrossRef] [PubMed]
- Reinbold, C.; Herrbach, E.; Brault, V. Posterior midgut and hindgut are both sites of acquisition of Cucurbit aphid-bome yellows virus in Myzus persicae and Aphis gossypii. J. Gen. Virol. 2003, 84, 3473–3484. [Google Scholar] [CrossRef] [PubMed]
- Brault, V.; Mutterer, J.; Scheidecker, D.; Simonis, M.T.; Herrbach, E.; Richards, K.; Ziegler-Graff, V. Effects of point mutations in the readthrough domain of the beet western yellows virus minor capsid protein on virus accumulation in planta and on transmission by aphids. J. Virol. 2000, 74, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Sivakumar, S.; Wang, Z.H.; Bonning, B.C.; Miller, W. The readthrough domain of pea enation mosaic virus coat protein is not essential for virus stability in the hemolymph of the pea aphid. Arch. Virol. 2009, 154, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Peter, K.A.; Liang, D.L.; Palukaitis, P.; Gray, S.M. Small deletions in the potato leafroll virus readthrough protein affect particle morphology, aphid transmission, virus movement and accumulation. J. Gen. Virol. 2008, 89, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Bruyere, A.; Brault, V.; ZieglerGraff, V.; Simonis, M.T.; VandenHeuvel, J.F.J.M.; Richards, K.; Guilley, H.; Jonard, G.; Herrbach, E. Effects of mutations in the beet western yellows virus readthrough protein on its expression and packaging and on virus accumulation, symptoms, and aphid transmission. Virology 1997, 230, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Cilia, M.; Peter, K.A.; Bereman, M.S.; Howe, K.; Fish, T.; Smith, D.; Gildow, F.; MacCoss, M.J.; Thannhauser, T.W.; Gray, S.M. Discovery and targeted LC-MS/MS of purified Polerovirus reveals differences in the virus-host interactome associated with altered aphid transmission. PLoS ONE 2012, 7, e48177. [Google Scholar] [CrossRef] [PubMed]
- Cicero, J.M.; Brown, J.K. Functional anatomy of whitefly organs associated with squash leaf curl Virus (Geminiviridae: Begomovirus) transmission by the B biotype of Bemisia tabaci (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2011, 104, 261–279. [Google Scholar] [CrossRef]
- Ghanim, M.; Morin, S.; Czosnek, H. Rate of Tomato yellow leaf curl virus translocation in the circulative transmission pathway of its vector, the whitefly Bemisia tabaci. Phytopathology 2001, 91, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Pinner, M.S.; Stanley, J.; Markham, P.G. Geminivirus coat protein gene replacement alters insect specificity. Virology 1990, 177, 85–94. [Google Scholar] [CrossRef]
- Wang, L.L.; Wei, X.M.; Ye, X.D.; Xu, H.X.; Zhou, X.P.; Liu, S.S.; Wang, X.W. Expression and functional characterisation of a soluble form of Tomato yellow leaf curl virus coat protein. Pest Manag. Sci. 2014, 70, 1624–1631. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, M. A review of the mechanisms and components that determine the transmission efficiency of Tomato yellow leaf curl virus (Geminiviridae, Begomovirus) by its whitefly vector. Virus Res. 2014, 186, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Rosen, R.; Kanakala, S.; Kliot, A.; Pakkianathan, B.C.; Abu Farich, B.; Santana-Magal, N.; Elimelech, M.; Kontsedalov, S.; Lebedev, G.; Cilia, M.; et al. Persistent, circulative transmission of begomoviruses by whitefly vectors. Curr. Opin. Virol. 2015, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Chen, Q.; Omura, T.; Uehara-Ichiki, T.; Wei, T.Y. Sequential infection of rice dwarf virus in the internal organs of its insect vector after ingestion of virus. Virus Res. 2011, 160, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Omura, T.; Yan, J.; Zhong, B.X.; Wada, M.; Zhu, Y.F.; Tomaru, M.; Maruyama, W.; Kikuchi, A.; Watanabe, Y.; Kimura, I. The P2 protein of rice dwarf phytoreovirus is required for adsorption of the virus to cells of the insect vector. J. Virol. 1998, 72, 9370–9373. [Google Scholar] [PubMed]
- Wei, T.Y.; Chen, H.Y.; Ichiki-Uehara, T.; Hibino, H.; Omura, T. Entry of Rice dwarf virus into cultured cells of its insect vector involves clathrin-mediated endocytosis. J. Virol. 2007, 81, 7811–7815. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Hibino, H.; Omura, T. Release of rice dwarf virus from insect vector cells involves secretory exosomes derived from multivesicular bodies. Commun. Integr. Biol. 2009, 2, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Pu, Y.; Weit, T.; Liu, H.; Deng, W.; Wei, C.; Ding, B.; Omura, T.; Lin, Y. The P2 capsid protein of the nonenveloped rice dwarf phytoreovirus induces membrane fusion in insect host cells. Proc. Natl. Acad. Sci. USA 2007, 104, 19547–19552. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Chen, H.Y.; Mao, Q.Z.; Liu, Q.F.; Shimizu, T.; Uehara-Ichiki, T.; Wu, Z.J.; Xie, L.H.; Omura, T.; Wei, T.Y. Tubular structure induced by a plant virus facilitates viral spread in its vector insect. PLoS Pathog. 2012, 8, e1003032. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, H.T.; Ren, T.Y.; Xie, L.H.; Wei, T.Y. Interaction between non-structural protein Pns10 of rice dwarf virus and cytoplasmic actin of leafhoppers is correlated with insect vector specificity. J. Gen. Virol. 2015, 96, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jia, D.S.; Chen, H.Y.; Chen, Q.; Xie, L.H.; Wu, Z.J.; Wei, T.Y. The P7-1 protein of southern rice black-streaked dwarf virus, a fijivirus, induces the formation of tubular structures in insect cells. Arch. Virol. 2011, 156, 1729–1736. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.S.; Mao, Q.Z.; Chen, H.Y.; Wang, A.M.; Liu, Y.Y.; Wang, H.T.; Xie, L.H.; Wei, T.Y. Virus-induced tubule: A vehicle for rapid spread of virions through basal lamina from midgut epithelium in the insect vector. J. Virol. 2014, 88, 10488–10500. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Wei, T.Y.; Hagiwara, K.; Higashi, T.; Kimura, I.; Akutsu, K.; Omura, T. Retention of Rice dwarf virus by descendants of pairs of viruliferous vector insects after rearing for 6 years. Phytopathology 2007, 97, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Pu, L.L.; Xie, G.H.; Ji, C.Y.; Ling, B.; Zhang, M.X.; Xu, D.L.; Zhou, G.H. Transmission characteristics of Southern rice black-streaked dwarf virus by rice planthoppers. Crop Protect. 2012, 41, 71–76. [Google Scholar] [CrossRef]
- Lewandowski, D.J.; Adkins, S. The tubule-forming NSm protein from Tomato spotted wilt virus complements cell-to-cell and long-distance movement of Tobacco mosaic virus hybrids. Virology 2005, 342, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Storms, M.M.H.; Kormelink, R.; Peters, D.; vanLent, J.W.M.; Goldbach, R.W. The nonstructural NSm protein of tomato spotted wilt virus induces tubular structures in plant and insect cells. Virology 1995, 214, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Moritz, G.; Kumm, S.; Mound, L. Tospovirus transmission depends on thrips ontogeny. Virus Res. 2004, 100, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Inoue-Nagata, A.K.; Smid, H.M.; Goldbach, R.; Peters, D. Tissue tropism related to vector competence of Frankliniella occidentalis for tomato spotted wilt tospovirus. J. Gen. Virol. 1999, 80, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Inoue-Nagata, A.K.; Prins, M.; Goldbach, R.; Peters, D. Impeded thrips transmission of defective tomato spotted wilt virus isolates. Phytopathology 2000, 90, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Resende, R.D.; Dehaan, P.; Deavila, A.C.; Kitajima, E.W.; Kormelink, R.; Goldbach, R.; Peters, D. Generation of envelope and defective interfering RNA mutants of tomato spotted wilt virus by mechanical passage. J. Gen. Virol. 1991, 72, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Sin, S.H.; McNulty, B.C.; Kennedy, G.G.; Moyer, J.W. Viral genetic determinants for thrips transmission of tomato spotted wilt virus. Proc. Natl. Acad. Sci. USA 2005, 102, 5168–5173. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Ullman, D.E.; German, T.L. Expression and characterization of a soluble form of tomato spotted wilt virus glycoprotein Gn. J. Virol. 2004, 78, 13197–13206. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Ullman, D.E.; German, T.L. Tomato spotted wilt virus glycoprotein Gc is cleaved at acidic pH. Virus Res. 2005, 110, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Garry, C.E.; Garry, R.F. Proteomics computational analyses suggest that the carboxyl terminal glycoproteins of bunyaviruses are class II viral fusion protein (beta-penetrenes). Theor. Biol. Med. Mod. 2004, 1, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badillo-Vargas, I.E.; Rotenberg, D.; Schneweis, D.J.; Hiromasa, Y.; Tomich, J.M.; Whitfield, A.E. Proteomic analysis of Frankliniella occidentalis and differentially expressed proteins in response to Tomato spotted wilt virus infection. J. Virol. 2012, 86, 8793–8809. [Google Scholar] [CrossRef] [PubMed]
- Rotenberg, D.; Whitfield, A.E. Analysis of expressed sequence tags for Frankliniella occidentalis, the western flower thrips. Insect Mol. Biol. 2010, 19, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Nault, L.R.; Ammar, E. Leafhopper and planthopper transmission of plant-viruses. Annu. Rev. Entomol. 1989, 34, 503–529. [Google Scholar] [CrossRef]
- Zheng, L.M.; Mao, Q.Z.; Xie, L.H.; Wei, T.Y. Infection route of rice grassy stunt virus, a tenuivirus, in the body of its brown planthopper vector, Nilaparvata lugens (Hemiptera: Delphacidae) after ingestion of virus. Virus Res. 2014, 188, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.H.; Li, S.; Hong, J.; Ji, Y.H.; Zhou, Y.J. Investigation on subcellular localization of in its vector small brown planthopper by electron microscopy. Virol. J. 2013, 10, 310. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zheng, L.M.; Chen, H.Y.; Jia, D.S.; Li, F.; Wei, T.Y. Nonstructural protein NS4 of Rice stripe virus plays a critical role in viral spread in the body of vector insects. PLoS ONE 2014, 9, e88636. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.L.; Zhang, G.Z.; Dai, X.J.; Hou, Y.L.; Li, M.; Liang, J.S.; Liang, C.Y. Processing and intracellular localization of rice stripe virus Pc2 protein in insect cells. Virology 2012, 429, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Liu, X.F.; Li, S.; Xu, Y.; Zhou, Y.J.; Zhou, X.P.; Tao, X.R. Rice stripe tenuivirus NSvc2 glycoproteins targeted to the golgi body by the N-terminal transmembrane domain and adjacent cytosolic 24 amino acids via the COP I- and COP II-dependent secretion pathway. J. Virol. 2014, 88, 3223–3234. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Liu, W.W.; Zhang, F.J.; Chen, X.Y.; Li, L.; Liu, Q.F.; Zhou, Y.J.; Wei, T.Y.; Fang, R.X.; Wang, X.F. Transovarial transmission of a plant virus is mediated by vitellogenin of its insect vector. PLoS Pathog. 2014, 10, e1003949. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.O.; Dietzgen, R.G.; Goodin, M.M.; Bragg, J.N.; Deng, M. Biology of plant rhabdoviruses. Annu. Rev. Phytopathol. 2005, 43, 623–660. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.S.; Dietzgen, R.G. Plant rhabdoviruses: New insights and research needs in the interplay of negative-strand RNA viruses with plant and insect hosts. Arch. Virol. 2014, 159, 1889–1900. [Google Scholar] [CrossRef] [PubMed]
- Chapman, R.F. The Insects: Structure and Function, 4th ed.; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Ammar, E.D.; Hogenhout, S.A. A neurotropic route for Maize mosaic virus (Rhabdoviridae) in its planthopper vector Peregrinus maidis. Virus Res. 2008, 131, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Cassone, B.J.; Wijeratne, S.; Michel, A.P.; Stewart, L.R.; Chen, Y.T.; Yan, P.; Redinbaugh, M.G. Virus-independent and common transcriptome responses of leafhopper vectors feeding on maize infected with semi-persistently and persistent propagatively transmitted viruses. BMC Genom. 2014, 15, 133. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Rotenberg, D. Disruption of insect transmission of plant viruses. Curr. Opin. Insect Sci. 2015, 8, 79–87. [Google Scholar] [CrossRef]
- Drucker, M.; Then, C. Transmission activation in non-circulative virus transmission: A general concept? Curr. Opin. Virol. 2015, 15, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Redinbaugh, M.G.; Ammar, E.D. Plant and animal rhabdovirus host range: A bug’s view. Trends Microbiol. 2003, 11, 264–271. [Google Scholar] [CrossRef]
- Rotenberg, D.; Jacobson, A.L.; Schneweis, D.J.; Whiffleld, A.E. Thrips transmission of tospoviruses. Curr. Opin. Virol. 2015, 15, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Ullman, D.E.; Whitfield, A.E.; German, T.L. Thrips and tospoviruses come of age: Mapping determinants of insect transmission. Proc. Natl. Acad. Sci. USA 2005, 102, 4931–4932. [Google Scholar] [CrossRef] [PubMed]
- Linz, L.B.; Liu, S.; Chougule, N.P.; Bonning, B.C. In vitro evidence supports membrane alanyl aminopeptidase N as a receptor for a plant virus in the pea aphid vector. J. Virol. 2015, 89, 11203–11212. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, R.B.; Resende, R.D.; de Avila, A.C. The plant virus Tomato spotted wilt virus activates the immune system of its main insect vector, Frankliniella occidentalis. J. Virol. 2004, 78, 4976–4982. [Google Scholar] [CrossRef] [PubMed]
- Cassone, B.J.; Carter, F.M.C.; Michel, A.P.; Stewart, L.R.; Redinbaugh, M.G. Genetic insights into Graminella nigrifrons competence for Maize fine streak virus infection and transmission. PLoS ONE 2014, 9, e113529. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Redinbaugh, M.G.; Michel, A.P. Molecular interactions and immune responses between Maize fine streak virus and the leafhopper vector Graminella nigrifrons through differential expression and RNA interference. Insect Mol. Biol. 2015, 24, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Rotenberg, D.; Aritua, V.; Hogenhout, S.A. Analysis of expressed sequence tags from Maize mosaic rhabdovirus-infected gut tissues of Peregrinus maidis reveals the presence of key components of insect innate immunity. Insect Mol. Biol. 2011, 20, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.H.; Wang, H.T.; Chen, Q.; Chen, H.Y.; Jia, D.S.; Mao, Q.Z.; Wei, T.Y. Small interfering RNA pathway modulates persistent infection of a plant virus in its insect vector. Sci. Rep. 2016, 6, 20699. [Google Scholar] [CrossRef] [PubMed]
- Stafford-Banks, C.A.; Rotenberg, D.; Johnson, B.R.; Whitfield, A.E.; Ullman, D.E. Analysis of the salivary gland transcriptome of Frankliniella occidentalis. PLoS ONE 2014, 9, e94447. [Google Scholar] [CrossRef] [PubMed]
- Montero-Astua, M.; Ullman, D.E.; Whitfield, A.E. Salivary gland morphology, tissue tropism and the progression of tospovirus infection in Frankliniella occidentalis. Virology 2016, 493, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Blanc, S.; Michalakis, Y. Manipulation of hosts and vectors by plant viruses and impact of the environment. Curr. Opin. Insect Sci. 2016, 16, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Stafford, C.A.; Walker, G.P.; Ullman, D.E. Infection with a plant virus modifies vector feeding behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 9350–9355. [Google Scholar] [CrossRef] [PubMed]
- Carmo-Sousa, M.; Moreno, A.; Garzo, E.; Fereres, A. A non-persistently transmitted-virus induces a pull-push strategy in its aphid vector to optimize transmission and spread. Virus Res. 2014, 186, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.M.; Preisser, E.L.; Chu, D.; Pan, H.P.; Xie, W.; Wang, S.L.; Wu, Q.J.; Zhou, X.G.; Zhang, Y.J. Multiple forms of vector manipulation by a plant-infecting virus: Bemisia tabaci and Tomato yellow leaf curl virus. J. Virol. 2013, 87, 4929–4937. [Google Scholar] [CrossRef] [PubMed]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Biochemical and physiological mechanisms underlying effects of Cucumber mosaic virus on host-plant traits that mediate transmission by aphid vectors. Plant Cell Environ. 2014, 37, 1427–1439. [Google Scholar] [CrossRef] [PubMed]
- Bosque-Perez, N.A.; Eigenbrode, S.D. The influence of virus-induced changes in plants on aphid vectors: Insights from luteovirus pathosystems. Virus Res. 2011, 159, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Rajabaskar, D.; Bosque-Perez, N.A.; Eigenbrode, S.D. Preference by a virus vector for infected plants is reversed after virus acquisition. Virus Res. 2014, 186, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Ingwell, L.L.; Eigenbrode, S.D.; Bosque-Perez, N.A. Plant viruses alter insect behavior to enhance their spread. Sci. Rep. 2012, 2, 578. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, D.L.; Pu, L.L.; Zhou, G.H. Southern rice black-streaked dwarf virus alters insect vectors’ host orientation preferences to enhance spread and increase Rice ragged stunt virus co-infection. Phytopathology 2014, 104, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Casteel, C.L.; Yang, C.L.; Nanduri, A.C.; De Jong, H.N.; Whitham, S.A.; Jander, G. The NIa-Pro protein of Turnip mosaic virus improves growth and reproduction of the aphid vector, Myzus persicae (green peach aphid). Plant J. 2014, 77, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Carn, V.M. The role of dipterous insects in the mechanical transmission of animal viruses. Br. Vet. J. 1996, 152, 377–393. [Google Scholar] [CrossRef]
- Day, M.F.; Fenner, F.; Woodroofe, G.M.; McIntyre, G.A. Further studies on the mechanism of mosquito transmission of myxomatosis in the European rabbit. J. Hyg. (Lond.) 1956, 54, 258–283. [Google Scholar] [CrossRef] [PubMed]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Mechanical transmission of lumpy skin disease virus by Aedes aegypti (Diptera: Culicidae). Epidemiol. Infect. 2001, 126, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Attempted mechanical transmission of lumpy skin disease virus by biting insects. Med. Vet. Entomol. 2003, 17, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Blanc, S.; Gutierrez, S. The specifics of vector transmission of arboviruses of vertebrates and plants. Curr. Opin. Virol. 2015, 15, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed]
- Aponte, H.A.; Penilla, R.P.; Dzul-Manzanilla, F.; Che-Mendoza, A.; Lopez, A.D.; Solis, F.; Manrique-Saide, P.; Ranson, H.; Lenhart, A.; McCall, P.J.; et al. The pyrethroid resistance status and mechanisms in Aedes aegypti from the Guerrero state, Mexico. Pestic. Biochem. Phys. 2013, 107, 226–234. [Google Scholar] [CrossRef]
- Bisset, J.A.; Marin, R.; Rodriguez, M.M.; Severson, D.W.; Ricardo, Y.; French, L.; Diaz, M.; Perez, O. Insecticide resistance in two Aedes aegypti (Diptera: Culicidae) strains from Costa Rica. J. Med. Entomol. 2013, 50, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L. Quantitative genetics of Aedes aegypti vector competence for dengue viruses: Towards a new paradigm? Trends Parasitol. 2011, 27, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Wijkamp, I.; Almarza, N.; Goldbach, R.; Peters, D. Distinct levels of specificity in thrips transmission of tospoviruses. Phytopathology 1995, 85, 1069–1074. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Kontsedalov, S.; Skaljac, M.; Brumin, M.; Sobol, I.; Czosnek, H.; Vavre, F.; Fleury, F.; et al. The transmission efficiency of tomato yellow leaf curl virus by the whitefly Bemisia tabaci is correlated with the presence of a specific symbiotic bacterium species. J. Virol. 2010, 84, 9310–9317. [Google Scholar] [CrossRef] [PubMed]
- Bedford, I.D.; Briddon, R.W.; Brown, J.K.; Rosell, R.C.; Markham, P.G. Geminivirus-transmission and biological characterization of Bemisia tabaci (Gennadius) biotypes from different geographic regions. Ann. Appl. Biol. 1994, 125, 311–325. [Google Scholar] [CrossRef]
- Franz, A.W.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue barriers to arbovirus infection in mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Liu, Y.; Wang, P.; Xiao, X. Mosquito defense strategies against viral infection. Trends Parasitol. 2016, 32, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Jupatanakul, N.; Dimopoulos, G. Mosquito immunity against arboviruses. Viruses 2014, 6, 4479–4504. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, R.G.; Kang, S.; Simoes, M.L.; Anglero-Rodriguez, Y.I.; Dimopoulos, G. Mosquito gut antiparasitic and antiviral immunity. Dev. Comp. Immunol. 2016, 64, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Lamiable, O.; Imler, J.L. Induced antiviral innate immunity in Drosophila. Curr. Opin. Microbiol. 2014, 20, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, M.B.; Huang, Z.; Hardy, R.W. Insect antiviral innate immunity: Pathways, effectors, and connections. J. Mol. Biol. 2013, 425, 4921–4936. [Google Scholar] [CrossRef] [PubMed]
- Goic, B.; Vodovar, N.; Mondotte, J.A.; Monot, C.; Frangeul, L.; Blanc, H.; Gausson, V.; Vera-Otarola, J.; Cristofari, G.; Saleh, M.C. RNA-mediated interference and reverse transcription control the persistence of RNA viruses in the insect model Drosophila. Nat. Immunol. 2013, 14, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Hegde, S.; Rasgon, J.L.; Hughes, G.L. The microbiome modulates arbovirus transmission in mosquitoes. Curr. Opin. Virol. 2015, 15, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Minard, G.; Mavingui, P.; Moro, C.V. Diversity and function of bacterial microbiota in the mosquito holobiont. Parasite Vector 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Short, S.M.; Bahia, A.C.; Saraiva, R.G.; Dong, Y.; Kang, S.; Tripathi, A.; Mlambo, G.; Dimopoulos, G. Chromobacterium Csp_P reduces malaria and dengue infection in vector mosquitoes and has entomopathogenic and in vitro anti-pathogen activities. PLoS Pathog. 2014, 10, e1004398. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Souza-Neto, J.; Cosme, R.T.; Rovira, J.; Ortiz, A.; Pascale, J.M.; Dimopoulos, G. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and Dengue virus influences vector competence. PLoS Neglect. Trop. Dis. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Apte-Deshpande, A.; Paingankar, M.; Gokhale, M.D.; Deobagkar, D.N. Serratia odorifera a midgut inhabitant of Aedes aegypti mosquito enhances its susceptibility to Dengue-2 virus. PLoS ONE 2012, 7, e40401. [Google Scholar] [CrossRef] [PubMed]
- Apte-Deshpande, A.D.; Paingankar, M.S.; Gokhale, M.D.; Deobagkar, D.N. Serratia odorifera mediated enhancement in susceptibility of Aedes aegypti for Chikungunya virus. Indian J. Med. Res. 2014, 139, 762–768. [Google Scholar] [PubMed]
- Carissimo, G.; Pondeville, E.; McFarlane, M.; Dietrich, I.; Mitri, C.; Bischoff, E.; Antoniewski, C.; Bourgouin, C.; Failloux, A.B.; Kohl, A.; et al. Antiviral immunity of Anopheles gambiae is highly compartmentalized, with distinct roles for RNA interference and gut microbiota. Proc. Natl. Acad. Sci. USA 2015, 112, E176–E185. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti Toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Joyce, J.D.; Nogueira, J.R.; Bales, A.A.; Pittman, K.E.; Anderson, J.R. Interactions between La Crosse virus and bacteria isolated from the digestive tract of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2011, 48, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Humphery-Smith, I.; Grulet, O.; Chastel, C. Pathogenicity of Spiroplasma taiwanense for larval Aedes aegypti mosquitoes. Med. Vet. Entomol. 1991, 5, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Humphery-Smith, I.; Grulet, O.; Le Goff, F.; Chastel, C. Spiroplasma (Mollicutes: Spiroplasmataceae) pathogenic for Aedes aegypti and Anopheles stephensi (Diptera: Culicidae). J. Med. Entomol. 1991, 28, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Williamson, D.L.; Tully, J.G.; Rosen, L.; Rose, D.L.; Whitcomb, R.F.; Abalain-Colloc, M.L.; Carle, P.; Bove, J.M.; Smyth, J. Spiroplasma diminutum sp. nov., from Culex annulus mosquitoes collected in Taiwan. Int. J. Syst. Bacteriol. 1996, 46, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Ferreira, A.; Ashburner, M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e1000002. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and virus protection in insects. Science 2008, 322, 702. [Google Scholar] [CrossRef] [PubMed]
- Osborne, S.E.; Leong, Y.S.; O’Neill, S.L.; Johnson, K.N. Variation in antiviral protection mediated by different Wolbachia strains in Drosophila simulans. PLoS Pathog. 2009, 5, e1000656. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.N. Bacteria and antiviral immunity in insects. Curr. Opin. Insect Sci. 2015, 8, 97–103. [Google Scholar] [CrossRef]
- Johnson, K.N. The impact of Wolbachia on virus infection in mosquitoes. Viruses 2015, 7, 5705–5717. [Google Scholar] [CrossRef] [PubMed]
- Blagrove, M.S.; Arias-Goeta, C.; Failloux, A.B.; Sinkins, S.P. Wolbachia strain wMel induces cytoplasmic incompatibility and blocks dengue transmission in Aedes albopictus. Proc. Natl. Acad. Sci. USA 2012, 109, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.W.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z.Y. The endosymbiotic bacterium Wolbachia induces resistance to Dengue virus in Aedes aegypti. PLoS Pathog. 2010, 6, e1000833. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.J.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Rainey, S.M.; Shah, P.; Kohl, A.; Dietrich, I. Understanding the Wolbachia-mediated inhibition of arboviruses in mosquitoes: Progress and challenges. J. Gen. Virol. 2014, 95, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Dutra, H.L.; Moreira, L.A. Exploiting intimate relationships: Controlling mosquito-transmitted disease with Wolbachia. Trends Parasitol. 2016, 32, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Bourtzis, K.; Dobson, S.L.; Xi, Z.; Rasgon, J.L.; Calvitti, M.; Moreira, L.A.; Bossin, H.C.; Moretti, R.; Baton, L.A.; Hughes, G.L.; et al. Harnessing mosquito-Wolbachia symbiosis for vector and disease control. Acta Trop. 2014, 132, S150–S163. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis, N.; Tesh, R.B. Insect-specific viruses and their potential impact on arbovirus transmission. Curr. Opin. Virol. 2015, 15, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Kean, J.; Rainey, S.M.; McFarlane, M.; Donald, C.L.; Schnettler, E.; Kohl, A.; Pondeville, E. Fighting arbovirus transmission: Natural and engineered control of vector competence in Aedes mosquitoes. Insects 2015, 6, 236–278. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Hobson-Peters, J.; Yam, A.W.; Lu, J.W.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; Davis, S.S.; Weir, R.; et al. A new insect-specific flavivirus from northern Australia suppresses replication of West Nile virus and Murray Valley encephalitis virus in co-infected mosquito cells. PLoS ONE 2013, 8, e56534. [Google Scholar] [CrossRef] [PubMed]
- Kenney, J.L.; Solberg, O.D.; Langevin, S.A.; Brault, A.C. Characterization of a novel insect-specific flavivirus from Brazil: Potential for inhibition of infection of arthropod cells with medically important flaviviruses. J. Gen. Virol. 2014, 95, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Jupatanakul, N.; Sim, S.; Dimopoulos, G. The insect microbiome modulates vector competence for arboviruses. Viruses 2014, 6, 4294–4313. [Google Scholar] [CrossRef] [PubMed]
- Dutra, H.L.; Rocha, M.N.; Dias, F.B.; Mansur, S.B.; Caragata, E.P.; Moreira, L.A. Wolbachia blocks currently circulating Zika virus isolates in Brazilian Aedes aegypti mosquitoes. Cell Host Microbe 2016, 19, 771–774. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Iturbe-Ormaetxe, I.; Callahan, A.G.; Phillips, B.L.; Billington, K.; Axford, J.K.; Montgomery, B.; Turley, A.P.; O’Neill, S.L. Stability of the wMel Wolbachia Infection following invasion into Aedes aegypti populations. PLoS Negl. Trop. Dis. 2014, 8, e3115. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbe-Ormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y.; et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Dutra, H.L.; Dos Santos, L.M.; Caragata, E.P.; Silva, J.B.; Villela, D.A.; Maciel-de-Freitas, R.; Moreira, L.A. From lab to field: The influence of urban landscapes on the invasive potential of Wolbachia in Brazilian Aedes aegypti mosquitoes. PLoS Negl. Trop. Dis. 2015, 9, e0003689. [Google Scholar] [CrossRef] [PubMed]
- Whyard, S.; Erdelyan, C.N.; Partridge, A.L.; Singh, A.D.; Beebe, N.W.; Capina, R. Silencing the buzz: A new approach to population suppression of mosquitoes by feeding larvae double-stranded RNAs. Parasit. Vectors 2015, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Taracena, M.L.; Oliveira, P.L.; Almendares, O.; Umana, C.; Lowenberger, C.; Dotson, E.M.; Paiva-Silva, G.O.; Pennington, P.M. Genetically modifying the insect gut microbiota to control Chagas disease vectors through systemic RNAi. PLoS Negl. Trop. Dis. 2015, 9, e0003358. [Google Scholar] [CrossRef] [PubMed]
- Gantz, V.M.; Jasinskiene, N.; Tatarenkova, O.; Fazekas, A.; Macias, V.M.; Bier, E.; James, A.A. Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles stephensi. Proc. Natl. Acad. Sci. USA 2015, 112, E6736–E6743. [Google Scholar] [CrossRef] [PubMed]
- Gantz, V.M.; Bier, E. Genome editing. The mutagenic chain reaction: A method for converting heterozygous to homozygous mutations. Science 2015, 348, 442–444. [Google Scholar] [CrossRef] [PubMed]
- Esvelt, K.M.; Smidler, A.L.; Catteruccia, F.; Church, G.M. Concerning RNA-guided gene drives for the alteration of wild populations. eLife 2014, 3, e03401. [Google Scholar] [CrossRef] [PubMed]
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef] [PubMed]
- Kollenberg, M.; Winter, S.; Gotz, M. Quantification and localization of watermelon chlorotic stunt virus and tomato yellow leaf curl virus (Geminiviridae) in populations of Bemisia tabaci (Hemiptera, Aleyrodidae) with differential virus transmission characteristics. PLoS ONE 2014, 9, e111968. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Li, S.J.; Ahmed, M.Z.; De Barro, P.J.; Ren, S.X.; Qiu, B.L. Inactivation of Wolbachia reveals its biological roles in whitefly host. PLoS ONE 2012, 7, e48148. [Google Scholar] [CrossRef] [PubMed]
- Augustinos, A.A.; Santos-Garcia, D.; Dionyssopoulou, E.; Moreira, M.; Papapanagiotou, A.; Scarvelakis, M.; Doudoumis, V.; Ramos, S.; Aguiar, A.F.; Borges, P.A.V.; et al. Detection and characterization of Wolbachia infections in natural populations of aphids: Is the hidden diversity fully unraveled? PLoS ONE 2011, 6, e28695. [Google Scholar] [CrossRef] [PubMed]
- De Clerck, C.; Fujiwara, A.; Joncour, P.; Leonard, S.; Felix, M.L.; Francis, F.; Jijakli, M.H.; Tsuchida, T.; Massart, S. A metagenomic approach from aphid’s hemolymph sheds light on the potential roles of co-existing endosymbionts. Microbiome 2015, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Albornoz, A.; Hoffmann, A.B.; Lozach, P.Y.; Tischler, N.D. Early bunyavirus-host cell interactions. Viruses 2016, 8, 143. [Google Scholar] [CrossRef] [PubMed]
- Bandla, M.D.; Campbell, L.R.; Ullman, D.E.; Sherwood, J.L. Interaction of tomato spotted wilt tospovirus (TSWV) glycoproteins with a thrips midgut protein, a potential cellular receptor for TSWV. Phytopathology 1998, 88, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ma, X.N.; Qian, S.S.; Zhou, X.; Sun, K.; Chen, X.L.; Zhou, X.P.; Jackson, A.O.; Li, Z.H. Rescue of a plant negative-strand RNA virus from cloned cDNA: Insights into enveloped plant virus movement and morphogenesis. PLoS Pathog. 2015, 11, e1005223. [Google Scholar] [CrossRef] [PubMed]
- Risco, C.; de Castro, I.F.; Sanz-Sanchez, L.; Narayan, K.; Grandinetti, G.; Subramaniam, S. Three-dimensional imaging of viral infections. Ann. Rev. Virol. 2014, 1, 453–473. [Google Scholar] [CrossRef] [PubMed]
- Badillo-Vargas, I.E.; Rotenberg, D.; Schneweis, B.A.; Whitfield, A.E. RNA interference tools for the western flower thrips, Frankliniella occidentalis. J. Insect Physiol. 2015, 76, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Sander, J.D.; Joung, J.K. CRISPR-Cas systems for editing, regulating and targeting genomes. Nat. Biotechnol. 2014, 32, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.J.; Zhang, P.J.; Li, W.D.; Zhang, J.M.; Huang, F.; Yang, J.; Bei, Y.W.; Lu, Y.B. De novo transcriptome sequencing in Frankliniella occidentalis to identify genes involved in plant virus transmission and insecticide resistance. Genomics 2013, 101, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Montero-Astua, M.; Rotenberg, D.; Leach-Kieffaber, A.; Schneweis, B.A.; Park, S.; Park, J.K.; German, T.L.; Whitfield, A.E. Disruption of vector transmission by a plant-expressed viral glycoprotein. Mol. Plant-Microbe Interact. 2014, 27, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Sivakumar, S.; Sparks, W.O.; Miller, W.A.; Bonning, B.C. A peptide that binds the pea aphid gut impedes entry of Pea enation mosaic virus into the aphid hemocoel. Virology 2010, 401, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Tamborindeguy, C.; Bereman, M.S.; DeBlasio, S.; Igwe, D.; Smith, D.M.; White, F.; MacCoss, M.J.; Gray, S.M.; Cilia, M. Genomic and proteomic analysis of Schizaphis graminum reveals cyclophilin proteins are involved in the transmission of Cereal yellow dwarf virus. PLoS ONE 2013, 8, e71620. [Google Scholar] [CrossRef] [PubMed]
- Bonning, B.C.; Pal, N.; Liu, S.J.; Wang, Z.H.; Sivakumar, S.; Dixon, P.M.; King, G.F.; Miller, W.A. Toxin delivery by the coat protein of an aphid-vectored plant virus provides plant resistance to aphids. Nat. Biotechnol. 2014, 32, 102–105. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dietzgen, R.G.; Mann, K.S.; Johnson, K.N. Plant Virus–Insect Vector Interactions: Current and Potential Future Research Directions. Viruses 2016, 8, 303. https://doi.org/10.3390/v8110303
Dietzgen RG, Mann KS, Johnson KN. Plant Virus–Insect Vector Interactions: Current and Potential Future Research Directions. Viruses. 2016; 8(11):303. https://doi.org/10.3390/v8110303
Chicago/Turabian StyleDietzgen, Ralf G., Krin S. Mann, and Karyn N. Johnson. 2016. "Plant Virus–Insect Vector Interactions: Current and Potential Future Research Directions" Viruses 8, no. 11: 303. https://doi.org/10.3390/v8110303
APA StyleDietzgen, R. G., Mann, K. S., & Johnson, K. N. (2016). Plant Virus–Insect Vector Interactions: Current and Potential Future Research Directions. Viruses, 8(11), 303. https://doi.org/10.3390/v8110303