
Dengue Virus Reporter Replicon is a Valuable Tool for Antiviral Drug Discovery and Analysis of Virus Replication Mechanisms

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
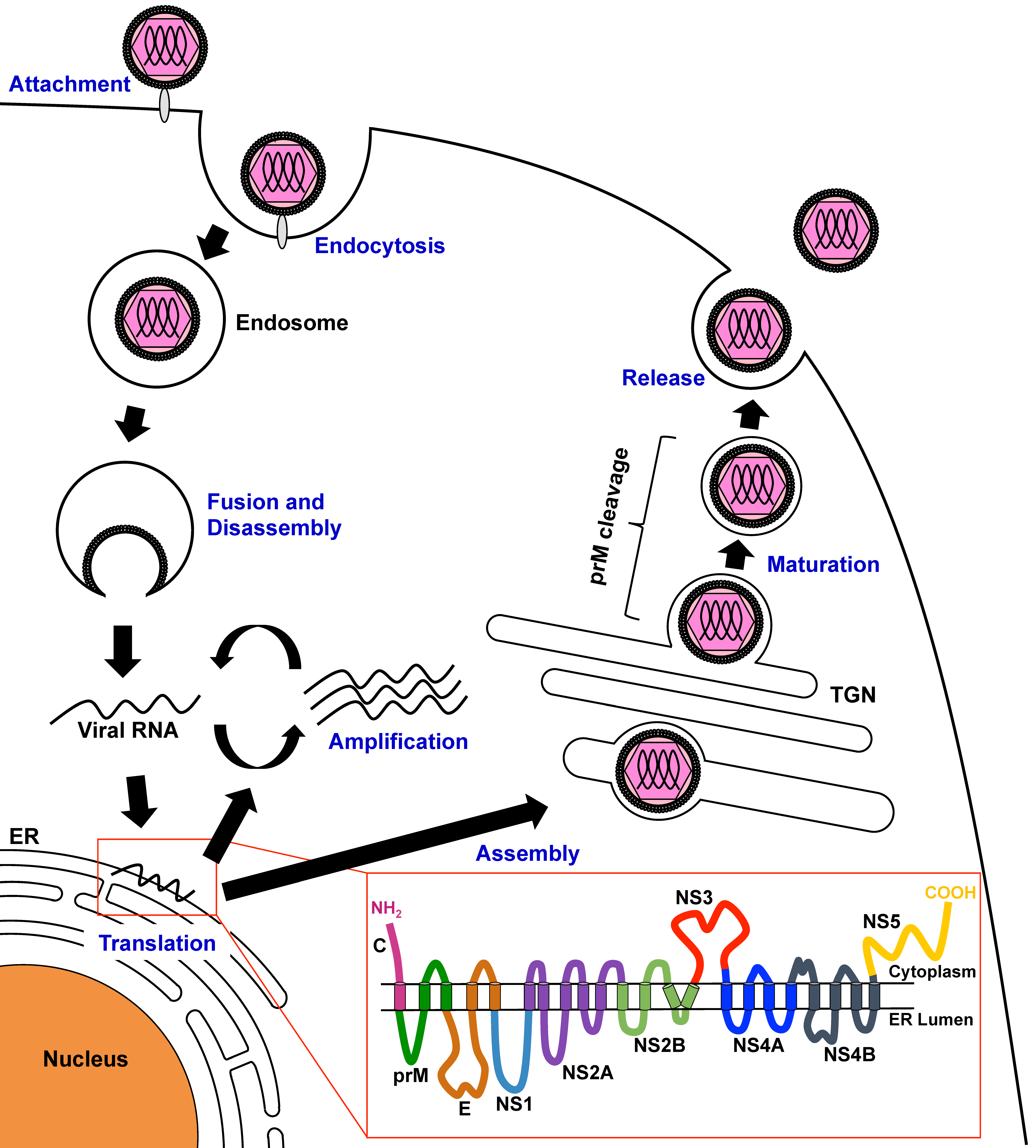
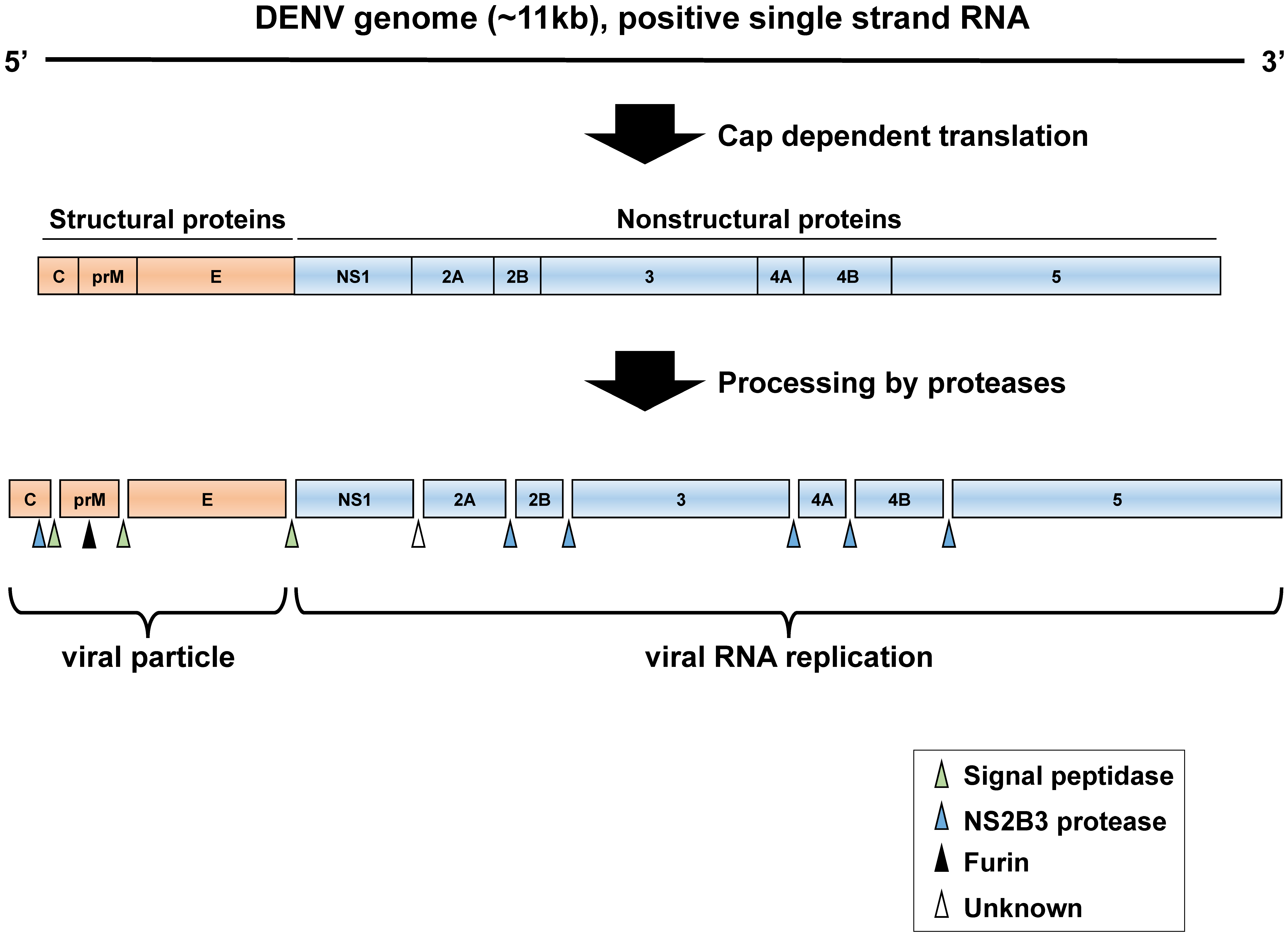
2. DENV Life Cycle
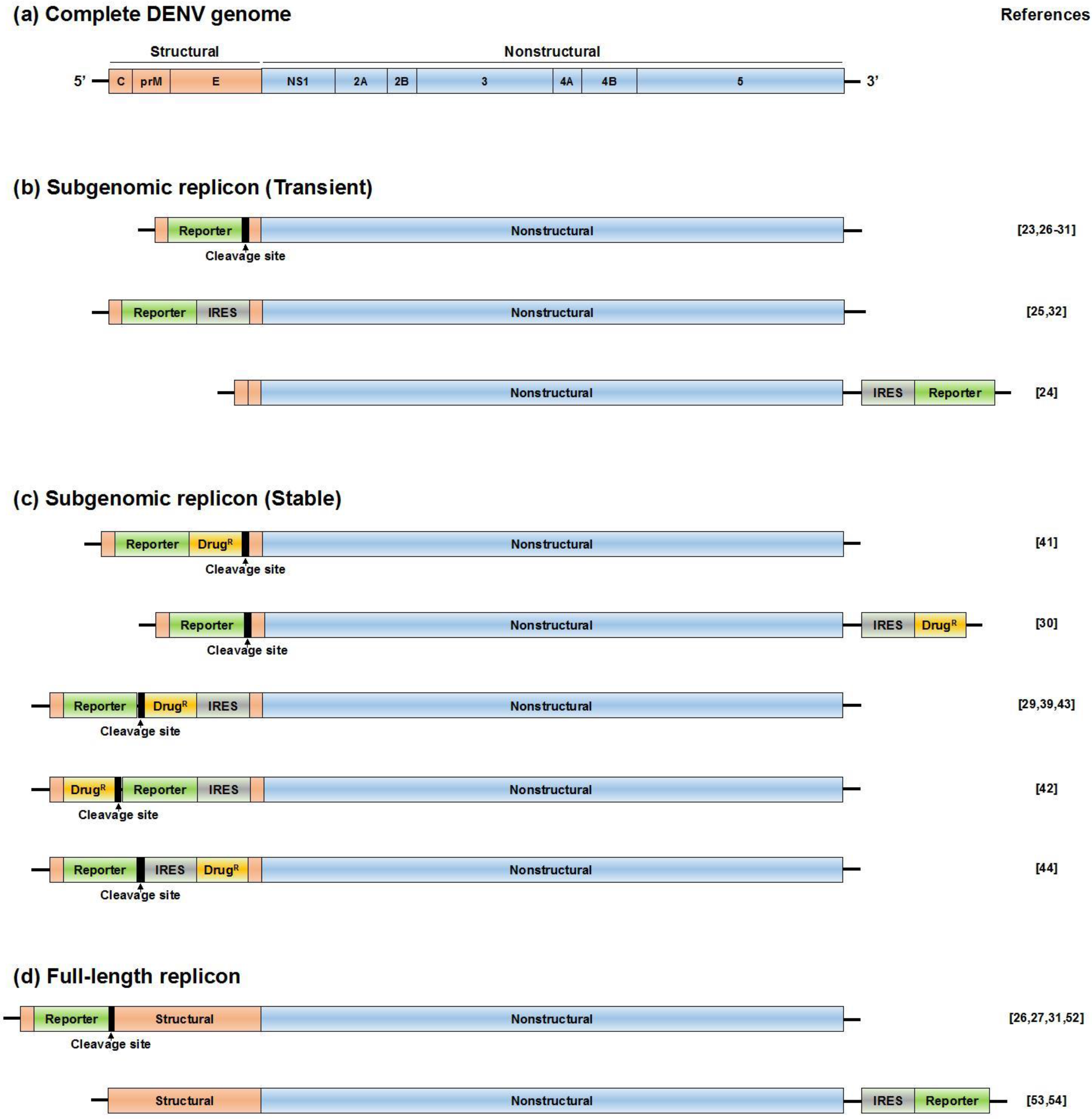
3. Subgenomic Reporter Replicon (Transient Expression of NS Proteins)
4. Subgenomic Reporter Replicon Cells (Stable Expression of NS Proteins)
5. Virus-like Particles
6. Full-Length Reporter Replicon
7. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gubler, D.J. Dengue and dengue hemorrhagic fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [PubMed]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Karabatsos, N.; Dalrymple, J.M.; Shope, R.E.; Porterfield, J.S.; Westaway, E.G.; Brandt, W.E. Antigenic relationships between flaviviruses as determined by cross-neutralization tests with polyclonal antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus genome organization, expression, and replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, D.E.; Lodeiro, M.F.; Luduena, S.J.; Pietrasanta, L.I.; Gamarnik, A.V. Long-range rna-rna interactions circularize the dengue virus genome. J. Virol. 2005, 79, 6631–6643. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, D.E.; Filomatori, C.V.; Gamarnik, A.V. Functional analysis of dengue virus cyclization sequences located at the 5′ and 3′utrs. Virology 2008, 375, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Friebe, P.; Harris, E. Interplay of rna elements in the dengue virus 5′ and 3′ ends required for viral rna replication. J. Virol. 2010, 84, 6103–6118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y.; Li, X.F.; Jiang, T.; Deng, Y.Q.; Zhao, H.; Wang, H.J.; Ye, Q.; Zhu, S.Y.; Qiu, Y.; Zhou, X.; et al. Novel cis-acting element within the capsid-coding region enhances flavivirus viral-rna replication by regulating genome cyclization. J. Virol. 2013, 87, 6804–6818. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Levis, R.; Shen, P.; Schlesinger, S.; Rice, C.M.; Huang, H.V. Sindbis virus: An efficient, broad host range vector for gene expression in animal cells. Science 1989, 243, 1188–1191. [Google Scholar] [CrossRef] [PubMed]
- Hagino-Yamagishi, K.; Nomoto, A. In vitro construction of poliovirus defective interfering particles. J. Virol. 1989, 63, 5386–5392. [Google Scholar] [PubMed]
- Liljestrom, P.; Garoff, H. A new generation of animal cell expression vectors based on the semliki forest virus replicon. Biotechnology (N. Y.) 1991, 9, 1356–1361. [Google Scholar] [CrossRef]
- McKnight, K.L.; Lemon, S.M. Capsid coding sequence is required for efficient replication of human rhinovirus 14 rna. J. Virol. 1996, 70, 1941–1952. [Google Scholar] [PubMed]
- Almazan, F.; Sola, I.; Zuniga, S.; Marquez-Jurado, S.; Morales, L.; Becares, M.; Enjuanes, L. Coronavirus reverse genetic systems: Infectious clones and replicons. Virus Res. 2014, 189, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, V.; Korner, F.; Koch, J.; Herian, U.; Theilmann, L.; Bartenschlager, R. Replication of subgenomic hepatitis c virus rnas in a hepatoma cell line. Science 1999, 285, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Khromykh, A.A.; Westaway, E.G. Subgenomic replicons of the flavivirus kunjin: Construction and applications. J. Virol. 1997, 71, 1497–1505. [Google Scholar] [PubMed]
- Pang, X.; Zhang, M.; Dayton, A.I. Development of dengue virus type 2 replicons capable of prolonged expression in host cells. BMC Microbiol. 2001, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.Y.; Tilgner, M.; Lo, M.K. Construction and characterization of subgenomic replicons of New York strain of west nile virus. Virology 2002, 296, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Corver, J.; Lenches, E.; Smith, K.; Robison, R.A.; Sando, T.; Strauss, E.G.; Strauss, J.H. Fine mapping of a cis-acting sequence element in yellow fever virus rna that is required for rna replication and cyclization. J. Virol. 2003, 77, 2265–2270. [Google Scholar] [CrossRef] [PubMed]
- Gehrke, R.; Ecker, M.; Aberle, S.W.; Allison, S.L.; Heinz, F.X.; Mandl, C.W. Incorporation of tick-borne encephalitis virus replicons into virus-like particles by a packaging cell line. J. Virol. 2003, 77, 8924–8933. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Oliveira, C.; Freire, J.M.; Conceicao, T.M.; Higa, L.M.; Castanho, M.A.; da Poian, A.T. Receptors and routes of dengue virus entry into the host cells. FEMS Microbiol. Rev. 2015, 39, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Teo, C.S.; Chu, J.J. Cellular vimentin regulates construction of dengue virus replication complexes through interaction with ns4a protein. J. Virol. 2014, 88, 1897–1913. [Google Scholar] [CrossRef] [PubMed]
- Welsch, S.; Miller, S.; Romero-Brey, I.; Merz, A.; Bleck, C.K.; Walther, P.; Fuller, S.D.; Antony, C.; Krijnse-Locker, J.; Bartenschlager, R. Composition and three-dimensional architecture of the dengue virus replication and assembly sites. Cell Host Microbe 2009, 5, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Holden, K.L.; Stein, D.A.; Pierson, T.C.; Ahmed, A.A.; Clyde, K.; Iversen, P.L.; Harris, E. Inhibition of dengue virus translation and rna synthesis by a morpholino oligomer targeted to the top of the terminal 3′ stem-loop structure. Virology 2006, 344, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; de Borba, L.; Duarte dos Santos, C.N.; Mason, P.W. Construction of an infectious cdna clone for a brazilian prototype strain of dengue virus type 1: Characterization of a temperature-sensitive mutation in ns1. Virology 2007, 362, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz-Estrada, S.L.; Manzano, M.I.; del Angel, R.M.; Levis, R.; Padmanabhan, R. Construction of a dengue virus type 4 reporter replicon and analysis of temperature-sensitive mutations in non-structural proteins 3 and 5. J. Gen. Virol. 2010, 91, 2713–2718. [Google Scholar] [CrossRef] [PubMed]
- Scaturro, P.; Trist, I.M.; Paul, D.; Kumar, A.; Acosta, E.G.; Byrd, C.M.; Jordan, R.; Brancale, A.; Bartenschlager, R. Characterization of the mode of action of a potent dengue virus capsid inhibitor. J. Virol. 2014, 88, 11540–11555. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.; Xu, H.Y.; Qing, M.; Wang, Q.Y.; Shi, P.Y. Development and characterization of a stable luciferase dengue virus for high-throughput screening. Antivir. Res. 2011, 91, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, D.E.; de Lella Ezcurra, A.L.; Fucito, S.; Gamarnik, A.V. Role of rna structures present at the 3′utr of dengue virus on translation, rna synthesis, and viral replication. Virology 2005, 339, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Puig-Basagoiti, F.; Tilgner, M.; Forshey, B.M.; Philpott, S.M.; Espina, N.G.; Wentworth, D.E.; Goebel, S.J.; Masters, P.S.; Falgout, B.; Ren, P.; et al. Triaryl pyrazoline compound inhibits flavivirus rna replication. Antimicrob. Agents Chemother. 2006, 50, 1320–1329. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.C.; Chen, N.C.; Chen, P.C.; Wang, C.C.; Cheng, W.C.; Wu, H.N. Identification of a small-molecule inhibitor of dengue virus using a replicon system. Arch. Virol. 2012, 157, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Leardkamolkarn, V.; Sirigulpanit, W.; Chotiwan, N.; Kumkate, S.; Huang, C.Y. Development of dengue type-2 virus replicons expressing gfp reporter gene in study of viral rna replication. Virus Res. 2012, 163, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Kato, F.; Kobayashi, T.; Tajima, S.; Takasaki, T.; Miura, T.; Igarashi, T.; Hishiki, T. Development of a novel dengue-1 virus replicon system expressing secretory gaussia luciferase for analysis of viral replication and discovery of antiviral drugs. Jpn. J. Infect. Dis. 2014, 67, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Khromykh, A.A.; Meka, H.; Guyatt, K.J.; Westaway, E.G. Essential role of cyclization sequences in flavivirus rna replication. J. Virol. 2001, 75, 6719–6728. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Falgout, B.; Markoff, L.; Padmanabhan, R. In vitro rna synthesis from exogenous dengue viral rna templates requires long range interactions between 5′- and 3′-terminal regions that influence rna structure. J. Biol. Chem. 2001, 276, 15581–15591. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.D.; Drew, J. Foot-and-mouth disease virus 2a oligopeptide mediated cleavage of an artificial polyprotein. EMBO J. 1994, 13, 928–933. [Google Scholar] [PubMed]
- Ljungberg, K.; Whitmore, A.C.; Fluet, M.E.; Moran, T.P.; Shabman, R.S.; Collier, M.L.; Kraus, A.A.; Thompson, J.M.; Montefiori, D.C.; Beard, C.; et al. Increased immunogenicity of a DNA-launched venezuelan equine encephalitis virus-based replicon DNA vaccine. J. Virol. 2007, 81, 13412–13423. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Yao, Q.; Fan, H.; Xiao, S.; Si, Y.; Chen, H. Development of a vaccine vector based on a subgenomic replicon of porcine reproductive and respiratory syndrome virus. J. Virol. Methods 2009, 160, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Li, X.F.; Yu, X.D.; Deng, Y.Q.; Jiang, T.; Zhu, Q.Y.; Qin, E.D.; Qin, C.F. A DNA-based west nile virus replicon elicits humoral and cellular immune responses in mice. J. Virol. Methods 2011, 178, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Tsai, M.H.; Hu, H.S.; Pu, S.Y.; Wu, R.H.; Wu, S.H.; Lin, H.M.; Song, J.S.; Chao, Y.S.; Yueh, A. Characterization of an efficient dengue virus replicon for development of assays of discovery of small molecules against dengue virus. Antivir. Res. 2013, 98, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Tannous, B.A.; Kim, D.E.; Fernandez, J.L.; Weissleder, R.; Breakefield, X.O. Codon-optimized gaussia luciferase cdna for mammalian gene expression in culture and in vivo. Mol. Ther. 2005, 11, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Masse, N.; Davidson, A.; Ferron, F.; Alvarez, K.; Jacobs, M.; Romette, J.L.; Canard, B.; Guillemot, J.C. Dengue virus replicons: Production of an interserotypic chimera and cell lines from different species, and establishment of a cell-based fluorescent assay to screen inhibitors, validated by the evaluation of ribavirin’s activity. Antivir. Res. 2010, 86, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.; Gu, F.; Phong, W.Y.; Chen, Y.L.; Lim, S.P.; Davidson, A.; Vasudevan, S.G. Construction and characterization of a stable subgenomic dengue virus type 2 replicon system for antiviral compound and sirna testing. Antivir. Res. 2007, 76, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Hsieh, Y.C.; Lee, S.J.; Wu, S.H.; Liao, C.L.; Tsao, C.H.; Chao, Y.S.; Chern, J.H.; Wu, C.P.; Yueh, A. Novel dengue virus-specific ns2b/ns3 protease inhibitor, bp2109, discovered by a high-throughput screening assay. Antimicrob. Agents Chemother. 2011, 55, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Leardkamolkarn, V.; Sirigulpanit, W. Establishment of a stable cell line coexpressing dengue virus-2 and green fluorescent protein for screening of antiviral compounds. J. Biomol. Screen 2012, 17, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.T.; Patkar, C.G.; Kuhn, R.J. Construction and applications of yellow fever virus replicons. Virology 2005, 331, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.W.; Nguyen, H.Y.; Hanna, S.L.; Sanchez, M.D.; Doms, R.W.; Pierson, T.C. West nile virus discriminates between dc-sign and dc-signr for cellular attachment and infection. J. Virol. 2006, 80, 1290–1301. [Google Scholar] [CrossRef] [PubMed]
- Qing, M.; Liu, W.; Yuan, Z.; Gu, F.; Shi, P.Y. A high-throughput assay using dengue-1 virus-like particles for drug discovery. Antivir. Res. 2010, 86, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Klasse, P.J.; Sattentau, Q.J. Mechanisms of virus neutralization by antibody. Curr. Top. Microbiol. Immunol. 2001, 260, 87–108. [Google Scholar] [PubMed]
- Yin, Z.; Chen, Y.L.; Schul, W.; Wang, Q.Y.; Gu, F.; Duraiswamy, J.; Kondreddi, R.R.; Niyomrattanakit, P.; Lakshminarayana, S.B.; Goh, A.; et al. An adenosine nucleoside inhibitor of dengue virus. Proc. Natl. Acad. Sci. USA 2009, 106, 20435–20439. [Google Scholar] [CrossRef] [PubMed]
- Ansarah-Sobrinho, C.; Nelson, S.; Jost, C.A.; Whitehead, S.S.; Pierson, T.C. Temperature-dependent production of pseudoinfectious dengue reporter virus particles by complementation. Virology 2008, 381, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Mattia, K.; Puffer, B.A.; Williams, K.L.; Gonzalez, R.; Murray, M.; Sluzas, E.; Pagano, D.; Ajith, S.; Bower, M.; Berdougo, E.; et al. Dengue reporter virus particles for measuring neutralizing antibodies against each of the four dengue serotypes. PLoS ONE 2011, 6, e27252. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; Dorner, M.; Feulner, M.; Imanaka, N.; Murphy, M.Y.; Ploss, A.; Rice, C.M. Dengue reporter viruses reveal viral dynamics in interferon receptor-deficient mice and sensitivity to interferon effectors in vitro. Proc. Natl. Acad. Sci. USA 2012, 109, 14610–14615. [Google Scholar] [CrossRef] [PubMed]
- Mondotte, J.A.; Lozach, P.Y.; Amara, A.; Gamarnik, A.V. Essential role of dengue virus envelope protein n glycosylation at asparagine-67 during viral propagation. J. Virol. 2007, 81, 7136–7148. [Google Scholar] [CrossRef] [PubMed]
- Kaptein, S.J.; de Burghgraeve, T.; Froeyen, M.; Pastorino, B.; Alen, M.M.; Mondotte, J.A.; Herdewijn, P.; Jacobs, M.; de Lamballerie, X.; Schols, D.; et al. A derivate of the antibiotic doxorubicin is a selective inhibitor of dengue and yellow fever virus replication in vitro. Antimicrob. Agents Chemother. 2010, 54, 5269–5280. [Google Scholar] [CrossRef] [PubMed]
- Deas, T.S.; Binduga-Gajewska, I.; Tilgner, M.; Ren, P.; Stein, D.A.; Moulton, H.M.; Iversen, P.L.; Kauffman, E.B.; Kramer, L.D.; Shi, P.Y. Inhibition of flavivirus infections by antisense oligomers specifically suppressing viral translation and rna replication. J. Virol. 2005, 79, 4599–4609. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Zachariah, M.; Harris, E. Mycophenolic acid inhibits dengue virus infection by preventing replication of viral rna. Virology 2002, 304, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.; Tan, G.K.; Qing, M.; Ng, J.K.; Yip, A.; Zou, G.; Xie, X.; Yuan, Z.; Schreiber, M.J.; Schul, W.; et al. A single amino acid in nonstructural protein ns4b confers virulence to dengue virus in ag129 mice through enhancement of viral rna synthesis. J. Virol. 2011, 85, 7775–7787. [Google Scholar] [CrossRef] [PubMed]
- Kyle, J.L.; Beatty, P.R.; Harris, E. Dengue virus infects macrophages and dendritic cells in a mouse model of infection. J. Infect. Dis. 2007, 195, 1808–1817. [Google Scholar] [CrossRef] [PubMed]
- Balsitis, S.J.; Coloma, J.; Castro, G.; Alava, A.; Flores, D.; McKerrow, J.H.; Beatty, P.R.; Harris, E. Tropism of dengue virus in mice and humans defined by viral nonstructural protein 3-specific immunostaining. Am. J. Trop. Med. Hyg. 2009, 80, 416–424. [Google Scholar] [PubMed]
- Zellweger, R.M.; Prestwood, T.R.; Shresta, S. Enhanced infection of liver sinusoidal endothelial cells in a mouse model of antibody-induced severe dengue disease. Cell Host Microbe 2010, 7, 128–139. [Google Scholar] [CrossRef] [PubMed]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato, F.; Hishiki, T. Dengue Virus Reporter Replicon is a Valuable Tool for Antiviral Drug Discovery and Analysis of Virus Replication Mechanisms. Viruses 2016, 8, 122. https://doi.org/10.3390/v8050122
Kato F, Hishiki T. Dengue Virus Reporter Replicon is a Valuable Tool for Antiviral Drug Discovery and Analysis of Virus Replication Mechanisms. Viruses. 2016; 8(5):122. https://doi.org/10.3390/v8050122
Chicago/Turabian StyleKato, Fumihiro, and Takayuki Hishiki. 2016. "Dengue Virus Reporter Replicon is a Valuable Tool for Antiviral Drug Discovery and Analysis of Virus Replication Mechanisms" Viruses 8, no. 5: 122. https://doi.org/10.3390/v8050122
APA StyleKato, F., & Hishiki, T. (2016). Dengue Virus Reporter Replicon is a Valuable Tool for Antiviral Drug Discovery and Analysis of Virus Replication Mechanisms. Viruses, 8(5), 122. https://doi.org/10.3390/v8050122