
Cross-Regulation between Transposable Elements and Host DNA Replication


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
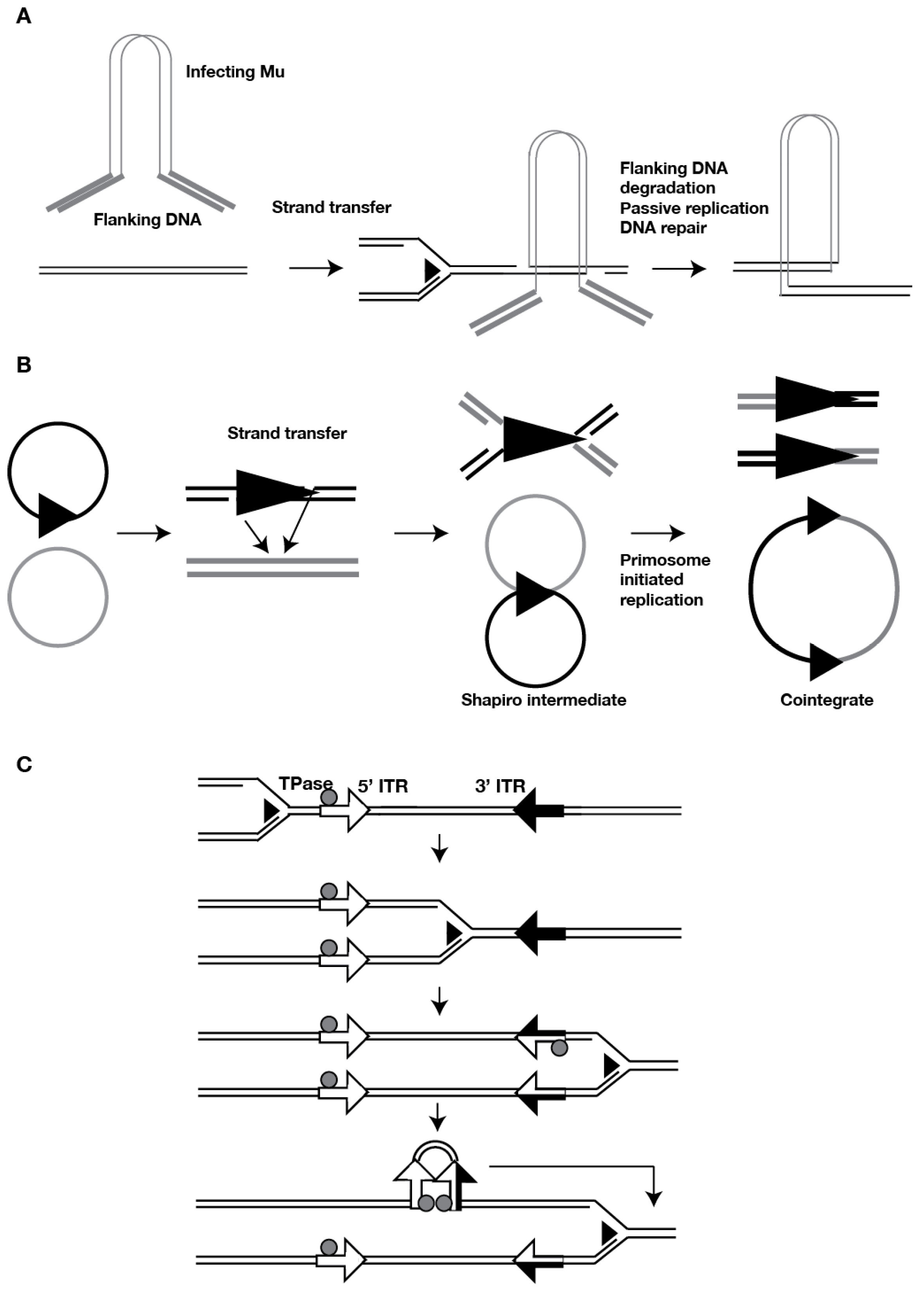
2. DNA Transposon Duplication
3. Role of the Replication Fork in Transposon Target-Site Selection
4. Influence of TEs Presence in Host DNA Replication and Homologous Recombination
Acknowledgments
Conflicts of Interest
References
- Winston, F.; Chaleff, D.T.; Valent, B.; Fink, G.R. Mutations affecting Ty-mediated expression of the HIS4 gene of Saccharomyces cerevisiae. Genetics 1984, 107, 179–197. [Google Scholar] [PubMed]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A. Molecular model for the transposition and replication of bacteriophage Mu and other transposable elements. Proc. Natl. Acad. Sci. USA 1979, 76, 1933–1937. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Pritham, E.J. Helitrons, the Eukaryotic Rolling-circle Transposable Elements. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Harshey, R.M. Repair of transposable phage Mu DNA insertions begins only when the E. coli replisome collides with the transpososome. Mol. Microbiol. 2015, 97, 746–758. [Google Scholar] [CrossRef] [PubMed]
- Nakai, H.; Doseeva, V.; Jones, J.M. Handoff from recombinase to replisome: Insights from transposition. Proc. Natl. Acad. Sci. USA 2001, 98, 8247–8254. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, I.M.; Brink, R.A. Twin Mutations in Medium Variegated Pericarp Maize. Genetics 1962, 47, 489–501. [Google Scholar] [PubMed]
- Greenblatt, I.M. A chromosome replication pattern deduced from pericarp phenotypes resulting from movements of the transposable element, modulator, in maize. Genetics 1984, 108, 471–485. [Google Scholar] [PubMed]
- Chen, J.; Greenblatt, I.M.; Dellaporta, S.L. Transposition of Ac from the P locus of maize into unreplicated chromosomal sites. Genetics 1987, 117, 109–116. [Google Scholar] [PubMed]
- Ros, F.; Kunze, R. Regulation of activator/dissociation transposition by replication and DNA methylation. Genetics 2001, 157, 1723–1733. [Google Scholar]
- Kunze, R.; Starlinger, P. The putative transposase of transposable element Ac from Zea mays L. interacts with subterminal sequences of Ac. EMBO J. 1989, 8, 3177–3185. [Google Scholar] [PubMed]
- Wang, L.; Heinlein, M.; Kunze, R. Methylation pattern of Activator transposase binding sites in maize endosperm. Plant Cell 1996, 8, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.; Hoopes, B.C.; McClure, W.R.; Kleckner, N. IS10 transposition is regulated by DNA adenine methylation. Cell 1985, 43, 117–130. [Google Scholar] [CrossRef]
- Claeys Bouuaert, C.; Liu, D.; Chalmers, R. A simple topological filter in a eukaryotic transposon as a mechanism to suppress genome instability. Mol. Cell. Biol. 2010, 31, 3925–3932. [Google Scholar] [CrossRef] [PubMed]
- Claeys Bouuaert, C.; Chalmers, R. Hsmar1 transposition is sensitive to the topology of the transposon donor and the target. PLoS ONE 2013, 8, e53690. [Google Scholar] [CrossRef] [PubMed]
- Saredi, G.; Huang, H.; Hammond, C.M.; Alabert, C.; Bekker-Jensen, S.; Forne, I.; Reverón-Gómez, N.; Foster, B.M.; Mlejnkova, L.; Bartke, T.; et al. H4K20me0 marks post-replicative chromatin and recruits the TONSL–MMS22L DNA repair complex. Nature 2016, 534, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, Y.; Inoue, K.; Oikawa, M.; Kamimura, S.; Ogonuki, N.; Kodama, E.N.; Ohkawa, Y.; Tsukada, Y.-I.; Ogura, A. Histone chaperone CAF-1 mediates repressive histone modifications to protect preimplantation mouse embryos from endogenous retrotransposons. Proc. Natl. Acad. Sci. USA 2015, 112, 14641–14646. [Google Scholar] [CrossRef] [PubMed]
- Bloom, L.B. Loading clamps for DNA replication and repair. DNA Repair (Amst.) 2009, 8, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Warbrick, E.; Heatherington, W.; Lane, D.P.; Glover, D.M. PCNA binding proteins in Drosophila melanogaster: The analysis of a conserved PCNA binding domain. Nucleic Acids Res. 1998, 26, 3925–3932. [Google Scholar] [CrossRef] [PubMed]
- Warbrick, E. The puzzle of PCNA’s many partners. Bioessays 2000, 22, 997–1006. [Google Scholar] [CrossRef]
- Parks, A.R.; Li, Z.; Shi, Q.; Owens, R.M.; Jin, M.M.; Peters, J.E. Transposition into replicating DNA occurs through interaction with the processivity factor. Cell 2009, 138, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Waddell, C.S.; Craig, N.L. Tn7 transposition: Two transposition pathways directed by five Tn7-encoded genes. Genes Dev. 1988, 2, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Waddell, C.S.; Craig, N.L. Tn7 transposition: Recognition of the attTn7 target sequence. Proc. Natl. Acad. Sci. USA 1989, 86, 3958–3962. [Google Scholar] [CrossRef] [PubMed]
- Wolkow, C.A.; DeBoy, R.T.; Craig, N.L. Conjugating plasmids are preferred targets for Tn7. Genes Dev. 1996, 10, 2145–2157. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.E.; Craig, N.L. Tn7 transposes proximal to DNA double-strand breaks and into regions where chromosomal DNA replication terminates. Mol. Cell 2000, 6, 573–582. [Google Scholar] [CrossRef]
- Peters, J.E.; Craig, N.L. Tn7 recognizes transposition target structures associated with DNA replication using the DNA-binding protein TnsE. Genes Dev. 2001, 15, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Fricker, A.D.; Peters, J.E. Vulnerabilities on the Lagging-Strand Template: Opportunities for Mobile Elements. Annu. Rev. Genet. 2014, 48, 167–186. [Google Scholar] [CrossRef] [PubMed]
- Ton-Hoang, B.; Guynet, C.; Ronning, D.R.; Cointin-Marty, B.; Dyda, F.; Chandler, M. Transposition of ISHp608, member of an unusual family of bacterial insertion sequences. EMBO J. 2005, 24, 3325–3338. [Google Scholar] [CrossRef] [PubMed]
- Guynet, C.; Hickman, A.B.; Barabas, O.; Dyda, F.; Chandler, M.; Ton-Hoang, B. In vitro reconstitution of a single-stranded transposition mechanism of IS608. Mol. Cell 2008, 29, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Ton-Hoang, B.; Pasternak, C.; Siguier, P.; Guynet, C.; Hickman, A.B.; Dyda, F.; Sommer, S.; Chandler, M. Single-stranded DNA transposition is coupled to host replication. Cell 2010, 142, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Lavatine, L.; He, S.; Caumont-Sarcos, A.; Guynet, C.; Marty, B.; Chandler, M.; Ton-Hoang, B. Single strand transposition at the host replication fork. Nucleic Acids Res. 2016, 44, 7866–7883. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.J.; Díaz-Maldonado, H.; González-Tortuero, E.; López de Saro, F.J. Chromosomal replication dynamics and interaction with the β sliding clamp determine orientation of bacterial transposable elements. Genome Biol. Evol. 2014, 6, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Mularoni, L.; Zhou, Y.; Bowen, T.; Gangadharan, S.; Wheelan, S.J.; Boeke, J.D. Retrotransposon Ty1 integration targets specifically positioned asymmetric nucleosomal DNA segments in tRNA hotspots. Genome Res. 2012, 22, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Daily, K.; Nguyen, K.; Wang, H.; Mayhew, D.; Rigor, P.; Forouzan, S.; Johnston, M.; Mitra, R.D.; Baldi, P.; et al. Retrotransposon profiling of RNA polymerase III initiation sites. Genome Res. 2012, 22, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Szilard, R.K.; Jacques, P.-E.; Laramée, L.; Cheng, B.; Galicia, S.; Bataille, A.R.; Yeung, M.; Mendez, M.; Bergeron, M.; Robert, F.; et al. Systematic identification of fragile sites via genome-wide location analysis of gamma-H2AX. Nat. Struct. Mol. Biol. 2010, 17, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.Z.; Rosado-Lugo, J.D.; Cranz-Mileva, S.; Ciccaglione, K.M.; Tournier, V.; Zaratiegui, M. Arrested replication forks guide retrotransposon integration. Science 2015, 349, 1549–1553. [Google Scholar] [CrossRef] [PubMed]
- Sabouri, N.; Capra, J.A.; Zakian, V.A. The essential Schizosaccharomyces pombe Pfh1 DNA helicase promotes fork movement past G-quadruplex motifs to prevent DNA damage. BMC Biol. 2014, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Zaratiegui, M.; Vaughn, M.W.; Irvine, D.V.; Goto, D.; Watt, S.; Bähler, J.; Arcangioli, B.; Martienssen, R.A. CENP-B preserves genome integrity at replication forks paused by retrotransposon LTR. Nature 2011, 469, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.S.; LaCava, J.; Mita, P.; Molloy, K.R.; Huang, C.R. L.; Li, D.; Adney, E.M.; Jiang, H.; Burns, K.H.; Chait, B.T.; et al. Affinity proteomics reveals human host factors implicated in discrete stages of LINE-1 retrotransposition. Cell 2013, 155, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, R.; Lai, M.S.; Foiani, M. Preventing replication stress to maintain genome stability: Resolving conflicts between replication and transcription. Mol. Cell 2012, 45, 710–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, S.; Carr, A.M. Replication stress and genome rearrangements: Lessons from yeast models. Curr. Opin. Genet. Dev. 2013, 23, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Labib, K.; Hodgson, B. Replication fork barriers: Pausing for a break or stalling for time? EMBO Rep. 2007, 8, 346–353. [Google Scholar] [CrossRef] [PubMed]
- García-Muse, T.; Aguilera, A. Transcription-replication conflicts: How they occur and how they are resolved. Nat. Rev. Mol. Cell Biol. 2016, 17, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.L.; Bastia, D. Mechanisms of polar arrest of a replication fork. Mol. Microbiol. 2009, 72, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Sabouri, N. The functions of the multi-tasking Pfh1(Pif1) helicase. Curr. Genet. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.; Froget, B.; Carr, A.M. Arrested replication fork processing: Interplay between checkpoints and recombination. DNA Repair (Amst.) 2007, 6, 1042–1061. [Google Scholar] [CrossRef] [PubMed]
- Bushman, F.D. Targeting survival: Integration site selection by retroviruses and LTR-retrotransposons. Cell 2003, 115, 135–138. [Google Scholar] [CrossRef]
- Bridier-Nahmias, A.; Tchalikian-Cosson, A.; Baller, J.A.; Menouni, R.; Fayol, H.; Flores, A.; Saïb, A.; Werner, M.; Voytas, D.F.; Lesage, P. An RNA polymerase III subunit determines sites of retrotransposon integration. Science 2015, 348, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, J.; Connolly, C.M.; Sandmeyer, S.B. Requirement of RNA polymerase III transcription factors for in vitro position-specific integration of a retroviruslike element. Science 1995, 267, 1488–1491. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Gai, X.; Zhu, Y.; Zappulla, D.C.; Sternglanz, R.; Voytas, D.F. Targeting of the yeast Ty5 retrotransposon to silent chromatin is mediated by interactions between integrase and Sir4p. Mol. Cell. Biol. 2001, 21, 6606–6614. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Dai, J.; Fuerst, P.G.; Voytas, D.F. Controlling integration specificity of a yeast retrotransposon. Proc. Natl. Acad. Sci. USA 2003, 100, 5891–5895. [Google Scholar] [CrossRef] [PubMed]
- Bowen, N.J.; Jordan, I.K.; Epstein, J.A.; Wood, V.; Levin, H.L. Retrotransposons and their recognition of pol II promoters: A comprehensive survey of the transposable elements from the complete genome sequence of Schizosaccharomyces pombe. Genome Res. 2003, 13, 1984–1997. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Levin, H.L. High-throughput sequencing of retrotransposon integration provides a saturated profile of target activity in Schizosaccharomyces pombe. Genome Res. 2010, 20, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, A.; Chatterjee, A.G.; Ripmaster, T.L.; Levin, H.L. Determinants that specify the integration pattern of retrotransposon Tf1 in the fbp1 promoter of Schizosaccharomyces pombe. J. Virol. 2011, 85, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Tsankov, A.; Yanagisawa, Y.; Rhind, N.; Regev, A.; Rando, O.J. Evolutionary divergence of intrinsic and trans-regulated nucleosome positioning sequences reveals plastic rules for chromatin organization. Genome Res. 2011, 21, 1851–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, A.; Esnault, C.; Majumdar, A.; Chatterjee, A.G.; Iben, J.R.; McQueen, P.G.; Yang, A.X.; Mizuguchi, T.; Grewal, S.I.S.; Levin, H.L. Single-Nucleotide-Specific Targeting of the Tf1 Retrotransposon Promoted by the DNA-Binding Protein Sap1 of Schizosaccharomyces pombe. Genetics 2015, 201, 905–924. [Google Scholar] [CrossRef] [PubMed]
- Mejía-Ramírez, E.; Sánchez-Gorostiaga, A.; Krimer, D.B.; Schvartzman, J.B.; Hernández, P. The mating type switch-activating protein Sap1 Is required for replication fork arrest at the rRNA genes of fission yeast. Mol. Cell. Biol. 2005, 25, 8755–8761. [Google Scholar] [CrossRef] [PubMed]
- Krings, G.; Bastia, D. Sap1p binds to Ter1 at the ribosomal DNA of Schizosaccharomyces pombe and causes polar replication fork arrest. J. Biol. Chem. 2005, 280, 39135–39142. [Google Scholar] [CrossRef] [PubMed]
- Krings, G.; Bastia, D. Molecular architecture of a eukaryotic DNA replication terminus-terminator protein complex. Mol. Cell. Biol. 2006, 26, 8061–8074. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.M.; Newlon, C.S. DNA replication fork pause sites dependent on transcription. Science 1996, 272, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Sabouri, N.; McDonald, K.R.; Webb, C.J.; Cristea, I.M.; Zakian, V.A. DNA replication through hard-to-replicate sites, including both highly transcribed RNA Pol II and Pol III genes, requires the S. pombe Pfh1 helicase. Genes Dev. 2012, 26, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Dubarry, M.; Loiodice, I.; Chen, C.L.; Thermes, C.; Taddei, A. Tight protein-DNA interactions favor gene silencing. Genes Dev. 2011, 25, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Spaller, T.; Kling, E.; Glöckner, G.; Hillmann, F.; Winckler, T. Convergent evolution of tRNA gene targeting preferences in compact genomes. Mob. DNA 2016, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Singh, P.K.; Levin, H.L. A long terminal repeat retrotransposon of Schizosaccharomyces japonicus integrates upstream of RNA pol III transcribed genes. Mob. DNA 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Rhind, N.; Chen, Z.; Yassour, M.; Thompson, D.A.; Haas, B.J.; Habib, N.; Wapinski, I.; Roy, S.; Lin, M.F.; Heiman, D.I.; et al. Comparative Functional Genomics of the Fission Yeasts. Science 2011, 332, 930–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Rouzic, A.; Boutin, T.S.; Capy, P. Long-term evolution of transposable elements. Proc. Natl. Acad. Sci. USA 2007, 104, 19375–19380. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, R.N.; Malik, H.S. Genetic conflicts: The usual suspects and beyond. J. Exp. Biol. 2017, 220, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Schaack, S.; Gilbert, C.; Feschotte, C. Promiscuous DNA: Horizontal transfer of transposable elements and why it matters for eukaryotic evolution. Trends Ecol. Evol. (Amst.) 2010, 25, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.E.; Craig, N.L. Tn7: Smarter than we thought. Nat. Rev. Mol. Cell Biol. 2001, 2, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Maldonado, H.; Gómez, M.J.; Moreno-Paz, M.; San Martín-Úriz, P.; Amils, R.; Parro, V.; López de Saro, F.J. Transposase interaction with the β sliding clamp: Effects on insertion sequence proliferation and transposition rate. Sci. Rep. 2015, 5, 13329. [Google Scholar] [CrossRef] [PubMed]
- Bastia, D.; Zaman, S. Mechanism and physiological significance of programmed replication termination. Semin. Cell Dev. Biol. 2014, 30, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Zaratiegui, M.; Castel, S.E.; Irvine, D.V.; Kloc, A.; Ren, J.; Li, F.; de Castro, E.; Marín, L.; Chang, A.-Y.; Goto, D.; et al. RNAi promotes heterochromatic silencing through replication-coupled release of RNA Pol II. Nature 2011, 479, 135–138. [Google Scholar] [CrossRef] [PubMed]
- McClintock, B. Induction of Instability at Selected Loci in Maize. Genetics 1953, 38, 579–599. [Google Scholar] [PubMed]
- McClintock, B. The origin and behavior of mutable loci in maize. Proc. Natl. Acad. Sci. USA 1950, 36, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Roeder, G.S.; Fink, G.R. DNA rearrangements associated with a transposable element in yeast. Cell 1980, 21, 239–249. [Google Scholar] [CrossRef]
- Chaleff, D.T.; Fink, G.R. Genetic events associated with an insertion mutation in yeast. Cell 1980, 21, 227–237. [Google Scholar] [CrossRef]
- Eichler, E.E.; Sankoff, D. Structural dynamics of eukaryotic chromosome evolution. Science 2003, 301, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Kazazian, H.H. Mobile elements: Drivers of genome evolution. Science 2004, 303, 1626–1632. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.; Pritham, E.J. DNA transposons and the evolution of eukaryotic genomes. Annu. Rev. Genet. 2007, 41, 331–368. [Google Scholar] [CrossRef] [PubMed]
- Garfinkel, D.J. Genome evolution mediated by Ty elements in Saccharomyces. Cytogenet. Genome Res. 2005, 110, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Batzer, M.A.; Deininger, P.L. Alu repeats and human genomic diversity. Nat. Rev. Genet. 2002, 3, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.E.; National Institute of Health Intramural Sequencing Center Comparative Sequencing Program; Cheng, Z.; Morrison, V.A.; Scherer, S.; Ventura, M.; Gibbs, R.A.; Green, E.D.; Eichler, E.E. Recurrent duplication-driven transposition of DNA during hominoid evolution. Proc. Natl. Acad. Sci. USA 2006, 103, 17626–17631. [Google Scholar] [PubMed]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Iskow, R.; Yang, L.; Gokcumen, O.; Haseley, P.; Luquette, L.J.; Lohr, J.G.; Harris, C.C.; Ding, L.; Wilson, R.K.; et al. Cancer Genome Atlas Research Network Landscape of somatic retrotransposition in human cancers. Science 2012, 337, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zuo, T.; Peterson, T. Generation of tandem direct duplications by reversed-ends transposition of maize ac elements. PLoS Genet. 2013, 9, e1003691. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, R. Deletions of a tyrosine tRNA gene in S. cerevisiae. Cell 1979, 17, 185–190. [Google Scholar] [CrossRef]
- Vitte, C.; Panaud, O. LTR retrotransposons and flowering plant genome size: Emergence of the increase/decrease model. Cytogenet. Genome Res. 2005, 110, 91–107. [Google Scholar] [CrossRef] [PubMed]
- Kupiec, M.; Petes, T.D. Allelic and ectopic recombination between Ty elements in yeast. Genetics 1988, 119, 549–559. [Google Scholar] [PubMed]
- Wallis, J.W.; Chrebet, G.; Brodsky, G.; Rolfe, M.; Rothstein, R. A hyper-recombination mutation in S. cerevisiae identifies a novel eukaryotic topoisomerase. Cell 1989, 58, 409–419. [Google Scholar] [CrossRef]
- Myung, K.; Datta, A.; Chen, C.; Kolodner, R.D. SGS1, the Saccharomyces cerevisiae homologue of BLM and WRN, suppresses genome instability and homeologous recombination. Nat. Genet. 2001, 27, 113–116. [Google Scholar] [PubMed]
- Rozenzhak, S.; Mejía-Ramírez, E.; Williams, J.S.; Schaffer, L.; Hammond, J.A.; Head, S.R.; Russell, P. Rad3 decorates critical chromosomal domains with gammaH2A to protect genome integrity during S-Phase in fission yeast. PLoS Genet. 2010, 6, e1001032. [Google Scholar] [CrossRef] [PubMed]
- Melamed, C.; Nevo, Y.; Kupiec, M. Involvement of cDNA in homologous recombination between Ty elements in Saccharomyces cerevisiae. Mol. Cell. Biol. 1992, 12, 1613–1620. [Google Scholar] [CrossRef] [PubMed]
- Nevo-Caspi, Y.; Kupiec, M. cDNA-mediated Ty recombination can take place in the absence of plus-strand cDNA synthesis, but not in the absence of the integrase protein. Curr. Genet. 1997, 32, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Hoff, E.F.; Levin, H.L.; Boeke, J.D. Schizosaccharomyces pombe retrotransposon Tf2 mobilizes primarily through homologous cDNA recombination. Mol. Cell. Biol. 1998, 18, 6839–6852. [Google Scholar] [CrossRef] [PubMed]
- Ben-Aroya, S.; Koren, A.; Liefshitz, B.; Steinlauf, R.; Kupiec, M. ELG1, a yeast gene required for genome stability, forms a complex related to replication factor C. Proc. Natl. Acad. Sci. USA 2003, 100, 9906–9911. [Google Scholar] [CrossRef] [PubMed]
- Scholes, D.T.; Banerjee, M.; Bowen, B.; Curcio, M.J. Multiple regulators of Ty1 transposition in Saccharomyces cerevisiae have conserved roles in genome maintenance. Genetics 2001, 159, 1449–1465. [Google Scholar] [PubMed]
- Bryk, M.; Banerjee, M.; Conte, D.; Curcio, M.J. The Sgs1 helicase of Saccharomyces cerevisiae inhibits retrotransposition of Ty1 multimeric arrays. Mol. Cell. Biol. 2001, 21, 5374–5388. [Google Scholar] [CrossRef] [PubMed]
- Stamenova, R.; Maxwell, P.H.; Kenny, A.E.; Curcio, M.J. Rrm3 protects the Saccharomyces cerevisiae genome from instability at nascent sites of retrotransposition. Genetics 2009, 182, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Burkett, T.J.; Garfinkel, D.J. Efficient homologous recombination of Ty1 element cDNA when integration is blocked. Mol. Cell. Biol. 1994, 14, 6540–6551. [Google Scholar] [CrossRef] [PubMed]
- Nevo-Caspi, Y.; Kupiec, M. Transcriptional induction of Ty recombination in yeast. Proc. Natl. Acad. Sci. USA 1994, 91, 12711–12715. [Google Scholar] [CrossRef] [PubMed]
- Cam, H.P.; Noma, K.-I.; Ebina, H.; Levin, H.L.; Grewal, S.I.S. Host genome surveillance for retrotransposons by transposon-derived proteins. Nature 2008, 451, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Nikolov, I.; Taddei, A. Linking replication stress with heterochromatin formation. Chromosoma 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.C.; Karpen, G.H. H3K9 methylation and RNA interference regulate nucleolar organization and repeated DNA stability. Nat. Cell. Biol. 2006, 9, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.C.; Karpen, G.H. Heterochromatic genome stability requires regulators of histone H3 K9 methylation. PLoS Genet. 2009, 5, e1000435. [Google Scholar] [CrossRef] [PubMed]
- Castel, S.E.; Ren, J.; Bhattacharjee, S.; Chang, A.-Y.; Sánchez, M.; Valbuena, A.; Antequera, F.; Martienssen, R.A. Dicer promotes transcription termination at sites of replication stress to maintain genome stability. Cell 2014, 159, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Zeller, P.; Padeken, J.; van Schendel, R.; Kalck, V.; Tijsterman, M.; Gasser, S.M. Histone H3K9 methylation is dispensable for Caenorhabditis elegans development but suppresses RNA:DNA hybrid-associated repeat instability. Nat. Genet. 2016, 48, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Roy-Engel, A.M.; Carroll, M.L.; El-Sawy, M.; Salem, A.-H.; Garber, R.K.; Nguyen, S.V.; Deininger, P.L.; Batzer, M.A. Non-traditional Alu evolution and primate genomic diversity. J. Mol. Biol. 2002, 316, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Ellison, C.E.; Bachtrog, D. Non-allelic gene conversion enables rapid evolutionary change at multiple regulatory sites encoded by transposable elements. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Moore, D.P.; Blomberg, M.A.; Braiterman, L.T.; Voytas, D.F.; Natsoulis, G.; Boeke, J.D. Hotspots for unselected Ty1 transposition events on yeast chromosome III are near tRNA genes and LTR sequences. Cell 1993, 73, 1007–1018. [Google Scholar] [CrossRef]
- Libuda, D.E.; Winston, F. Amplification of histone genes by circular chromosome formation in Saccharomyces cerevisiae. Nature 2006, 443, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Libuda, D.E.; Winston, F. Alterations in DNA replication and histone levels promote histone gene amplification in Saccharomyces cerevisiae. Genetics 2010, 184, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Dunham, M.J.; Badrane, H.; Ferea, T.; Adams, J.; Brown, P.O.; Rosenzweig, F.; Botstein, D. Characteristic genome rearrangements in experimental evolution of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2002, 99, 16144–16149. [Google Scholar] [CrossRef] [PubMed]
- Voineagu, I.; Narayanan, V.; Lobachev, K.S.; Mirkin, S.M. Replication stalling at unstable inverted repeats: Interplay between DNA hairpins and fork stabilizing proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 9936–9941. [Google Scholar] [CrossRef] [PubMed]
- Wallace, N.A.; Belancio, V.P.; Deininger, P.L. L1 mobile element expression causes multiple types of toxicity. Gene 2008, 419, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, N.; Lutz-Prigge, S.; Moran, J.V. Genomic deletions created upon LINE-1 retrotransposition. Cell 2002, 110, 315–325. [Google Scholar] [CrossRef]



© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaratiegui, M. Cross-Regulation between Transposable Elements and Host DNA Replication. Viruses 2017, 9, 57. https://doi.org/10.3390/v9030057
Zaratiegui M. Cross-Regulation between Transposable Elements and Host DNA Replication. Viruses. 2017; 9(3):57. https://doi.org/10.3390/v9030057
Chicago/Turabian StyleZaratiegui, Mikel. 2017. "Cross-Regulation between Transposable Elements and Host DNA Replication" Viruses 9, no. 3: 57. https://doi.org/10.3390/v9030057
APA StyleZaratiegui, M. (2017). Cross-Regulation between Transposable Elements and Host DNA Replication. Viruses, 9(3), 57. https://doi.org/10.3390/v9030057