
LTR-Retrotransposons from Bdelloid Rotifers Capture Additional ORFs Shared between Highly Diverse Retroelement Types




Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatics
2.2. Nucleic Acid Manipulations
3. Results
3.1. An Overview of LTR Retrotransposon Structure in Bdelloids
3.2. Types of Acquired Env-Like ORFs
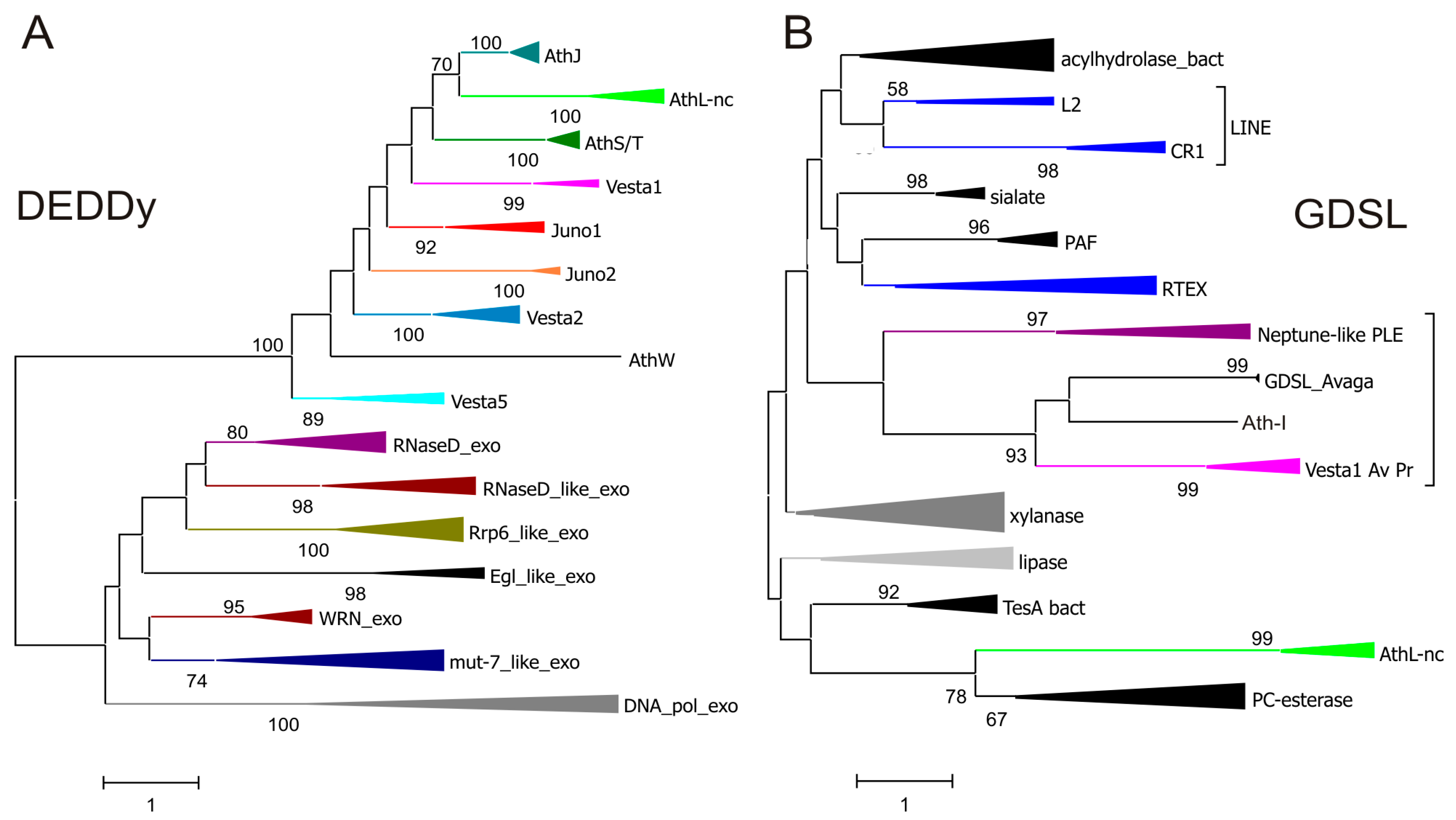
3.3. Unexpected Diversity of Non-Envelope ORF3 Functions
3.4. Different ORF3 Types Are Shared between Highly Diverse Retroelements
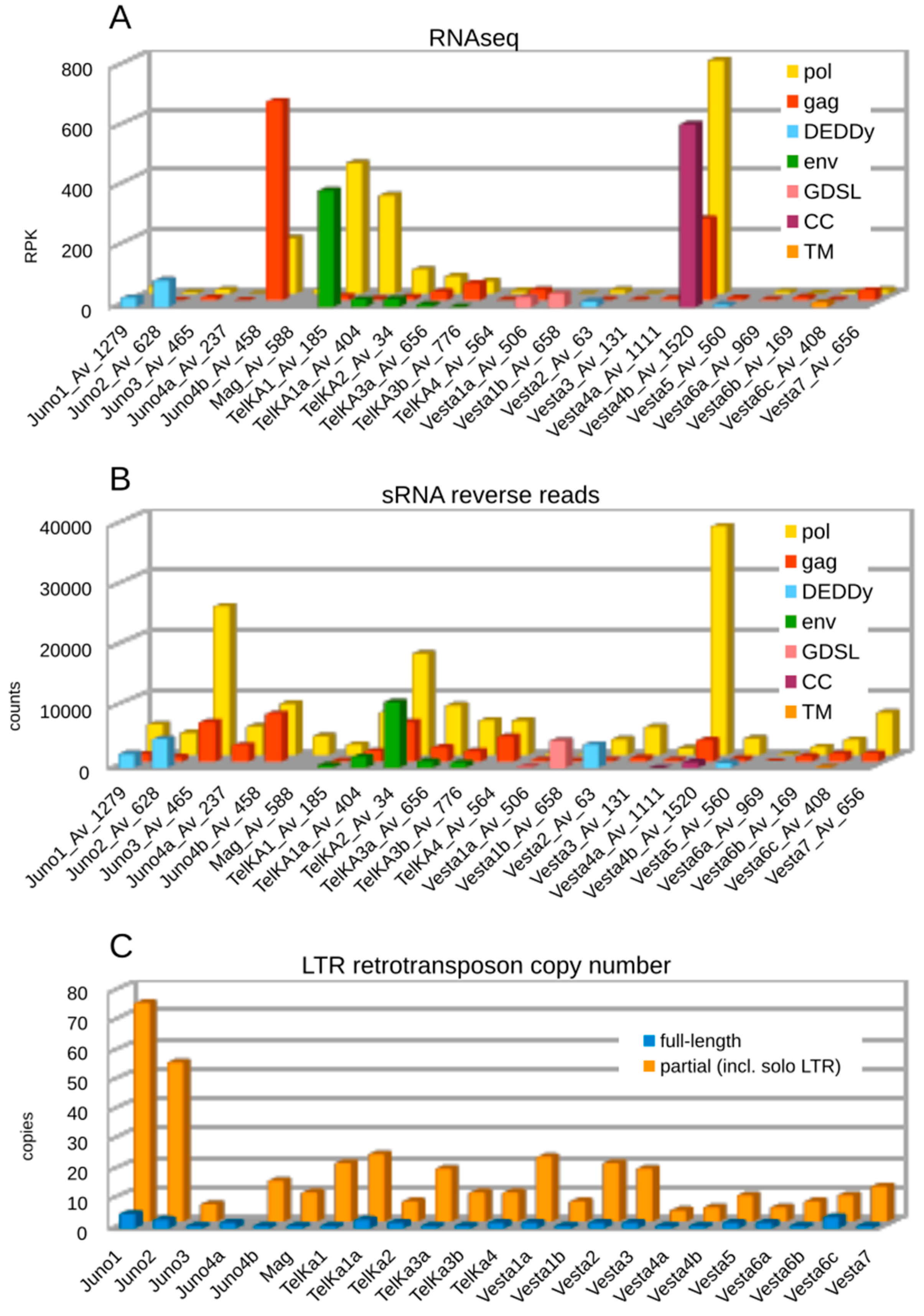
3.5. Transcription, Small RNA-Mediated Silencing, and Copy Numbers
3.6. Sequence Variation in Env-Like and GDSL-Like ORFs
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Craig, N.L.; Chandler, M.; Gellert, M.; lambowitz, A.M.; Rice, P.A.; Sandmeyer, S.B. Mobile DNA III; ASM Press: Washington, DC, USA, 2015. [Google Scholar]
- Arkhipova, I.R.; Mazo, A.M.; Cherkasova, V.A.; Gorelova, T.V.; Schuppe, N.G.; Ilyin, Y.V. The steps of reverse transcription of Drosophila mobile genetic elements and U3-R-U5 structure of their LTRs. Cell 1986, 44, 555–563. [Google Scholar] [CrossRef]
- Malik, H.S.; Henikoff, S.; Eickbush, T.H. Poised for contagion: Evolutionary origins of the infectious abilities of invertebrate retroviruses. Genome Res. 2000, 10, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Terzian, C.; Santamaria, P.; Pélisson, A.; Prud’homme, N.; Bucheton, A. Retroviruses in invertebrates: The gypsy retrotransposon is apparently an infectious retrovirus of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1994, 91, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Song, S.U.; Gerasimova, T.; Kurkulos, M.; Boeke, J.D.; Corces, V.G. An env-like protein encoded by a Drosophila retroelement: Evidence that gypsy is an infectious retrovirus. Genes Dev. 1994, 8, 2046–2057. [Google Scholar] [CrossRef] [PubMed]
- Misseri, Y.; Cerutti, M.; Devauchelle, G.; Bucheton, A.; Terzian, C. Analysis of the Drosophila gypsy endogenous retrovirus envelope glycoprotein. J. Gen. Virol. 2004, 85, 3325–3331. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Lee, X.; Li, X.-p.; Veldman, G.M.; Finnerty, H.; Racie, L.; LaVallie, E.; Tang, X.-Y.; Edouard, P.; Howes, S.; et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature 2000, 403, 785–789. [Google Scholar] [PubMed]
- Malik, H.S.; Henikoff, S. Positive selection of Iris, a retroviral envelope-derived host gene in Drosophila melanogaster. PLoS Genet. 2005, 1, e44. [Google Scholar] [CrossRef] [PubMed]
- Malfavon-Borja, R.; Feschotte, C. Fighting fire with fire: Endogenous retrovirus envelopes as restriction factors. J. Virol. 2015, 89, 4047–4050. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, E.A.; Meselson, M.; Arkhipova, I.R. Massive horizontal gene transfer in bdelloid rotifers. Science 2008, 320, 1210–1213. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, E.; Meselson, M. Extreme resistance of bdelloid rotifers to ionizing radiation. Proc. Natl. Acad. Sci. USA 2008, 105, 5139–5144. [Google Scholar] [CrossRef] [PubMed]
- Mark Welch, D.B.; Mark Welch, J.L.; Meselson, M. Evidence for degenerate tetraploidy in bdelloid rotifers. Proc. Natl. Acad. Sci. USA 2008, 105, 5145–5149. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, E.A.; Meselson, M.; Arkhipova, I.R. A deep-branching clade of retrovirus-like retrotransposons in bdelloid rotifers. Gene 2007, 390, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, E.; Arkhipova, I.R. Telomere-associated endonuclease-deficient Penelope-like retroelements in diverse eukaryotes. Proc. Natl. Acad. Sci. USA 2007, 104, 9352–9357. [Google Scholar] [CrossRef] [PubMed]
- Flot, J.F.; Hespeels, B.; Li, X.; Noel, B.; Arkhipova, I.; Danchin, E.G.; Hejnol, A.; Henrissat, B.; Koszul, R.; Aury, J.M.; et al. Genomic evidence for ameiotic evolution in the bdelloid rotifer Adineta vaga. Nature 2013, 500, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Alva, V.; Nam, S.-Z.; Söding, J.; Lupas, A.N. The MPI bioinformatics Toolkit as an integrative platform for advanced protein sequence and structure analysis. Nucleic Acids Res. 2016, 44, W410–W415. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, F.; Arkhipova, I.R. Multitasking of the piRNA silencing machinery: Targeting transposable elements and foreign genes in the bdelloid rotifer Adineta vaga. Genetics 2016, 203, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J. BLAT—The BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Green, P. Consed: A graphical editor for next-generation sequencing. Bioinformatics 2013, 29, 2936–2937. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a database of repetitive elements in eukaryotic genomes. Mob. DNA 2015, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.H.; Van Doninck, K.; Mandigo, M.L.; Meselson, M. Degenerate tetraploidy was established before bdelloid rotifer families diverged. Mol. Biol. Evol. 2009, 26, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Malik, H.S.; Eickbush, T.H. Modular evolution of the integrase domain in the Ty3/Gypsy class of LTR retrotransposons. J. Virol. 1999, 73, 5186–5190. [Google Scholar] [PubMed]
- Hizi, A.; Herzig, E. dUTPase: The frequently overlooked enzyme encoded by many retroviruses. Retrovirology 2015, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Chong, A.Y.; Kojima, K.K.; Jurka, J.; Ray, D.A.; Smit, A.F.A.; Isberg, S.R.; Gongora, J. Evolution and gene capture in ancient endogenous retroviruses-insights from the crocodilian genomes. Retrovirology 2014, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, C.; Rubini, A.; Belfiori, B.; Passeri, V.; Paolocci, F.; Arcioni, S. Tmt1: The first LTR-retrotransposon from a Tuber spp. Curr. Genet. 2008, 53, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.A.; Paterson, R.G.; Jardetzky, T.S. Paramyxovirus membrane fusion: Lessons from the F and HN atomic structures. Virology 2006, 344, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Laten, H.M.; Majumdar, A.; Gaucher, E.A. SIRE-1, a copia/Ty1-like retroelement from soybean, encodes a retroviral envelope-like protein. Proc. Natl. Acad. Sci. USA 1998, 95, 6897–6902. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Tian, Z.; Hans, C.S.; Laten, H.M.; Cannon, S.B.; Jackson, S.A.; Shoemaker, R.C.; Ma, J. Evolutionary conservation, diversity and specificity of LTR-retrotransposons in flowering plants: Insights from genome-wide analysis and multi-specific comparison. Plant J. 2010, 63, 584–598. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.A.; Voytas, D.F. Athila4 of Arabidopsis and Calypso of Soybean Define a Lineage of Endogenous Plant Retroviruses. Genome Res. 2002, 12, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Peterson-Burch, B.D.; Wright, D.A.; Laten, H.M.; Voytas, D.F. Retroviruses in plants? Trends Genet. 2000, 16, 151–152. [Google Scholar] [CrossRef]
- Vicient, C.M.; Kalendar, R.; Schulman, A.H. Envelope-class retrovirus-like elements are widespread, transcribed and spliced, and insertionally polymorphic in plants. Genome Res. 2001, 11, 2041–2049. [Google Scholar] [CrossRef] [PubMed]
- Steinbauerová, V.; Neumann, P.; Novák, P.; Macas, J. A widespread occurrence of extra open reading frames in plant Ty3/gypsy retrotransposons. Genetica 2011, 139, 1543–1555. [Google Scholar] [CrossRef] [PubMed]
- Aiewsakun, P.; Katzourakis, A. Marine origin of retroviruses in the early Palaeozoic Era. Nat. Commun. 2017, 8, 13954. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Deutcher, M.P. Exoribonuclease superfamilies: Structural analysis and phylogenetic distribution. Nucleic Acids Res. 2001, 29, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Akoh, C.C.; Lee, G.C.; Liaw, Y.C.; Huang, T.H.; Shaw, J.F. GDSL family of serine esterases/lipases. Prog. Lipid Res. 2004, 43, 534–552. [Google Scholar] [CrossRef] [PubMed]
- Evgen’ev, M.B.; Arkhipova, I.R. Penelope-like elements—A new class of retroelements: Distribution, function and possible evolutionary significance. Cytogenet. Genome Res. 2005, 110, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Anantharaman, V.; Aravind, L. Novel eukaryotic enzymes modifying cell-surface biopolymers. Biol. Direct 2010, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Kapitonov, V.V.; Jurka, J. The esterase and PHD domains in CR1-like non-LTR retrotransposons. Mol. Biol. Evol. 2003, 20, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.M.; Schmidt, S.; Jonas, S.; Vollmer, B.; Khazina, E.; Weichenrieder, O. Structure and properties of the esterase from non-LTR retrotransposons suggest a role for lipids in retrotransposition. Nucleic Acids Res. 2013, 41, 10563–10572. [Google Scholar] [CrossRef] [PubMed]
- Hestand, M.S.; Houdt, J.V.; Cristofoli, F.; Vermeesch, J.R. Polymerase specific error rates and profiles identified by single molecule sequencing. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2016, 784–785, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Ketting, R.F.; Haverkamp, T.H.; van Luenen, H.G.; Plasterk, R.H. Mut-7 of C. elegans, required for transposon silencing and RNA interference, is a homolog of Werner syndrome helicase and RNaseD. Cell 1999, 99, 133–141. [Google Scholar] [CrossRef]
- Arkhipova, I.; Meselson, M. Deleterious transposable elements and the extinction of asexuals. Bioessays 2005, 27, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-M.; Campbell, E.; Pandey, R.R.; Yang, Z.; McCarthy, A.A.; Pillai, R.S. Metazoan Maelstrom is an RNA-binding protein that has evolved from an ancient nuclease active in protists. RNA 2015, 21, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chen, K.-M.; Pandey, R.R.; Homolka, D.; Reuter, M.; Janeiro, B.K.R.; Sachidanandam, R.; Fauvarque, M.-O.; McCarthy, A.A.; Pillai, R.S. PIWI Slicing and EXD1 Drive Biogenesis of Nuclear piRNAs from Cytosolic Targets of the Mouse piRNA Pathway. Mol. Cell 2016, 61, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Arkhipova, I. Distribution and phylogeny of Penelope-like elements in eukaryotes. Syst. Biol. 2006, 55, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Magiorkinis, G.; Gifford, R.J.; Katzourakis, A.; De Ranter, J.; Belshaw, R. Env-less endogenous retroviruses are genomic superspreaders. Proc. Natl. Acad. Sci. USA 2012, 109, 7385–7390. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Clone | Reference Scaffold/Contig 1 | Substitutions, bp | Substitutions, aa | Natural aa Differences | Unique aa Differences |
|---|---|---|---|---|---|
| env1 | 1591/5150 | 4 | 4 | R-Q, E-Q, T-I | I-V |
| env1a.1 | 3 | 2 | I-T, V-A | ||
| env1a.4 | 1200/4393 | 0 | 0 | ||
| env1a.8 | 3 | 3 | V-A, T-I | S-F | |
| env2.1 | 34/303 | 0 | 0 | ||
| env2.2 | 2 | 1 | I-T | ||
| env2.3 | 4 | 3 | V-I, I-T | M-I | |
| env2n.1 | 680/3155 | 1 | 1 | A-S | |
| env2n.2 | 2 | 1 | D-G | ||
| env3b.1 | 776/3459 | 0 | 0 | ||
| env3b.2 | 1 | 1 | I-T | ||
| ves1 | 494/2540 | 8 | 3 | T-S, R-S, H-Q | |
| ves1a | 506/2575 | 1 | 0 | silent | |
| ves1b.1 | 658/3084 | 1 | 0 | silent | |
| ves1b.3 | 1 | 0 | silent |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez, F.; Kenefick, A.W.; Arkhipova, I.R. LTR-Retrotransposons from Bdelloid Rotifers Capture Additional ORFs Shared between Highly Diverse Retroelement Types. Viruses 2017, 9, 78. https://doi.org/10.3390/v9040078
Rodriguez F, Kenefick AW, Arkhipova IR. LTR-Retrotransposons from Bdelloid Rotifers Capture Additional ORFs Shared between Highly Diverse Retroelement Types. Viruses. 2017; 9(4):78. https://doi.org/10.3390/v9040078
Chicago/Turabian StyleRodriguez, Fernando, Aubrey W. Kenefick, and Irina R. Arkhipova. 2017. "LTR-Retrotransposons from Bdelloid Rotifers Capture Additional ORFs Shared between Highly Diverse Retroelement Types" Viruses 9, no. 4: 78. https://doi.org/10.3390/v9040078
APA StyleRodriguez, F., Kenefick, A. W., & Arkhipova, I. R. (2017). LTR-Retrotransposons from Bdelloid Rotifers Capture Additional ORFs Shared between Highly Diverse Retroelement Types. Viruses, 9(4), 78. https://doi.org/10.3390/v9040078