
Porcine Circoviruses and Xenotransplantation


Abstract
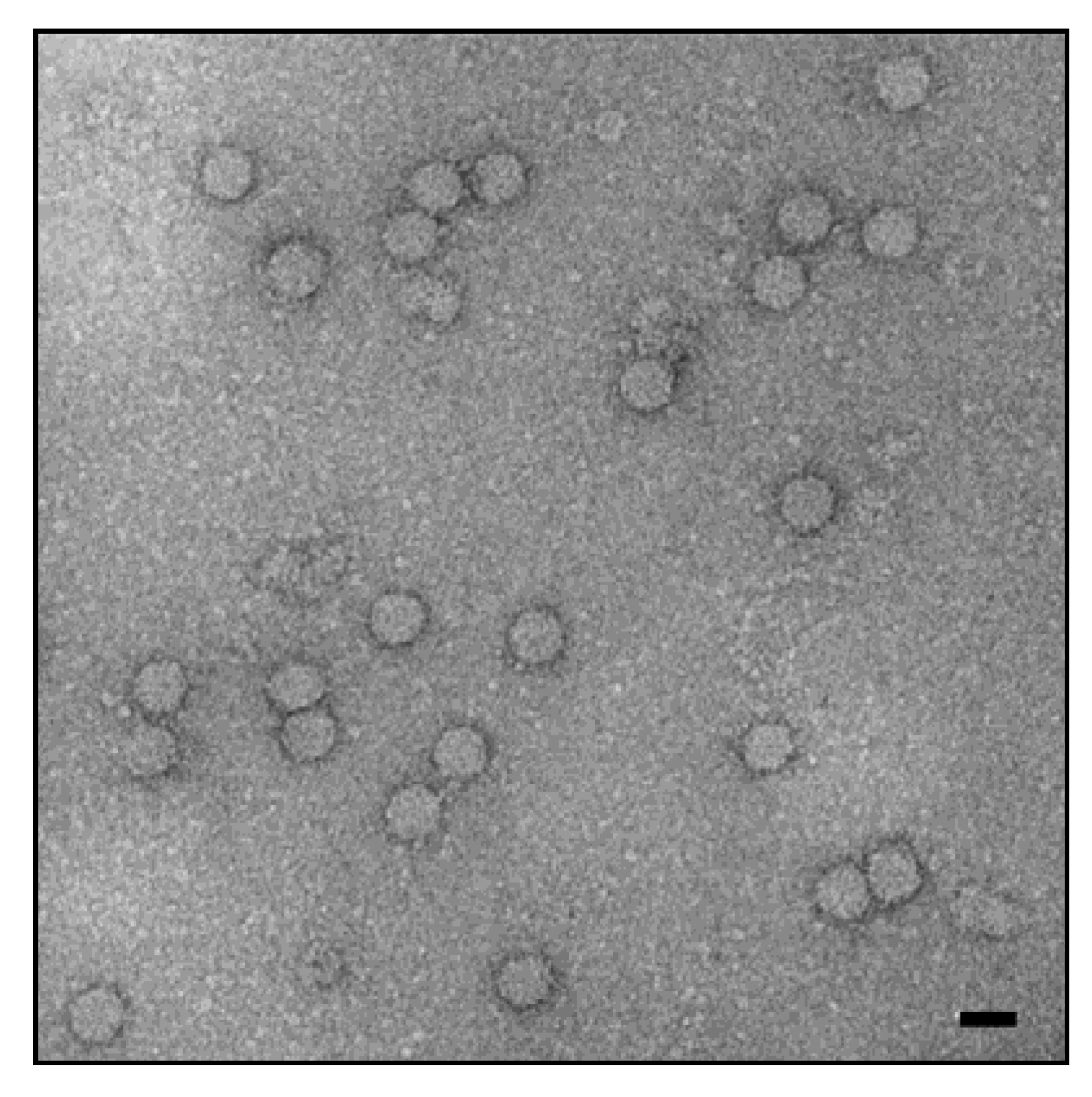
:1. Introduction
2. Diagnosis and Transmission
3. PCV2-Related Diseases in Pigs
4. PCV Does Not Infect Immunocompetent Humans
5. PCV2 and First Preclinical and Clinical Xenotransplantations
6. Treatment and Vaccination
7. How to Eliminate PCV
8. Summary
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fishman, J.A. Infection in solid-organ transplant recipients. N. Engl. J. Med. 2007, 357, 2601–2614. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.A.; Razonable, R.R. Infections in liver transplant recipients. World J. Hepatol. 2011, 3, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Kim, S. Bacterial infection after liver transplantation. World J. Gastroenterol. 2014, 20, 6211–6220. [Google Scholar] [CrossRef] [PubMed]
- Petrosillo, N.; Puro, V.; D’Anna, C.; Marquez, J.R.; Ippolito, G. HIV infection after kidney transplantation. Nephron 1996, 72, 124. [Google Scholar] [CrossRef] [PubMed]
- Vora, N.M.; Basavaraju, S.V.; Feldman, K.A.; Paddock, C.D.; Orciari, L.; Gitterman, S.; Griese, S.; Wallace, R.M.; Said, M.; Blau, D.M.; et al. Raccoon rabies virus variant transmission through solid organ transplantation. JAMA 2013, 310, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Schwarting, A.; Mauer, D.; Ross, R.S.; Martens, A.; Kliem, V.; Wahl, J.; Panning, M.; Baumgarte, S.; Müller, T.; et al. Management and outcomes after multiple corneal and solid organ transplantations from a donor infected with rabies virus. Clin. Infect. Dis. 2010, 50, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Burton, E.C.; Kuehnert, M.J.; Rupprecht, C.; Sutker, W.L.; Ksiazek, T.G.; Paddock, C.D.; Guarner, J.; Shieh, W.J.; Goldsmith, C.; et al. Transmission of rabies virus from an organ donor to four transplant recipients. N. Engl. J. Med. 2005, 352, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, P.; Razonable, R.R. Cytomegalovirus infections in solid organ transplantation: A review. Infect. Chemother. 2013, 45, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Razonable, R.R. Cytomegalovirus infection after liver transplantation: Current concepts and challenges. World J. Gastroenterol. 2008, 14, 4849–4860. [Google Scholar] [CrossRef] [PubMed]
- Lischka, P.; Zimmermann, H. Antiviral strategies to combat cytomegalovirus infections in transplant recipients. Curr. Opin. Pharmacol. 2008, 8, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Fishman, J.A. Assessment of infectious risk in clinical xenotransplantation: The lessons for clinical allotransplantation. Xenotransplantation 2014, 21, 307–308. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Tasaki, M.; Sekijima, M.; Wilkinson, R.A.; Villani, V.; Morgan, S.G.; Cormack, T.A.; Hanekamp, I.M.; Hawley, R.J.; Arn, J.S.; et al. Porcine cytomegalovirus infection is associated with early rejection of kidney grafts in a pig to baboon xenotransplantation model. Transplantation 2014, 98, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Sekijima, M.; Waki, S.; Sahara, H.; Tasaki, M.; Wilkinson, R.A.; Villani, V.; Shimatsu, Y.; Nakano, K.; Matsunari, H.; Nagashima, H.; et al. Result of life-supporting galactosyltransferase knockout swine. Transplantation 2014, 98, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Abicht, J.M.; Reichart, B.; Mayr, T.; Guethoff, S.; Denner, J. Active replication of porcine cytomegalovirus (PSMV) following transplantation of a pig heart into a baboon despite undetected virus in the donor pig. Ann. Virol. Res. 2016, 2, 1018. [Google Scholar]
- Morozov, V.A.; Plotzki, E.; Rotem, A.; Barkai, U.; Denner, J. Extended microbiologicial characterization of Göttingen minipigs: Porcine cytomegalovirus and other viruses. Xenotransplantation 2016, 24, 34–40. [Google Scholar]
- Harding, J.C.; Baker, C.; Rhodes, C.; McIntosh, K.A.; Bonneau, M. Ring tests to evaluate the performance of porcine circovirus-2 (PCV-2) polymerase chain reaction (PCR) assays used in North American diagnostic laboratories. Can. J. Vet. Res. 2009, 73, 7–14. [Google Scholar] [PubMed]
- Denner, J. Xenotransplantation and hepatitis E virus. Xenotransplantation 2015, 22, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Finsterbusch, T.; Mankertz, A. Porcine circoviruses—Small but powerful. Virus Res. 2009, 143, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Tischer, I.; Rasch, R.; Tochtermann, G. Characterization of papovavirus-and picornavirus-like particles in permanent pig kidney cell lines. Zentralbl. Bakteriol. Orig. A. 1974, 226, 153–167. [Google Scholar] [PubMed]
- Lima, F.E.; Cibulski, S.P.; Dall Bello, A.G.; Mayer, F.Q.; Witt, A.A.; Roehe, P.M.; d’Azevedo, P.A. A novel chiropteran circovirus genome recovered from a Brazilian insectivorous bat species. Genome Announc. 2015, 3, e01393-15. [Google Scholar] [CrossRef] [PubMed]
- Han, H.J.; Wen, H.L.; Zhao, L..; Liu, J.W.; Luo, L.M.; Zhou, C.M.; Qin, X.R.; Zhu, Y.L.; Liu, M.M.; Qi, R.; et al. Novel coronaviruses; astroviruses; adenoviruses and circoviruses in insectivorous bats from northern China. Zoonoses Public Health 2017. [Google Scholar] [CrossRef]
- Kapoor, A.; Dubovi, E.J.; Henriquez-Rivera, J.A.; Lipkin, W.I. Complete genome sequence of the first canine circovirus. J. Virol. 2012, 86, 7018. [Google Scholar] [CrossRef] [PubMed]
- Zaccaria, G.; Malatesta, D.; Scipioni, G.; Di Felice, E.; Campolo, M.; Casaccia, C.; Savini, G.; Di Sabatino, D.; Lorusso, A. Circovirus in domestic and wild carnivores: An important opportunistic agent? Virology 2016, 490, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Thaiwong, T.; Wise, A.G.; Maes, R.K.; Mullaney, T.; Kiupel, M. Canine circovirus 1 (CaCV-1) and canine parvovirus 2 (CPV-2): Recurrent dual Infections in a Papillon breeding colony. Vet. Pathol. 2016, 53, 1204–1209. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.; Hartmann, K.; Leutenegger, C.M.; Proksch, A.L.; Mueller, R.S.; Unterer, S. Role of canine circovirus in dogs with acute haemorrhagic diarrhoea. Vet. Rec. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.; Liu, Y.; Li, N.; Wang, Y.; Zhang, S.; Hu, R. Novel circovirus from mink; China. Emerg. Infect. Dis. 2014, 20, 1548–1550. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.S.; Sun, N.; Tian, F.L.; Wen, Y.J.; Xu, C.; Li, J.; Chen, Q.; Wang, J.B. Genetic analysis of porcine circovirus type 2 from dead minks. J. Gen. Virol. 2016, 97, 2316–2322. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.R.; Opriessnig, T. Epidemiology and horizontal transmission of porcine circovirus type 2 (PCV2). Anim. Health Res. Rev. 2010, 11, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Finsterbusch, T.; Steinfeldt, T.; Doberstein, K.; Rödner, C.; Mankertz, A. Interaction of the replication proteins and the capsid protein of porcine circovirus type 1 and 2 with host proteins. Virology 2009, 386, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Guo, K.; Wang, T.; Zhang, C.; Zhang, Y. Porcine circovirus type 2 ORF4 protein binds heavy chain ferritin. J. Biosci. 2015, 40, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Guo, K.; Xu, H.; Wang, T.; Zhang, Y. Identification of putative ORF5 protein of porcine circovirus type 2 and functional analysis of GFP-fused ORF5 protein. PLoS ONE 2015, 10, e0127859. [Google Scholar]
- Allan, G.M.; McNeilly, F.; Foster, J.C.; Adair, B.M. Infection of leucocyte cell cultures derived from different species with pig circovirus. Vet. Microbiol. 1994, 41, 267–279. [Google Scholar] [CrossRef]
- Sanchez, R.E., Jr.; Meerts, P.; Nauwynck, H.J.; Pensaert, M.B. Change of porcine circovirus 2 target cells in pigs during development from fetal to early postnatal life. Vet. Microbiol. 2003, 95, 15–25. [Google Scholar] [CrossRef]
- Meerts, P.; Misinzo, G.; McNeilly, F.; Nauwynck, H.J. Replication kinetics of different porcine circovirus 2 strains in PK-15 cells, fetal cardiomyocytes and macrophages. Arch. Virol. 2005, 150, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Darwich, L.; Segales, J.; Mateu, E. Pathogenesis of postweaning multisystemic wasting syndrome caused by porcine circovirus 2: An immune riddle. Arch. Virol. 2004, 149, 857–874. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.L.; Sattentau, Q.J. Attachment factors. Adv. Exp. Med. Biol. 2013, 790, 1–23. [Google Scholar] [PubMed]
- Misinzo, G.; Delputte, P.L.; Meerts, P.; Lefebvre, D.J.; Nauwynck, H.J. Porcine circovirus 2 uses heparan sulfate and chondroitin sulfate B glycosaminoglycans as receptors for its attachment to host cells. J. Virol. 2006, 80, 3487–3494. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.-J. Porcine circovirus type 2 (PCV2): Pathogenesis and interaction with the immune system. Annu. Rev. Anim. Biosci. 2013, 1, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J.; Mateu, E. Immunosuppression as a feature of postweaning multisystemic wasting syndrome. Vet. J. 2006, 171, 396–397. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.; Opriessnig, T.; Meng, X.J.; Pelzer, K.; Buechner-Maxwell, V. Porcine circovirus type 2 and porcine circovirus-associated disease. J. Vet. Intern. Med. 2009, 23, 1151–1163. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J.; Allan, G.M.; Domingo, M. Porcine circovirus diseases. Anim. Health Res. Rev. 2005, 6, 119–142. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Langohr, I. Current state of knowledge on porcine circovirus type 2-associated lesions. Vet. Pathol. 2013, 50, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Meng, X.J.; Halbur, P.G. Porcine circovirus type 2 associated disease: Update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J. Vet. Diagn. Investig. 2007, 19, 591–615. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Sensitive methods and improved screening strategies are needed for the detection of pig viruses. Xenotransplantation. in press.
- Quintana, J.; Segalés, J.; Calsamiglia, M.; Domingo, M. Detection of porcine circovirus type 1 in commercial pig vaccines using polymerase chain reaction. Vet. J. 2006, 171, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J.; Calsamiglia, M.; Olvera, A.; Sibila, M.; Badiella, L.; Domingo, M. Quantification of porcine circovirus type 2 (PCV2) DNA in serum and tonsillar, nasal, tracheo-bronchial, urinary and faecal swabs of pigs with and without postweaning multisystemic wasting syndrome (PMWS). Vet. Microbiol. 2005, 111, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Mankertz, A.; Domingo, M.; Folch, J.M.; LeCann, P.; Jestin, A.; Segalés, J.; Chmielewicz, B.; Plana-Durán, J.; Soike, D. Characterisation of PCV-2 isolates from Spain, Germany and France. Virus Res. 2000, 66, 65–77. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.; Liu, L.; Li, R.; Yuan, W. Rapid detection of Porcine circovirus 2 by recombinase polymerase amplification. J. Vet. Diagn. Investig. 2016, 28, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Lin, H.; Chen, S.; Yang, M.; Yan, Q.; Wen, C.; Hao, Z.; Yan, Y.; Sun, Y.; Hu, J.; et al. Sensitive detection of Porcine circovirus-2 by droplet digital polymerase chain reaction. J. Vet. Diagn. Investig. 2015, 27, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Pang, V.F.; Lee, F.; Liao, P.C.; Huang, Y.L.; Lin, Y.L.; Lai, S.S.; Jeng, C.R. Development and evaluation of a loop-mediated isothermal amplification method for rapid detection and differentiation of two genotypes of porcine circovirus type 2. J. Microbiol. Immunol. Infect. 2014, 47, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Pérez, L.J.; Perera, C.L.; Frías, M.T.; Núñez, J.I.; Ganges, L.; de Arce, H.D. A multiple SYBR Green I-based real-time PCR system for the simultaneous detection of porcine circovirus type 2; porcine parvovirus; pseudorabies virus and Torque teno sus virus 1 and 2 in pigs. J. Virol. Methods. 2012, 179, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Blomström, A.L.; Belák, S.; Fossum, C.; Fuxler, L.; Wallgren, P.; Berg, M. Studies of porcine circovirus type 2; porcine boca-like virus and torque teno virus indicate the presence of multiple viral infections in postweaning multisystemic wasting syndrome pigs. Virus Res. 2010, 152, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, P.; Mahé, D.; Cariolet, R.; Truong, C.; Le Dimna, M.; Arnauld, C.; Rose, N.; Eveno, E.; Albina, E.; Madec, F.; et al. An ORF2 protein-based ELISA for porcine circovirus type 2 antibodies in post-weaning multisystemic wasting syndrome. Vet. Microbiol. 2003, 94, 183–194. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, L.; Willson, P.; O’Connor, B.; Keenliside, J.; Chirino-Trejo, M.; Babiuk, L. Seroprevalence of porcine circovirus type 2 in swine populations in Canada and Costa Rica. Can. J. Vet. Res. 2002, 66, 225–231. [Google Scholar] [PubMed]
- Patterson, A.R.; Johnson, J.; Ramamoorthy, S.; Ramamoorthy, S.; Meng, X.J.; Halbur, P.G.; Opriessnig, T. Comparison of three enzyme-linked immunosorbent assays to detect porcine circovirus-2 (PCV-2)-specific antibodies after vaccination or inoculation of pigs with distinct PCV-1 or PCV-2 isolates. J. Vet. Diagn. Investig. 2008, 20, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Mahé, D.; Blanchard, P.; Truong, C.; Arnauld, C.; Le Cann, P.; Cariolet, R.; Madec, F.; Albina, E.; Jestin, A. Differential recognition of ORF2 protein from type 1 and type 2 porcine circoviruses and identification of immunorelevant epitopes. J. Gen. Virol. 2000, 81, 1815–1824. [Google Scholar] [CrossRef] [PubMed]
- Nawagitgul, P.; Harms, P.A.; Morozov, I.; Thacker, B.J.; Sorden, S.D.; Lekcharoensuk, C.; Paul, P.S. Modified indirect porcine circovirus (PCV) type 2-based and recombinant capsid protein (ORF2)-based enzyme-linked immunosorbent assays for detection of antibodies to PCV. Clin. Diagn. Lab. Immunol. 2002, 9, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.R.; Johnson, J.K.; Ramamoorthy, S.; Patterson, A.R.; Johnson, J.K.; Ramamoorthy, S.; Hesse, R.A.; Murtaugh, M.P.; Puvanendiran, S.; Meng, X.J. Interlaboratory comparison of porcine circovirus-2 indirect immunofluorescent antibody test and enzyme-linked immunosorbent assay results on experimentally infected pigs. J. Vet. Diagn. Investig. 2011, 23, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, A.; Wang, C.; Prickett, J.R.; Pogranichniy, R.; Yoon, K.J.; Main, R.; Kurtz, A. Efficient surveillance of pig populations using oral fluids. Prev. Vet. Med. 2012, 104, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Allan, G.M.; Ellis, J.A. Porcine circoviruses: A review. J. Vet. Diagn. Investig. 2000, 12, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.R.; Madson, D.M.; Halbur, P.G.; Opriessnig, T. Shedding and infection dynamics of porcine circovirus type 2 (PCV2) after natural exposure. Vet. Microbiol. 2011, 149, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.R.; Ramamoorthy, S.; Madson, D.M.; Meng, X.J.; Halbur, P.G.; Opriessnig, T. Shedding and infection dynamics of porcine circovirus type 2 (PCV2) after experimental infection. Vet. Microbiol. 2011, 149, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Lang, Ch.; Griessler, A.; Pirker, E.; Söllner, H.; Segalés, J.; Kekarainen, T.; Ritzmann, M. Detection of porcine circovirus type 2 and Torque-teno-sus-virus 1 and 2 in semen samples of boars from an Austrian artificial insemination centre. Tierarztl. Prax. Ausg. G. Grosstiere Nutztiere 2011, 39, 201–204. [Google Scholar] [PubMed]
- Grasland, B.; Blanchard, P.; Kéranflech, A.; Bigault, L.; Oger, A.; Rose, N.; Madec, F.; Jestin, A.; Cariolet, R. Evaluation of the transmission of porcine circovirus type 2 (PCV-2) genogroups a and b with semen from infected specific-pathogen-free boars. Vet. Microbiol. 2013, 162, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.L.; Chen, S.N.; Zhang, J.W.; Wei, Z.Z.; Long, J.X.; Yuan, S.S.; Wei, W.K.; Chen, Q.L.; Wu, H.X.; Wu, D.C. Dissection of the possible routes on porcine circoviruses infecting human. J. Anim. Veterin. Adv. 2012, 11, 1281–1286. [Google Scholar]
- Larochelle, R.; Magar, S.; D’Allaire, S. Genetic characterization and phylogenetic analysis of porcine circovirus type 2 (PCV2) strains from cases presenting various clinical conditions. Virus Res. 2002, 90, 101–112. [Google Scholar] [CrossRef]
- Firth, C.; Charleston, M.A.; Duffy, S.; Shapiro, B.; Holmes, E.C. Insights into the evolutionary history of an emerging livestock pathogen: Porcine circovirus 2. J. Virol. 2009, 83, 12813–12821. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J.; Olvera, A.; Grau-Roma, L.; Charreyre, C.; Nauwynck, H.; Larsen, L.; Dupont, K.; McCullough, K.; Ellis, J.; Krakowka, S.; et al. PCV-2 genotype definition and nomenclature. Vet. Rec. 2008, 162, 867–868. [Google Scholar] [CrossRef] [PubMed]
- Rose, N.; Opriessnig, T.; Grasland, B.; Jestin, A. Epidemiology and transmission of porcine circovirus type 2 (PCV2). Virus Res. 2012, 164, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. J. Gen. Virol. 2015, 96, 1830–1841. [Google Scholar] [CrossRef] [PubMed]
- Ssemadaali, M.A.; Ilha, M.; Ramamoorthy, S. Genetic diversity of porcine circovirus type 2 and implications for detection and control. Res. Vet. Sci. 2015, 103, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhai, S.L.; Zeng, S.Y.; Sun, B.B.; Deng, S.F.; Chen, H.L.; Zheng, Y.; Wang, H.X.; Li, X.P.; Liu, J.K.; et al. Identification of natural recombinants derived from PCV2a and PCV2b. Genet. Mol. Res. 2015, 14, 11780–11790. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, C.A.; Music, N.; Fontaine, G.; Tremblay, D.; Harel, J. Emergence of a new type of porcine circovirus in swine (PCV): A type 1 and type 2 PCV recombinant. Vet. Microbiol. 2010, 144, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Phan, G.T.; Giannitti, F.; Rossow, S.; Marthaler, D.; Knutson, T.; Linlin, L.; Deng, X.; Resende, T.; Vannucci, F.; Delwart, E. Detection of a novel circovirus PCV3 in pigs with cardiac and multi-systemic inflammation. Virol. J. 2016, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Liu, X.; Zhang, P.; Wang, L.; Liu, Y.; Zhang, L.; Liang, P.; Song, C. Genome characterization of a porcine circovirus type 3 in South China. Transbound. Emerg. Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ku, X.; Chen, F.; Li, P.; Wang, Y.; Yu, X.; Fan, S.; Qian, P.; Wu, M.; He, Q. Identification and genetic characterization of porcine circovirus type 3 in China. Transbound. Emerg. Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel porcine circovirus distantly related to known circoviruses is associated with porcine dermatitis and nephropathy syndrome and reproductive failure. J. Virol. 2016, 91, e01879-16. [Google Scholar] [CrossRef] [PubMed]
- Meehan, B.M.; McNeilly, F.; McNair, I.; Walker, I.; Ellis, J.A.; Krakowka, S.; Allan, G.M. Isolation and characterization of porcine circovirus 2 from cases of sow abortion and porcine dermatitis and nephropathy syndrome. Arch. Virol. 2001, 146, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Rosell, C.; Segalés, J.; Ramos-Vara, J.A.; Folch, J.M.; Rodríguez-Arrioja, G.M.; Duran, C.O.; Balasch, M.; Plana-Durán, J.; Domingo, M. Identification of porcine circovirus in tissues of pigs with porcine dermatitis and nephropathy syndrome. Vet. Rec. 2000, 146, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Ladekjaer-Mikkelsen, A.S.; Nielsen, J.; Storgaard, T.; Bøtner, A.; Allan, G.; McNeilly, F. Transplacental infection with PCV-2 associated with reproductive failure in a gilt. Vet. Rec. 2001, 148, 759–760. [Google Scholar] [PubMed]
- West, K.H.; Bystrom, J.M.; Wojnarowicz, C.; Bøtner, A.; Allan, G.; McNeilly, F. Myocarditis and abortion associated with intrauterine infection of sows with porcine circovirus 2. J. Vet. Diagn. Investig. 1999, 11, 530–532. [Google Scholar] [CrossRef] [PubMed]
- Hattermann, K.; Roedner, C.; Schmitt, C.; Finsterbusch, T.; Steinfeldt, T.; Mankertz, A. Infection studies on human cell lines with porcine circovirus type 1 and porcine circovirus type 2. Xenotransplantation 2004, 11, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Dubin, G.; Toussaint, J.F.; Cassart, J.P.; Howe, B.; Boyce, D.; Friedland, L.; Abu-Elyazeed, R.; Poncelet, S.; Han, H.H.; Debrus, S. Investigation of a regulatory agency enquiry into potential porcine circovirus type 1 contamination of the human rotavirus vaccine, Rotarix: Approach and outcome. Hum. Vaccin Immunother. 2013, 9, 2398–2408. [Google Scholar] [CrossRef] [PubMed]
- Arteaga-Troncoso, G.; Guerra-Infante, F.; Rosales-Montano, L.M.; Diaz-Garcia, F.J.; Flores-Medina, S. Ultrastructural alterations in human blood leukocytes induced by porcine circovirus type 1 infection. Xenotransplantation 2005, 12, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Tischer, I.; Bode, L.; Apodaca, J.; Timm, H.; Peters, D.; Rasch, R.; Pociuli, S.; Gerike, E. Presence of antibodies reacting with porcine circovirus in sera of humans, mice, and cattle. Arch. Virol. 1995, 140, 1427–1439. [Google Scholar] [CrossRef] [PubMed]
- Victoria, J.G.; Wang, C.; Jones, M.S.; Victoria, J.G.; Wang, C.; Jones, M.S.; Jaing, C.; McLoughlin, K.; Gardner, S.; Delwart, E.L. Viral nucleic acids in live-attenuated vaccines: Detection of minority variants and an adventitious virus. J. Virol. 2010, 84, 6033–6040. [Google Scholar] [CrossRef] [PubMed]
- Gilliland, S.M.; Forrest, L.; Carre, H.; Jenkins, A.; Berry, N.; Martin, J.; Minor, P.; Schepelmann, S. Investigation of porcine circovirus contamination in human vaccines. Biologicals 2012, 40, 270–277. [Google Scholar] [CrossRef] [PubMed]
- McClenahan, S.D.; Krause, P.R.; Uhlenhaut, C. Molecular and infectivity studies of porcine circovirus in vaccines. Vaccine 2011, 29, 4745–4753. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Palacios, G.M.; Pérez-Schael, I.; Velázquez, F.R.; Abate, H.; Breuer, T.; Clemens, S.C.; Cheuvart, B.; Espinoza, F.; Gillard, P.; Innis, B.L.; et al. Human Rotavirus Vaccine Study Group. Safety and efficacy of an attenuated vaccine against severe rotavirus gastroenteritis. N. Engl. J. Med. 2006, 354, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Vesikari, T.; Clark, H.F.; Offit, P.A.; Dallas, M.J.; DiStefano, D.J.; Goveia, M.G.; Ward, R.L.; Schödel, F.; Karvonen, A.; Drummond, J.E.; et al. Effects of the potency and composition of the multivalent human-bovine (WC3) reassortant rotavirus vaccine on efficacy, safety and immunogenicity in healthy infants. Vaccine 2006, 24, 4821–4829. [Google Scholar] [CrossRef] [PubMed]
- Baylis, S.A.; Finsterbusch, T.; Bannert, N.; Blümel, J.; Mankertz, A. Analysis of porcine circovirus type 1 detected in Rotarix vaccine. Vaccine 2011, 29, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Paradis, K.; Langford, G.; Long, Z.; Heneine, W.; Sandstrom, P.; Switzer, W.M.; Chapman, L.E.; Lockey, C.; Onions, D. Otto ESearch for cross-species transmission of porcine endogenous retrovirus in patients treated with living pig tissue. The XEN 111 Study Group. Science 1999, 285, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Abalovich, A.; Wechsler, C.J.; Wynyard, S.; Elliott, R.B. Clinical Benefit of Islet Xenotransplantation for the Treatment of Type 1. EBio Med. 2016, 12, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Wynyard, S.; Nathu, D.; Garkavenko, O.; Denner, J.; Elliott, R.B. Microbiological safety of the first clinical pig islet xenotransplantation trial in New Zealand. Xenotransplantation 2014, 21, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.K.; Matsumoto, S.; Abalovich, A.; Itoh, T.; Mourad, N.I.; Gianello, P.R.; Wolf, E.; Cozzi, E. Progress in clinical encapsulated islet xenotransplantation. Transplantation 2016, 100, 2301–2308. [Google Scholar] [CrossRef] [PubMed]
- Morozov, A.V.; Wynyard, S.; Matsumoto, S.; Abalovich, A.; Denner, J.; Elliott, R.B. No PERV transmission during a clinical trial of pig islet cell transplantation. Virus Res. 2016, 227, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Muzina, M.; Muzina, Z.; Powels, K.; Elliot, R.B.; Croxson, M.C. Monitoring for potentially xenozoonotic virus in New Zealand pigs. J. Med. Virol. 2004, 72, 338–344. [Google Scholar]
- Garkavenko, O.; Dieckhoff, B.; Wynyard, S.; Denner, J.; Elliot, R.B.; Tan, P.L.; Croxson, M.C. Absence of transmission of potentially xenotic viruses in a prospective pig to primate islet xenotransplantation study. J. Med. Virol. 2008, 80, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.M.; Shin, J.S.; Kim, K.S.; Gong, C.H.; Park, S.K.; Kim, J.S.; Yeom, S.C.; Hwang, E.S.; Lee, C.T.; Kim, S.J.; et al. Islet isolation from adult designated pathogen-free pigs: Use of the newer bovine nervous tissue-free enzymes and a revised donor selection strategy would improve the islet graft function. Xenotransplantation 2011, 18, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Gazda, L.S.; Collins, J.; Lovatt, A.; Holdcraft, R.W.; Morin, M.J.; Galbraith, D.; Graham, M.; Laramore, M.A.; Maclean, C.; Black, J.; et al. A comprehensive microbiological safety approach for agarose encapsulated porcine islets intended for clinical trials. Xenotransplantation 2016, 23, 444–463. [Google Scholar] [CrossRef] [PubMed]
- Beach, N.M.; Meng, X.J. Efficacy and future prospects of commercially available and experimental vaccines against porcine circovirus type 2 (PCV2). Virus Res. 2012, 164, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J. Best practice and future challenges for vaccination against porcine circovirus type 2. Expert Rev. Vaccines 2015, 14, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Madec, F.; Rose, N.; Grasland, B.; Cariolet, R.; Jestin, A. Post-weaning multisystemic wasting syndrome and other PCV2-related problems in pigs: A 12-year experience. Transbound Emerg. Dis. 2008, 55, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Glass, F. PCV2 vaccination changing the pig industry. Part 3. Reduced antibiotic usage and improved performance go together. Pig Progr. 2010, 26, 28–30. [Google Scholar]
- Fort, M.; Sibila, M.; Allepuz, A.; Mateu, E.; Roerink, F.; Segalés, J. Porcine circovirus type 2 (PCV2) vaccination of conventional pigs prevents viremia against PCV2 isolates of different genotypes and geographic origins. Vaccine 2008, 26, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Madson, D.M.; Patterson, A.R.; Ramamoorthy, S.; Pal, N.; Meng, X.J.; Opriessnig, T. Effect of porcine circovirus type 2 (PCV2) vaccination of the dam on PCV2 replication in utero. Clin. Vaccine Immunol. 2009, 16, 830–834. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.F.; Garrocho, F.M.; Lana, A.M.Q.; Lobato, Z.I.P. Fetal infections and antibody profiles in pigs naturally infected with porcine circovirus type 2 (PCV2). Can. J. Vet. Res. 2012, 76, 38–44. [Google Scholar] [PubMed]
- O’Neill, K.C.; Shen, H.G.; Lin, K.; Hemann, M.; Beach, N.M.; Meng, X.J.; Halbur, PG.; Opriessnig, T. Studies on porcine circovirus type 2 vaccination of 5-day-old piglets. Clin. Vaccine Immunol. 2011, 18, 1865–1871. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.W.; Han, K.; Park, C.; Chae, C. Clinical, virological, immunological and pathological evaluation of four porcine circovirus type 2 vaccines. Vet. J. 2014, 200, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Afghah, Z.; Webb, B.; Meng, X.J.; Ramamoorthy, S. Ten years of PCV2 vaccines and vaccination: Is eradication a possibility. Vet. Mic. 2016. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Xenotransplantation and porcine cytomegalovirus (PCMV). Xenotransplantation 2015, 22, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Mueller, N.J. Preventing transfer of infectious agents. Int. J. Surg. 2015, 23, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Kim, J.; Ha, Y.; Jung, K.; Choi, C.; Lim, J.K.; Kim, S.H.; Chae, C. Birth abnormalities in pregnant sows infected intranasally with porcine circovirus 2. J. Com. Pathol. 2005, 132, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Heinze, J.; Plotzki, E.; Denner, J. Virus safety of xenotranplantation: Prevalence of porcine circovirus 2 (PCV2) in pigs. Ann. Virol. Res. 2016, 2, 1023. [Google Scholar]
- Schuurman, H.J. Microbiological Denner, J. safety of clinical xenotransplantation products: Monitoring strategies and regulatory aspects. A commentary. Xenotransplantation 2016, 23, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Bielanski, A.; Algire, J.; Lalonde, A.; Garceac, A.; Pollard, J.W.; Plante, C. Nontransmission of porcine circovirus 2 (PCV2) by embryo transfer. Theriogenology 2013, 80, 77–83. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Vaccine | Producer | Vaccine Based on |
|---|---|---|
| Circumvent PCV, Porcilis PCV, Circumvent G2 PCV | MSD/Merck Animal Health (Madison, New Jersey, United States) | PCV2a Cap protein expressed by baculovirus |
| Ingelvac CircoFLEX | Boehringer-Ingelheim (St. Joseph, Missouri, United States) | PCV2a Cap protein expressed by baculovirus |
| Fostera PCV, Suvaxyn PCV | Zoetis (Parsippany, New Jersey, United States) | Inactivated recombinant PCV1 expressing the PCV2a Cap protein (ORF2 from PCV2) |
| Circovac | Merial (Lyon France) | Inactivated whole PCV2a |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denner, J.; Mankertz, A. Porcine Circoviruses and Xenotransplantation. Viruses 2017, 9, 83. https://doi.org/10.3390/v9040083
Denner J, Mankertz A. Porcine Circoviruses and Xenotransplantation. Viruses. 2017; 9(4):83. https://doi.org/10.3390/v9040083
Chicago/Turabian StyleDenner, Joachim, and Annette Mankertz. 2017. "Porcine Circoviruses and Xenotransplantation" Viruses 9, no. 4: 83. https://doi.org/10.3390/v9040083
APA StyleDenner, J., & Mankertz, A. (2017). Porcine Circoviruses and Xenotransplantation. Viruses, 9(4), 83. https://doi.org/10.3390/v9040083