
Nasal Infection of Enterovirus D68 Leading to Lower Respiratory Tract Pathogenesis in Ferrets (Mustela putorius furo)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Animal Experiments
2.2. Cells and Virus
2.3. Real-Time PCR Test for Viral Load Quantity
2.4. Histopathological and Immunohistochemical (IHC) Staining
2.5. Laser Confocal Microscopy Analysis of the Infected Ferret Lung Sections
2.6. Cytokine Quantification by Magnetic Beads-Based Bio-Plex Assay
2.7. Quantification of Cytokine mRNA
2.8. Neutralization Antibody Titer Test
2.9. Statistics
3. Results
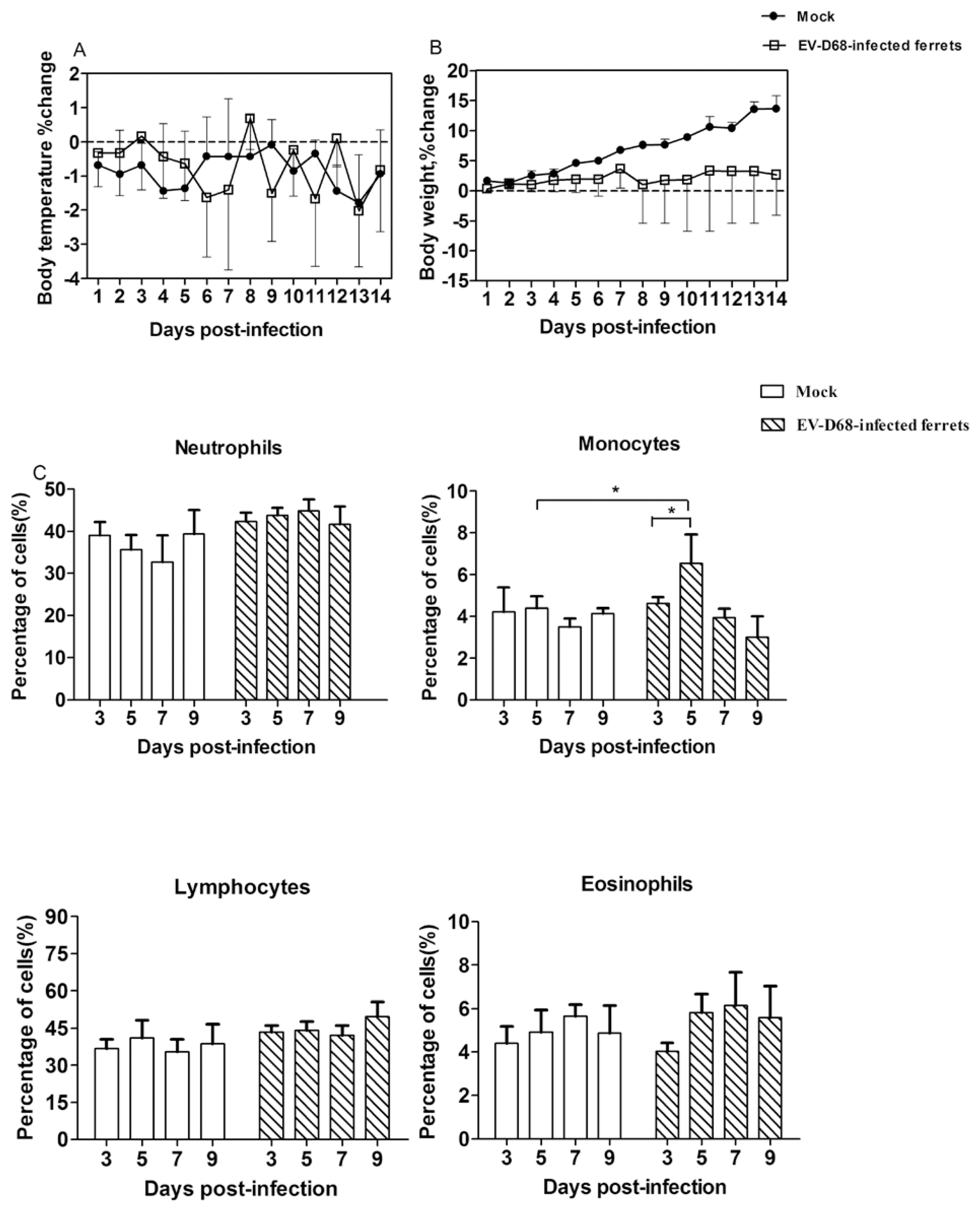
3.1. EV-D68 Infection in Ferrets Caused Normal, Cold-Like Clinical Signs
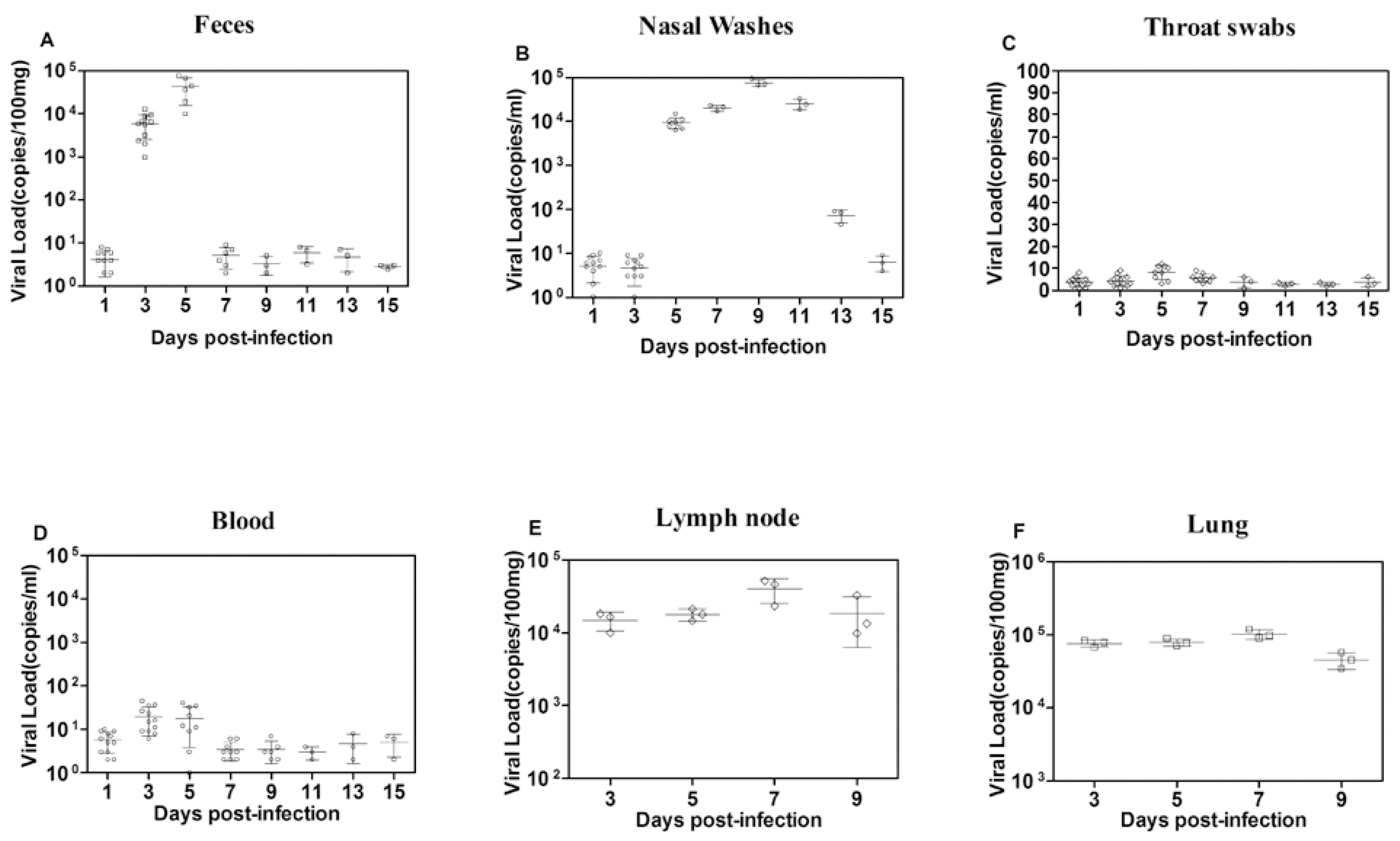
3.2. Virus Shedding Potential and Distribution in Different Tissues of EV-D68-Infected Ferrets
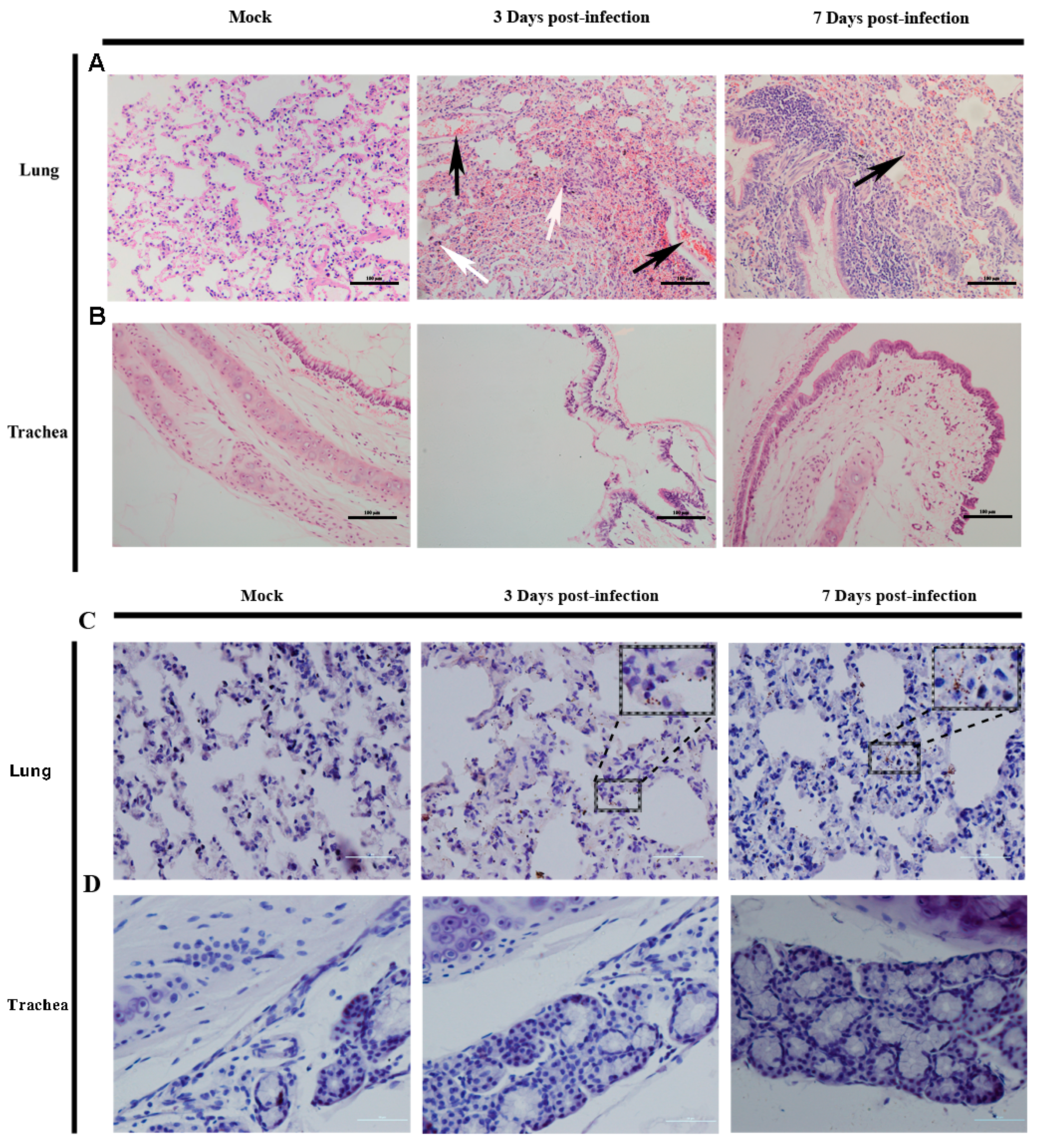
3.3. Histopathological Examinationand Immunohistochemical Analysis
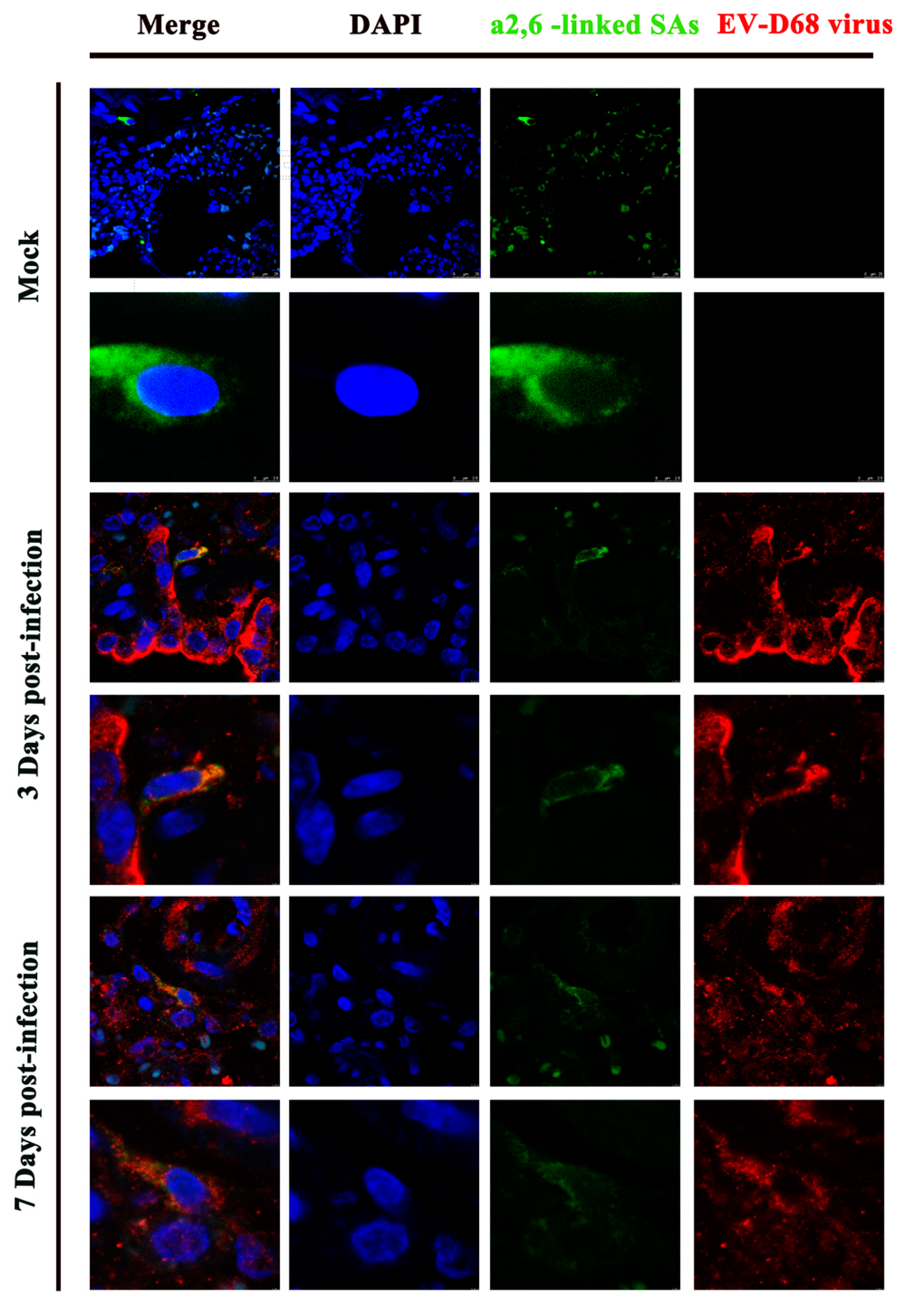
3.4. Confocal Imaging of EV-D68 VP1 with the α2,6-Linked SAs in the Lung Tissue of EV-D68-Infected Ferrets
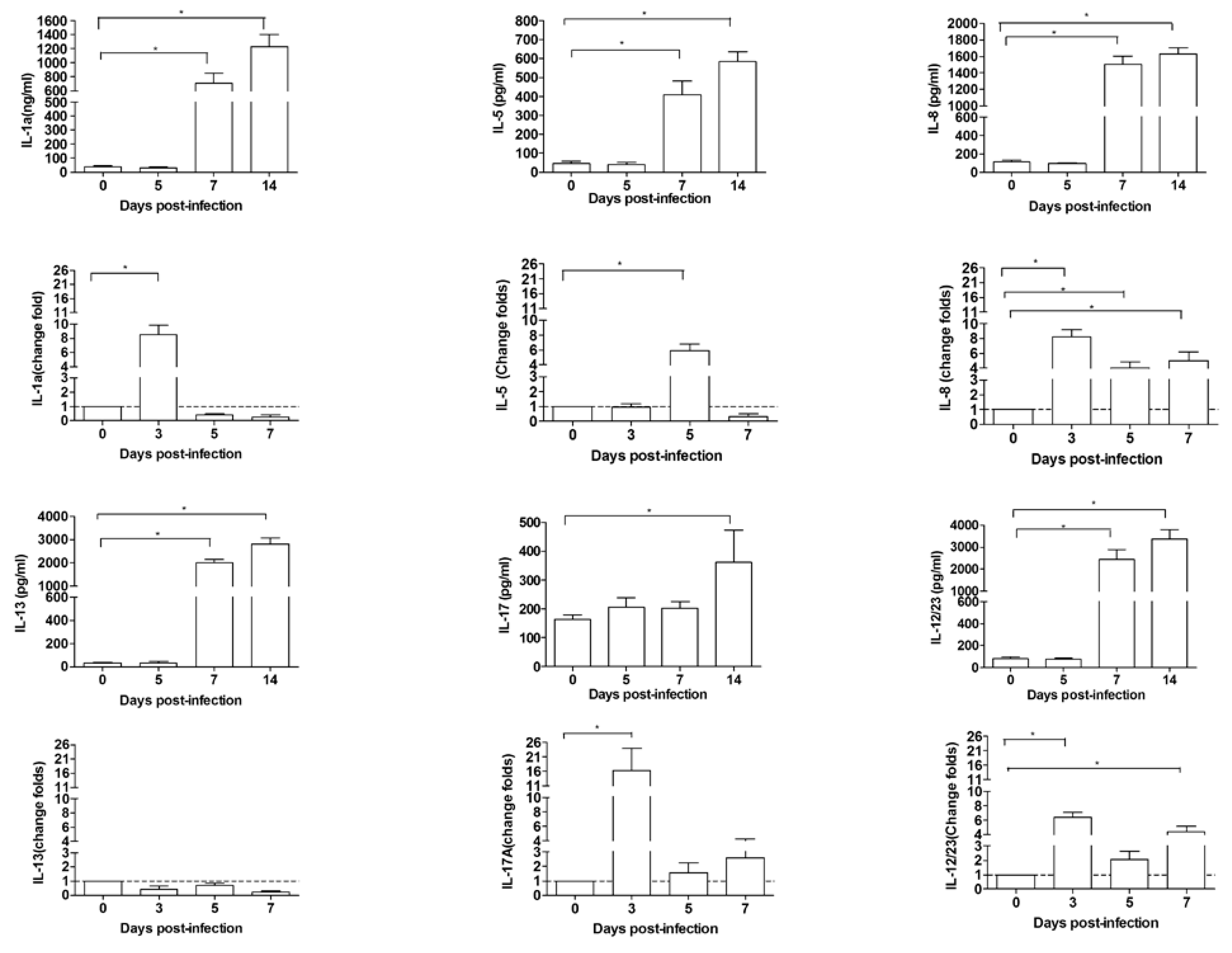
3.5. Inflammatory Cytokines Increased in Pulmonary Pathogenesis
3.6. Immunological Response of Ferrets after EV-D68 Infection
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tan, Y.; Hassan, F.; Schuster, J.E.; Simenauer, A.; Selvarangan, R.; Halpin, R.A.; Lin, X.; Fedorova, N.; Stockwell, T.B.; Lam, T.T.; et al. Molecular evolution and intraclade recombination of enterovirus D68 during the 2014 outbreak in the United States. J. Virol. 2015, 90, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, S.; Prifert, C.; Weissbrich, B.; Adams, O.; Aldabbagh, S.; Eis-Hubinger, A.M.; Diedrich, S. Detection of enterovirus D68 in patients hospitalised in three tertiary university hospitals in Germany, 2013 to 2014. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Farrell, J.J.; Ikladios, O.; Wylie, K.M.; O’Rourke, L.M.; Lowery, K.S.; Cromwell, J.S.; Wylie, T.N.; Melendez, E.L.; Makhoul, Y.; Sampath, R.; et al. Enterovirus D68-associated acute respiratory distress syndrome in adult, United States, 2014. Emerg. Infect. Dis. 2015, 21, 914–916. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Schreiner, T.L.; Maloney, J.A.; Wallace, A.; Ludke, J.; Oberste, M.S.; Nix, W.A.; Robinson, C.C.; Glodé, M.P.; Abzug, M.J.; et al. A cluster of acute flaccid paralysis and cranial nerve dysfunction temporally associated with an outbreak of enterovirus D68 in children in Colorado, USA. Lancet 2015, 385, 1662–1671. [Google Scholar] [CrossRef]
- Greninger, A.L.; Naccache, S.N.; Messacar, K.; Clayton, A.; Yu, G.; Somasekar, S.; Federman, S.; Stryke, D.; Anderson, C.; Yagi, S.; et al. A novel outbreak enterovirus D68 strain associated with acute flaccid myelitis cases in the USA (2012–2014): A retrospective cohort study. Lancet Infect. Dis. 2015, 15, 671–682. [Google Scholar] [CrossRef]
- Xiang, Z.; Liu, L.; Lei, X.; Zhou, Z.; He, B.; Wang, J. 3C protease of enterovirus D68 inhibits cellular defense mediated by interferon regulatory factor 7. J. Virol. 2015, 90, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Li, L.; Lei, X.; Zhou, H.; Zhou, Z.; He, B.; Wang, J. Enterovirus 68 3C protease cleaves TRIF to attenuate antiviral responses mediated by Toll-like receptor 3. J. Virol. 2014, 88, 6650–6659. [Google Scholar] [CrossRef] [PubMed]
- Holm-Hansen, C.C.; Midgley, S.E.; Fischer, T.K. Global emergence of enterovirus D68: A systematic review. Lancet Infect. Dis. 2016, 16, e64–e75. [Google Scholar] [CrossRef]
- Aliabadi, N.; Messacar, K.; Pastula, D.M.; Robinson, C.C.; Leshem, E.; Sejvar, J.J.; Nix, W.A.; Oberste, M.S.; Feikin, D.R.; Dominguez, S.R. Enterovirus D68 infection in children with acute flaccid myelitis, Colorado, USA, 2014. Emerg. Infect. Dis. 2016, 22, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Awano, H.; Ogi, M.; Tomioka, K.; Unzaki, A.; Nishiyama, M.; Toyoshima, D.; Taniguchi-Ikeda, M.; Ishida, A.; Nagase, H.; et al. A pediatric patient with interstitial pneumonia due to enterovirus D68. J. Infect. Chemother. 2016, 22, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sheng, J.; Baggen, J.; Meng, G.; Xiao, C.; Thibaut, H.J.; van Kuppeveld, F.J.; Rossmann, M.G. Sialic acid-dependent cell entry of human enterovirus D68. Nat. Commun. 2015, 6, 8865. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Okamoto, M.; Nakakita, S.; Suzuki, A.; Saito, M.; Tamaki, R.; Lupisan, S.; Roy, C.N.; Hiramatsu, H.; Sugawara, K.E.; et al. Antigenic and receptor binding properties of enterovirus 68. J. Virol. 2014, 88, 2374–2384. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.C.; Wang, W.; Pletneva, L.M.; Rajagopala, S.V.; Tan, Y.; Hartert, T.V.; Boukhvalova, M.S.; Vogel, S.N.; Das, S.R.; Blanco, J.C. Enterovirus D-68 infection, prophylaxis, and vaccination in a novel permissive animal model, the cotton rat (Sigmodon hispidus). PLoS ONE 2016, 11, e0166336. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Blixt, O.; Chen, L.M.; Pappas, C.; Maines, T.R.; van Hoeven, N.; Donis, R.; Busch, J.; McBride, R.; Paulson, J.C.; et al. Contemporary North American influenza H7 viruses possess human receptor specificity: Implications for virus transmissibility. Proc. Natl. Acad. Sci. USA 2008, 105, 7558–7563. [Google Scholar] [CrossRef] [PubMed]
- Matrosovich, M.N.; Gambaryan, A.S.; Teneberg, S.; Piskarev, V.E.; Yamnikova, S.S.; Lvov, D.K.; Robertson, J.S.; Karlsson, K.A. Avian influenza A viruses differ from human viruses by recognition of sialyloligosaccharides and gangliosides and by a higher coservation of the HA receptor-binding site. Virology 1997, 233, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Sutton, T.C.; Subbarao, K. Development of animal models against emerging coronaviruses: From SARS to MERS coronavirus. Virology 2015, 479–480, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Shinya, K.; Ebina, M.; Yamada, S.; Ono, M.; Kasai, N.; Kawaoka, Y. Avian flu: Influenza virus receptors in the human airway. Nature 2006, 440, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Leigh, M.W.; Connor, R.J.; Kelm, S.; Baum, L.G.; Paulson, J.C. Receptor specificity of Influenza virus influences severity of illness in ferrets. Vaccine 1995, 13, 1468–1473. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, H.; Zhang, Y.; Wang, J.; Che, Y.; Dong, C.; Zhang, X.; Na, R.; Shi, H.; Jiang, L.; et al. Neonatal rhesus monkey is a potential animal model for studying pathogenesis of EV71 infection. Virology 2011, 412, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Zhang, X.; Hu, Y.; Dong, C.; Liu, L.; Yang, E.; Che, Y.; Pu, J.; Wang, X.; et al. Pathologic and immunologic characteristics of coxsackievirus A16 infection in rhesus macaques. Virology 2017, 500, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Eash, S.; Tavares, R.; Stopa, E.G.; Robbins, S.H.; Brossay, L.; Atwood, W.J. Differential distribution of the JC virus receptor-type sialic acid in normal human tissues. Am. J. Pathol. 2004, 164, 419–428. [Google Scholar] [CrossRef]
- Carolan, L.A.; Butler, J.; Rockman, S.; Guarnaccia, T.; Hurt, A.C.; Reading, P.; Kelso, A.; Barr, I.; Laurie, K.L. TaqMan real time RT-PCR assays for detecting ferret innate and adaptive immune responses. J. Virol. Methods 2014, 205, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Rowe, T.; Leon, A.J.; Banner, D.; Danesh, A.; Xu, L.; Ran, L.; Bosinger, S.E.; Guan, Y.; Chen, H.; et al. Molecular characterization of in vivo adjuvant activity in ferrets vaccinated against influenza virus. J. Virol. 2010, 84, 8369–8388. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ma, Y.; Xu, Y.; Yang, H.; Shi, L.; Che, Y.; Liao, G.; Jiang, S.; Zhang, S.; Li, Q. Dynamic profiles of neutralizing antibody responses elicited in rhesus monkeys immunized with a combined tetravalent DTaP-Sabin IPV candidate vaccine. Vaccine 2014, 32, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Moore, D.D.; Nix, W.A.; Oberste, M.S.; Weldon, W.C. Neutralization of Enterovirus D68 isolated from the 2014 US outbreak by commercial intravenous immune globulin products. J. Clin. Virol. 2015, 69, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Linsuwanon, P.; Puenpa, J.; Suwannakarn, K.; Auksornkitti, V.; Vichiwattana, P.; Korkong, S.; Theamboonlers, A.; Poovorawan, Y. Molecular epidemiology and evolution of human enterovirus serotype 68 in Thailand 2006–2011. PLoS ONE 2012, 7, e35190. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Perez, J.A.; Ramirez-Gonzalez, J.E.; Moreno-Valencia, Y.; Hernandez-Hernandez, V.A.; Romero-Espinoza, J.A.; Castillejos-Lopez, M.; Hernandez, A.; Perez-Padilla, R.; Oropeza-Lopez, L.E.; Escobar-Escamilla, N.; et al. EV-D68 infection in children with asthma exacerbation and pneumonia in Mexico City during 2014 autumn. Influenza Other Respir. Viruses 2016, 10, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Chidini, G.; Cinnante, C.; Napolitano, L.; Giannini, A.; Terranova, L.; Niesters, H.; Principi, N.; Calderini, E. Acute flaccid myelitis associated with enterovirus-D68 infection in an otherwise healthy child. Virol. J. 2017, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Smura, T.; Ylipaasto, P.; Klemola, P.; Kaijalainen, S.; Kyllonen, L.; Sordi, V.; Piemonti, L.; Roivainen, M. Cellular tropism of human enterovirus D species serotypes EV-94, EV-70, and EV-68 in vitro: Implications for pathogenesis. J. Med. Virol. 2010, 82, 1940–1949. [Google Scholar] [CrossRef] [PubMed]
- Thongpan, I.; Wanlapakorn, N.; Vongpunsawad, S.; Linsuwanon, P.; Theamboonlers, A.; Payungporn, S.; Poovorawan, Y. Prevalence and phylogenetic characterization of enterovirus D68 in pediatric patients with acute respiratory tract infection in Thailand. Jpn. J. Infect. Dis. 2016, 69, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Enterovirus D68: Background, Pathophysiology, Etiology. Available online: http://emedicine.medscape.com/article/2236902-overview (accessed on 11 January 2016).
- Schieble, J.H.; Fox, V.L.; Lennette, E.H. A probable new human picornavirus associated with respiratory diseases. Am. J. Epidemiol. 1967, 85, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Ward, N.S.; Hughes, B.L.; Mermel, L.A. Enterovirus D68 infection in an adult. Am. J. Crit. Care 2016, 25, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Uncapher, C.R.; Dewitt, C.M.; Colonno, R.J. The major and minor group receptor families contain all but one human rhinovirus serotype. Virology 1991, 180, 814–817. [Google Scholar] [CrossRef]
- Martel, C.J.; Aasted, B. Characterization of antibodies against ferret immunoglobulins, cytokines and CD markers. Vet. Immunol. Immunopathol. 2009, 132, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, L.G.; Castelruiz, Y.; Jacobsen, S.; Aasted, B. Identification of monoclonal antibodies that cross-react with cytokins from different animal species. Vet. Immunol. Immunopathol. 2002, 88, 111–112. [Google Scholar] [CrossRef]
- Camp, J.V.; Bagci, U.; Chu, Y.K.; Squier, B.; Fraig, M.; Uriarte, S.M.; Guo, H.; Mollura, D.J.; Jonsson, C.B. Lower respiratory tract infection of the ferret by 2009 H1N1 pandemic influenza A virus triggers biphasic, systemic, and local recruitment of neutrophils. J. Virol. 2015, 89, 8733–8748. [Google Scholar] [CrossRef] [PubMed]
- Gustin, K.M.; Belser, J.A.; Veguilla, V.; Zeng, H.; Katz, J.M.; Tumpey, T.M.; Maines, T.R. Environmental conditions affect exhalation of H3N2 seasonal and variant influenza viruses and respiratory droplet transmission in ferrets. PLoS ONE 2015, 10, e0125874. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Banner, D.; Paquette, S.G.; Leon, A.J.; Kelvin, A.A.; Kelvin, D.J. Pathogenic influenza B virus in the ferret model establishes lower respiratory tract infection. J. Gen. Virol. 2014, 95, 2127–2139. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Oshitani, H. Global reemergence of enterovirus D68 as an important pathogen for acute respiratory infections. Rev. Med. Virol. 2015, 25, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Ren, L.; Zheng, S.; Wang, L.; Xie, X.; Deng, Y.; Zhao, Y.; Zhao, X.; Luo, Z.; Fu, Z.; et al. Prevalence and molecular characterizations of enterovirus D68 among children with acute respiratory infection in China between 2012 and 2014. Sci. Rep. 2015, 5, 16639. [Google Scholar] [CrossRef] [PubMed]
- Orvedahl, A.; Padhye, A.; Barton, K.; O’Bryan, K.; Baty, J.; Gruchala, N.; Niesen, A.; Margoni, A.; Srinivasan, M. Clinical characterization of children presenting to the hospital with enterovirus D68 infection during the 2014 outbreak in St. Louis. Pediatr. Infect. Dis. J. 2016, 35, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Suzuki, A.; Lupisan, S.; Okamoto, M.; Aniceto, R.; Egos, R.J.; Daya, E.E.; Tamaki, R.; Saito, M.; Fuji, N.; et al. Molecular evolution of enterovirus 68 detected in the Philippines. PLoS ONE 2013, 8, e74221. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.-W.; Sun, M.; Guo, L.; Wang, J.-J.; Song, J.; Li, J.-Q.; Li, H.-Z.; Ning, R.-T.; Yang, Z.-N.; Fan, H.-T.; et al. Nasal Infection of Enterovirus D68 Leading to Lower Respiratory Tract Pathogenesis in Ferrets (Mustela putorius furo). Viruses 2017, 9, 104. https://doi.org/10.3390/v9050104
Zheng H-W, Sun M, Guo L, Wang J-J, Song J, Li J-Q, Li H-Z, Ning R-T, Yang Z-N, Fan H-T, et al. Nasal Infection of Enterovirus D68 Leading to Lower Respiratory Tract Pathogenesis in Ferrets (Mustela putorius furo). Viruses. 2017; 9(5):104. https://doi.org/10.3390/v9050104
Chicago/Turabian StyleZheng, Hui-Wen, Ming Sun, Lei Guo, Jing-Jing Wang, Jie Song, Jia-Qi Li, Hong-Zhe Li, Ruo-Tong Ning, Ze-Ning Yang, Hai-Tao Fan, and et al. 2017. "Nasal Infection of Enterovirus D68 Leading to Lower Respiratory Tract Pathogenesis in Ferrets (Mustela putorius furo)" Viruses 9, no. 5: 104. https://doi.org/10.3390/v9050104
APA StyleZheng, H.-W., Sun, M., Guo, L., Wang, J.-J., Song, J., Li, J.-Q., Li, H.-Z., Ning, R.-T., Yang, Z.-N., Fan, H.-T., He, Z.-L., & Liu, L.-D. (2017). Nasal Infection of Enterovirus D68 Leading to Lower Respiratory Tract Pathogenesis in Ferrets (Mustela putorius furo). Viruses, 9(5), 104. https://doi.org/10.3390/v9050104