
A Motif in the F Homomorph of Rabbit Haemorrhagic Disease Virus Polymerase Is Important for the Subcellular Localisation of the Protein and Its Ability to Induce Redistribution of Golgi Membranes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Plasmids
2.3. Reference Sequences
2.4. Antibodies
2.5. Immunofluorescence
2.6. Cell Counting
2.7. Hydrophobicity Scales, Prediction and Protein Structure Analysis
2.8. Molecular Dynamics
3. Results
3.1. Subcellular Localisation of Recombinant Calicivirus RdRps
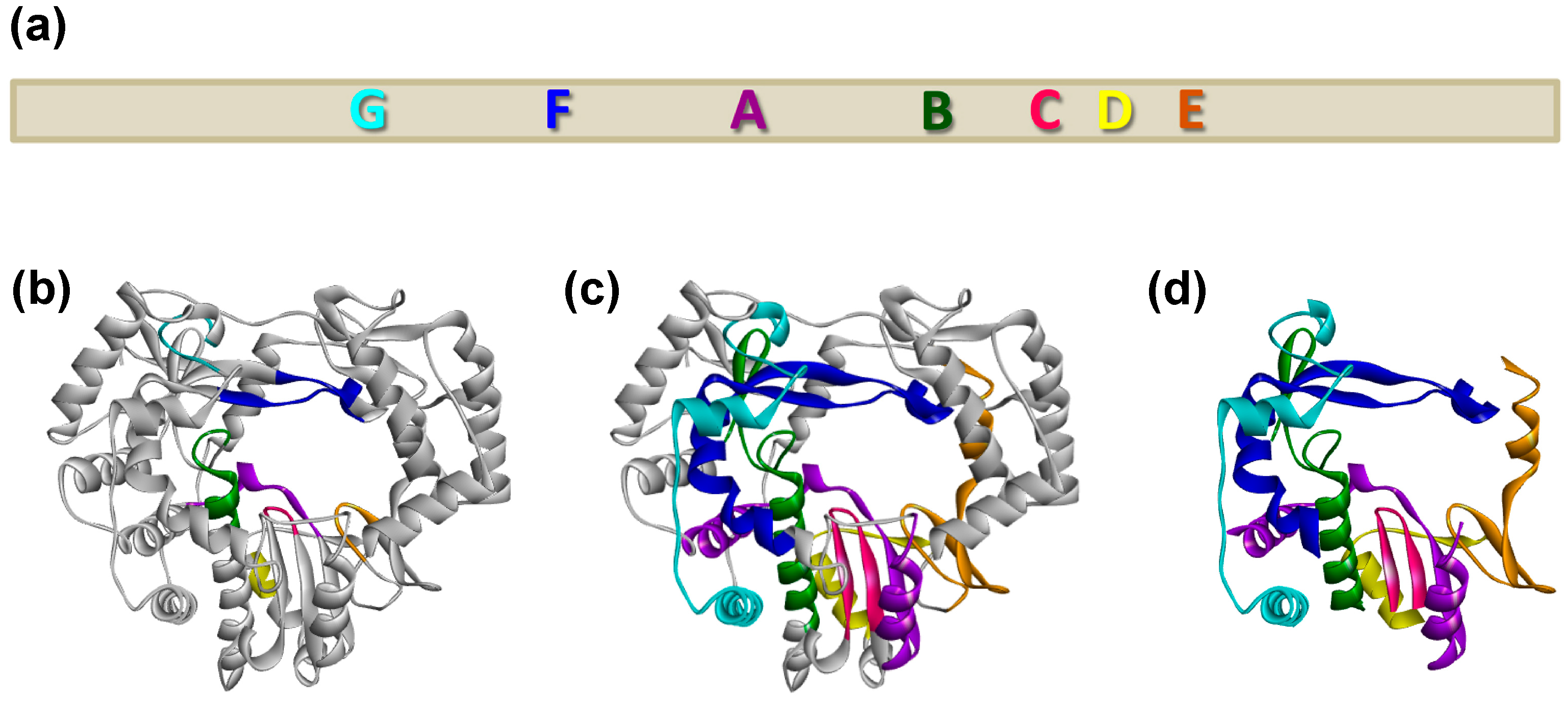
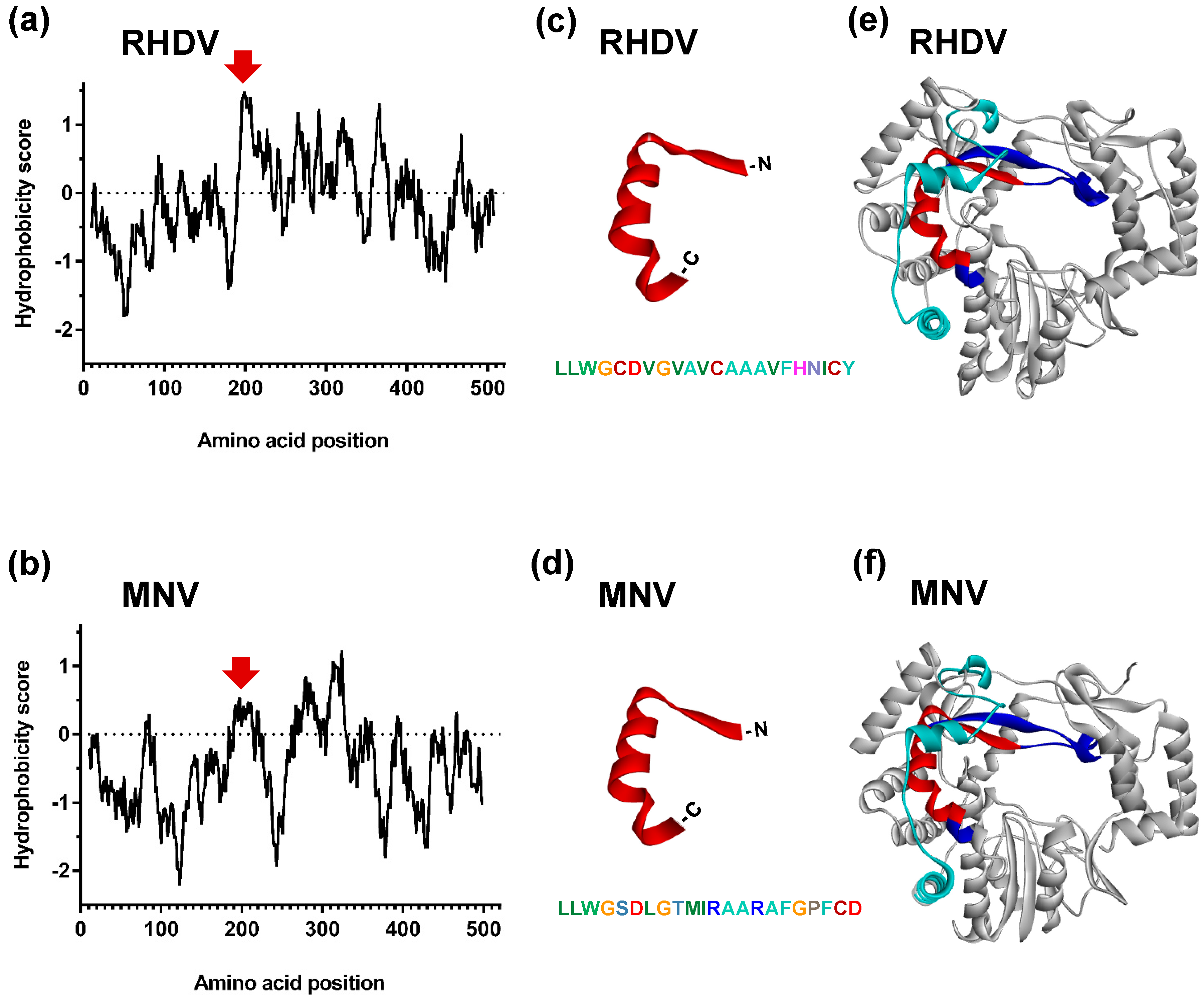
3.2. Identification of a Putative Motif That Is Important for the Subcellular Localisation of RHDV RdRp
3.3. Role of the Proposed Hydrophobic Motif in Regulating the Subcellular Localisation of RHDV RdRp
3.4. Amino Acid Substitutions Decreasing the Hydrophobicity of the Putative Interacting Motif Change the Subcellular Localisation of RHDV RdRp and Reduce Its Ability to Induce Redistribution of the Golgi Membranes
3.5. Molecular Dynamics
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Abrantes, J.; van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and Rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef] [PubMed]
- Thiel, H.-J.; König, M. Caliciviruses: An overview. Vet. Microbiol. 1999, 69, 55–62. [Google Scholar] [CrossRef]
- Clarke, I.N.; Lambden, P.R. The molecular biology of caliciviruses. J. Gen. Virol. 1997, 78, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.-J. Genomic and subgenomic RNAs of rabbit hemorrhagic disease virus are both protein-linked and packaged into particles. Virology 1991, 184, 677–686. [Google Scholar] [CrossRef]
- Meyers, G.; Wirblich, C.; Thiel, H.-J. Rabbit hemorrhagic disease virus—Molecular cloning and nucleotide sequencing of a calicivirus genome. Virology 1991, 184, 664–676. [Google Scholar] [CrossRef]
- Wirblich, C.; Thiel, H.-J.; Meyers, G. Genetic map of the calicivirus rabbit hemorrhagic disease virus as deduced from in vitro translation studies. J. Virol. 1996, 70, 7974–7983. [Google Scholar] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.-J.; Thumfart, J.O. Rabbit hemorrhagic disease virus: Genome organization and polyprotein processing of a calicivirus studied after transient expression of cDNA constructs. Virology 2000, 276, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.H. Viral polymerases. Adv. Exp. Med. Biol. 2012, 726, 267–304. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Orta, C.; Arias, A.; Escarmís, C.; Verdaguer, N. A comparison of viral RNA-dependent RNA polymerases. Curr. Opin. Struct. Biol. 2006, 16, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.K.S.; Cherney, M.M.; Vázquez, A.L.; Machín, Á.; Alonso, J.M.M.; Parra, F.; James, M.N. Crystal structures of active and inactive conformations of a caliciviral RNA-dependent RNA polymerase. J. Biol. Chem. 2002, 277, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, S.W.B.; Blaschke, M.; Coutard, B.; Gebhardt, J.; Gorbalenya, A.; Canard, B.; Tucker, P.A.; Rohayem, J. Structural and functional characterization of sapovirus RNA-dependent RNA polymerase. J. Virol. 2007, 81, 1858–1871. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.K.-S.; Pendás-Franco, N.; Rojo, J.; Boga, J.A.; Machín, Á.; Alonso, J.M.M.; Parra, F. Crystal structure of Norwalk virus polymerase reveals the carboxyl terminus in the active site cleft. J. Biol. Chem. 2004, 279, 16638–16645. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Alam, I.; Han, K.R.; Cho, S.; Shin, S.; Kang, S.; Yang, J.M.; Kim, K.H. Crystal structures of murine norovirus-1 RNA-dependent RNA polymerase. J. Gen. Virol. 2011, 92, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Te Velthuis, A.J.W. Common and unique features of viral RNA-dependent polymerases. Cell. Mol. Life Sci. 2014, 71, 4403–4420. [Google Scholar] [CrossRef] [PubMed]
- Jácome, R.; Becerra, A.; de León, S.P.; Lazcano, A. Structural analysis of monomeric RNA-dependent polymerases: Evolutionary and therapeutic implications. PLoS ONE 2015, 10, e0139001. [Google Scholar] [CrossRef] [PubMed]
- Bruenn, J.A. A structural and primary sequence comparison of the viral RNA-dependent RNA polymerases. Nucleic Acids Res. 2003, 31, 1821–1829. [Google Scholar] [CrossRef] [PubMed]
- Lang, D.M.; Zemla, A.T.; Zhou, C.L.E. Highly similar structural frames link the template tunnel and NTP entry tunnel to the exterior surface in RNA-dependent RNA polymerases. Nucleic Acids Res. 2013, 41, 1464–1482. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.K.-S.; Arnold, J.J.; Cameron, C.E. Structure-function relationships among RNA-dependent RNA polymerases. Curr. Top. Microbiol. Immunol. 2008, 320, 137–156. [Google Scholar] [PubMed]
- Pan, J.; Vakharia, V.N.; Tao, Y.J. The structure of a birnavirus polymerase reveals a distinct active site topology. Proc. Natl. Acad. Sci. USA 2007, 104, 7385–7390. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Pringle, F.M.; Zeddam, J.-L.; Luke, B.T.; Cameron, C.E.; Kalmakoff, J.; Hanzlik, T.N.; Gordon, K.H.; Ward, V.K. The palm subdomain-based active site is internally permuted in viral RNA-dependent RNA polymerases of an ancient lineage. J. Mol. Biol. 2002, 324, 47–62. [Google Scholar] [CrossRef]
- Černý, J.; Bolfíková, B.Č.; Valdes, J.J.; Grubhoffer, L.; Růžek, D. Evolution of tertiary structure of viral RNA dependent polymerases. PLoS ONE 2014, 9, e96070. [Google Scholar] [CrossRef] [PubMed]
- Hyde, J.L.; Mackenzie, J.M. Subcellular localization of the MNV-1 ORF1 proteins and their potential roles in the formation of the MNV-1 replication complex. Virology 2010, 406, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Brockway, S.M.; Clay, C.T.; Lu, X.T.; Denison, M.R. Characterization of the expression, intracellular localization, and replication complex association of the putative mouse hepatitis virus RNA-dependent RNA polymerase. J. Virol. 2003, 77, 10515–10527. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.B.; Park, K.-J.; Kim, Y.-S.; Sung, Y.C.; Lai, M.M.C. Hepatitis C virus NS5B protein is a membrane-associated phosphoprotein with a predominantly perinuclear localization. Virology 1997, 227, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.M.; Kenney, M.T.; Westaway, E.G. West Nile virus strain Kunjin NS5 polymerase is a phosphoprotein localized at the cytoplasmic site of viral RNA synthesis. J. Gen. Virol. 2007, 88, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Raychaudhuri, S.; Dasgupta, A. Nuclear entry of poliovirus protease-polymerase precursor 3CD: Implications for host cell transcription shut-off. Virology 2004, 320, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Urakova, N.; Frese, M.; Hall, R.N.; Liu, J.; Matthaei, M.; Strive, T. Expression and partial characterisation of Rabbit haemorrhagic disease virus non-structural proteins. Virology 2015, 484, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Urakova, N.; Strive, T.; Frese, M. RNA-dependent RNA polymerases of both virulent and benign rabbit caliciviruses induce striking rearrangement of Golgi membranes. PLoS ONE 2017, 12, e0169913. [Google Scholar] [CrossRef] [PubMed]
- Eltahla, A.A.; Lim, K.L.; Eden, J.-S.; Kelly, A.G.; Mackenzie, J.M.; White, P.A. Nonnucleoside inhibitors of norovirus RNA polymerase: Scaffolds for rational drug design. Antimicrob. Agents Chemother. 2014, 58, 3115–3123. [Google Scholar] [CrossRef] [PubMed]
- Bull, R.A.; Hyde, J.; Mackenzie, J.M.; Hansman, G.S.; Oka, T.; Takeda, N.; White, P.A. Comparison of the replication properties of murine and human calicivirus RNA-dependent RNA polymerases. Virus Genes 2011, 42, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.L.; Hewitt, J.; Sitabkhan, A.; Eden, J.-S.; Lun, J.; Levy, A.; Merif, J.; Smith, D.; Rawlinson, W.D.; White, P.A. A multi-site study of norovirus molecular epidemiology in Australia and New Zealand, 2013-2014. PLoS ONE 2016, 11, e0145254. [Google Scholar] [CrossRef] [PubMed]
- Urakova, N.; Netzler, N.; Kelly, A.G.; Frese, M.; White, P.A.; Strive, T. Purification and biochemical characterisation of rabbit calicivirus RNA-dependent RNA polymerases and identification of non-nucleoside inhibitors. Viruses 2016, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Snider, C.; Jayasinghe, S.; Hristova, K.; White, S.H. MPEx: A tool for exploring membrane proteins. Protein Sci. 2009, 18, 2624–2628. [Google Scholar] [CrossRef] [PubMed]
- Deber, C.M.; Wang, C.; Liu, L.-P.; Prior, A.S.; Agrawal, S.; Muskat, B.L.; Cuticchia, A.J. TM Finder: A prediction program for transmembrane protein segments using a combination of hydrophobicity and nonpolar phase helicity scales. Protein Sci. 2001, 10, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, K.; Stoffel, W. TMbase—A database of membrane spanning protein segments. Biol. Chem. Hoppe-Seyler 1993, 374, 166. [Google Scholar]
- Case, D.A.; Cerutti, D.S.; Cheatham, T.E., III; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; Greene, D.; Homeyer, N.; et al. AMBER 2017; University of California: San Francisco, CA, USA, 2017; Available online: http://ambermd.org/ (accessed on 29 July 2017).
- Ryckaert, J.-P.; Ciccotti, G.; Berendsen, H.J.C. Numerical integration of the Cartesian equations of motion of a system with constraints: Molecular Dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Antonny, B. Mechanisms of membrane curvature sensing. Annu. Rev. Biochem. 2011, 80, 101–123. [Google Scholar] [CrossRef] [PubMed]
- McMahon, H.T.; Boucrot, E. Membrane curvature at a glance. J. Cell Sci. 2015, 128, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Blobel, G. Intracellular protein topogenesis. Proc. Natl. Acad. Sci. USA 1980, 77, 1496–1500. [Google Scholar] [CrossRef] [PubMed]
- Whited, A.M.; Johs, A. The interactions of peripheral membrane proteins with biological membranes. Chem. Phys. Lipids 2015, 192, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; London, E. An amino acid “transmembrane tendency” scale that approaches the theoretical limit to accuracy for prediction of transmembrane helices: Relationship to biological hydrophobicity. Protein Sci. 2006, 15, 1987–2001. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.; Rath, A.; Johnson, R.M.; Deber, C.M. Distinctions between hydrophobic helices in globular proteins and transmembrane segments as factors in protein sorting. J. Biol. Chem. 2009, 284, 5395–5402. [Google Scholar] [CrossRef] [PubMed]
- Spuul, P.; Salonen, A.; Merits, A.; Jokitalo, E.; Kääriäinen, L.; Ahola, T. Role of the amphipathic peptide of Semliki forest virus replicase protein nsP1 in membrane association and virus replication. J. Virol. 2007, 81, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Orta, C.; de la Higuera, I.; Caridi, F.; Sánchez-Aparicio, M.T.; Moreno, E.; Perales, C.; Singh, K.; Sarafianos, S.G.; Sobrino, F.; Domingo, E.; et al. Multifunctionality of a picornavirus polymerase domain: Nuclear localization signal and nucleotide recognition. J. Virol. 2015, 89, 6848–6859. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Aparicio, M.T.; Rosas, M.F.; Sobrino, F. Characterization of a nuclear localization signal in the foot-and-mouth disease virus polymerase. Virology 2013, 444, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-C.; Kuo, R.-L.; Lin, J.-Y.; Huang, P.-N.; Huang, Y.; Liu, H.; Arnold, J.J.; Chen, S.J.; Wang, R.Y.; Cameron, C.E.; et al. Cytoplasmic viral RNA-dependent RNA polymerase disrupts the intracellular splicing machinery by entering the nucleus and interfering with Prp8. PLoS Pathog. 2014, 10, e1004199. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vector | Construct | Primer Sequence (5′→3′) |
|---|---|---|
| pcDNA 3.1/myc-His(-)C | pRHDV-RdRp-Nter | F: ATCGTTATAGCGGCCGCCTGCAGTAGCCACCATGACGTCAAACTTCTTCTGTGG R: TATGGGATCCTCTTCTTTCCCTCCTTCACCTTGTCAAG |
| pRHDV-RdRp-Cter | F: TATGGCGGCCGCGCCACCATGTACGAACTAAAGATGGTCGCGCGG R: ATCGTCATCGGATCCAGATATCCTCCATAACATTCACAAATTCGTC | |
| pRHDV-RdRp-Cter-PIM | F: TATGGGATCCTAGCCACCATGCTTGACAAGGTGAAGGAGGGAAAGAAG R: ATCGTCATCGGATCCAGATATCCTCCATAACATTCACAAATTCGTC | |
| pRHDV-FAA-PIM-FAA:myc | F: TATGGGATCCTAGCCACCATGCTTGACAAGGTGAAGGAGGGAAAGAAG R: TATCAAGCTTCGGGGCCAAACCGCGCGACCATC | |
| pMNV-RdRp | F: TATGGCGGCCGCCACCATGCTTCCCCGCCCCTC R: TATGGGATCCTCTCATCCTCATTCACAAAG | |
| pSaV-RdRp | F: TATGGCGGCCGCCACCATGGATGAATTCCAATGGAAGGG R: TATGGGATCCTCTCCATCTCAAACACTATTTTG | |
| pNoV-RdRp | F: TATGGCGGCCGCCACCATGGGTGGTGACAGTAAGGG R: TATGGGATCCTTTCGACGCCATCTTCATTC | |
| pFCV-RdRp | F: TATGGCGGCCGCCACCATGGTGACAGCCCAGAAATATGATG R: TATGGGATCCTCACTTCAAACACATCACAATGC | |
| pEGFP-N1 | pRHDV-FAA-PIM-FAA:GFP | F: TATGAAGCTTTAGCCACCATGCTTGACAAGGTGAAGGAGGGAAAGAAG R: TATCGGATCCTCGGGGCCAAACCGCGCGACCATC |
| pcDNA 3.1/myc-His(-)C | pRHDV-V195S-V197S | F: CCGTGTGTGCTGCCGCAGTTTTC R: CGGAACCAGAGTCACAGCC |
| pRHDV-V199S-V204S | F: CCTCGTGTGCTGCCGCATCTTTC R: CGACACCAACGTCACAGCC | |
| pRHDV-V195S-V197S-V199S-V204S | F: CCTCGTGTGCTGCCGCATCTTTC R: CGGAACCAGAGTCACAGCC |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urakova, N.; Warden, A.C.; White, P.A.; Strive, T.; Frese, M. A Motif in the F Homomorph of Rabbit Haemorrhagic Disease Virus Polymerase Is Important for the Subcellular Localisation of the Protein and Its Ability to Induce Redistribution of Golgi Membranes. Viruses 2017, 9, 202. https://doi.org/10.3390/v9080202
Urakova N, Warden AC, White PA, Strive T, Frese M. A Motif in the F Homomorph of Rabbit Haemorrhagic Disease Virus Polymerase Is Important for the Subcellular Localisation of the Protein and Its Ability to Induce Redistribution of Golgi Membranes. Viruses. 2017; 9(8):202. https://doi.org/10.3390/v9080202
Chicago/Turabian StyleUrakova, Nadya, Andrew C. Warden, Peter A. White, Tanja Strive, and Michael Frese. 2017. "A Motif in the F Homomorph of Rabbit Haemorrhagic Disease Virus Polymerase Is Important for the Subcellular Localisation of the Protein and Its Ability to Induce Redistribution of Golgi Membranes" Viruses 9, no. 8: 202. https://doi.org/10.3390/v9080202
APA StyleUrakova, N., Warden, A. C., White, P. A., Strive, T., & Frese, M. (2017). A Motif in the F Homomorph of Rabbit Haemorrhagic Disease Virus Polymerase Is Important for the Subcellular Localisation of the Protein and Its Ability to Induce Redistribution of Golgi Membranes. Viruses, 9(8), 202. https://doi.org/10.3390/v9080202