Rotavirus VP6 as an Adjuvant for Bivalent Norovirus Vaccine Produced in Nicotiana benthamiana

Abstract
:1. Introduction
2. Materials and Methods
2.1. Recombinant Proteins
2.2. Immunization of Mice
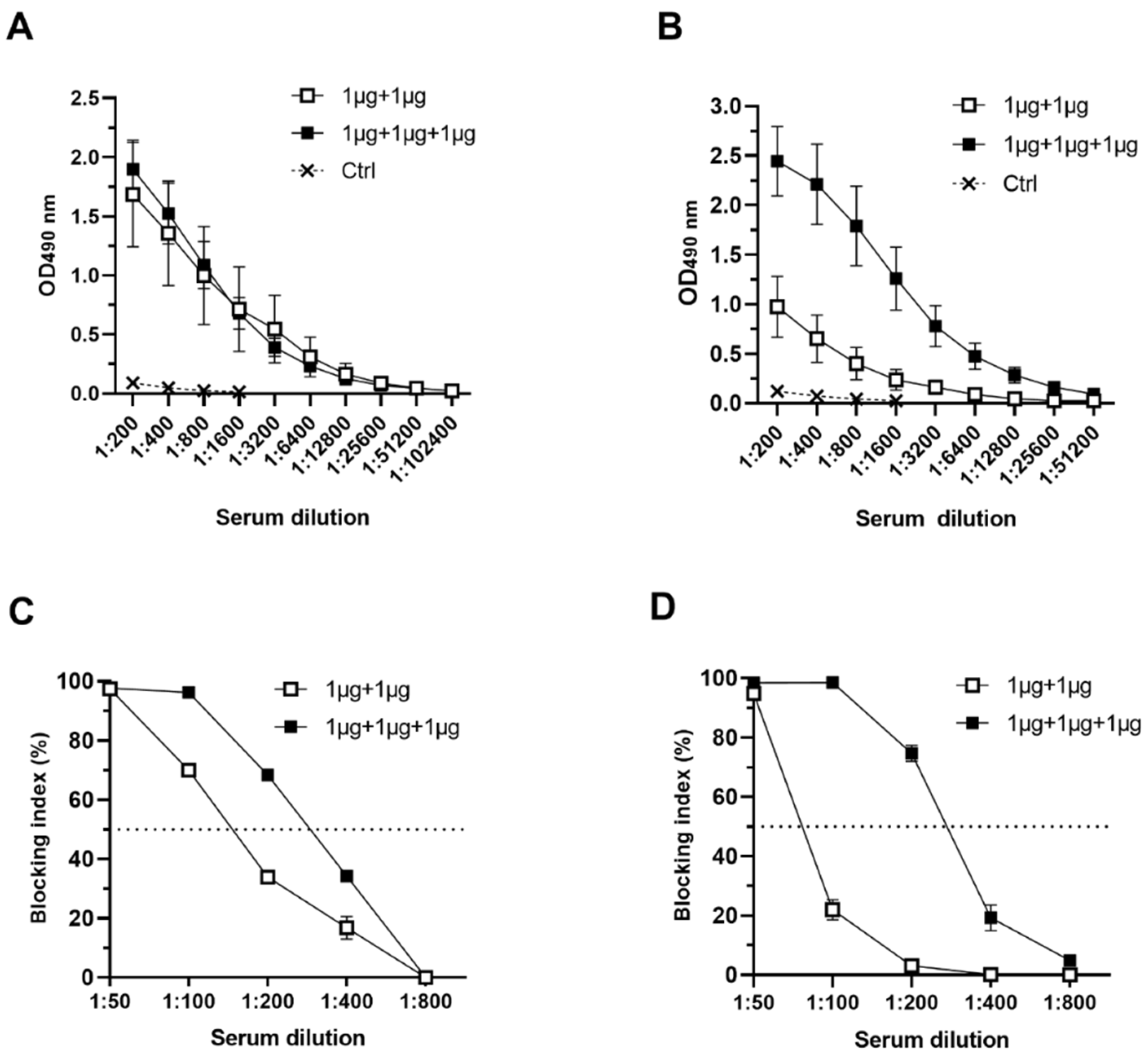
2.3. Serum IgG, IgG1, and IgG2a ELISA
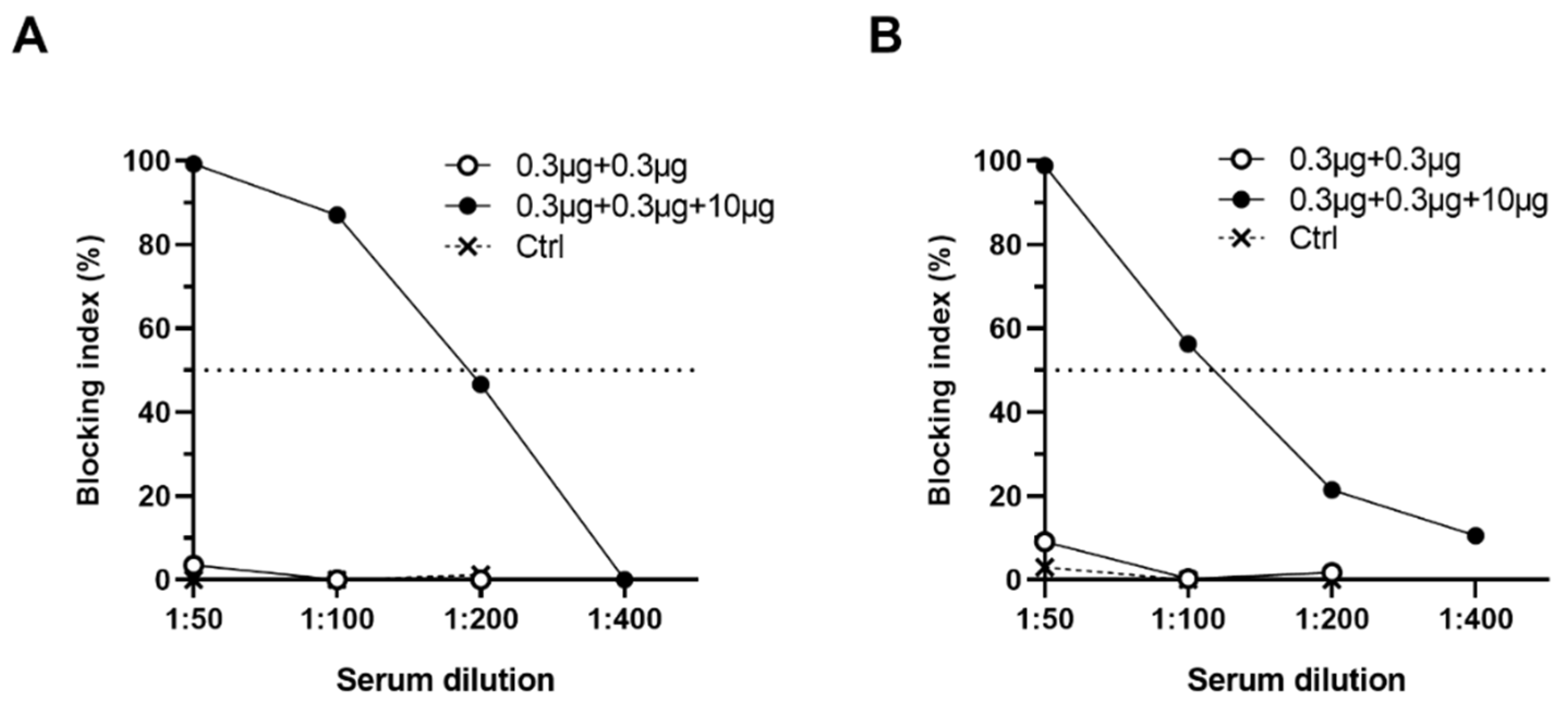
2.4. Blocking Assays
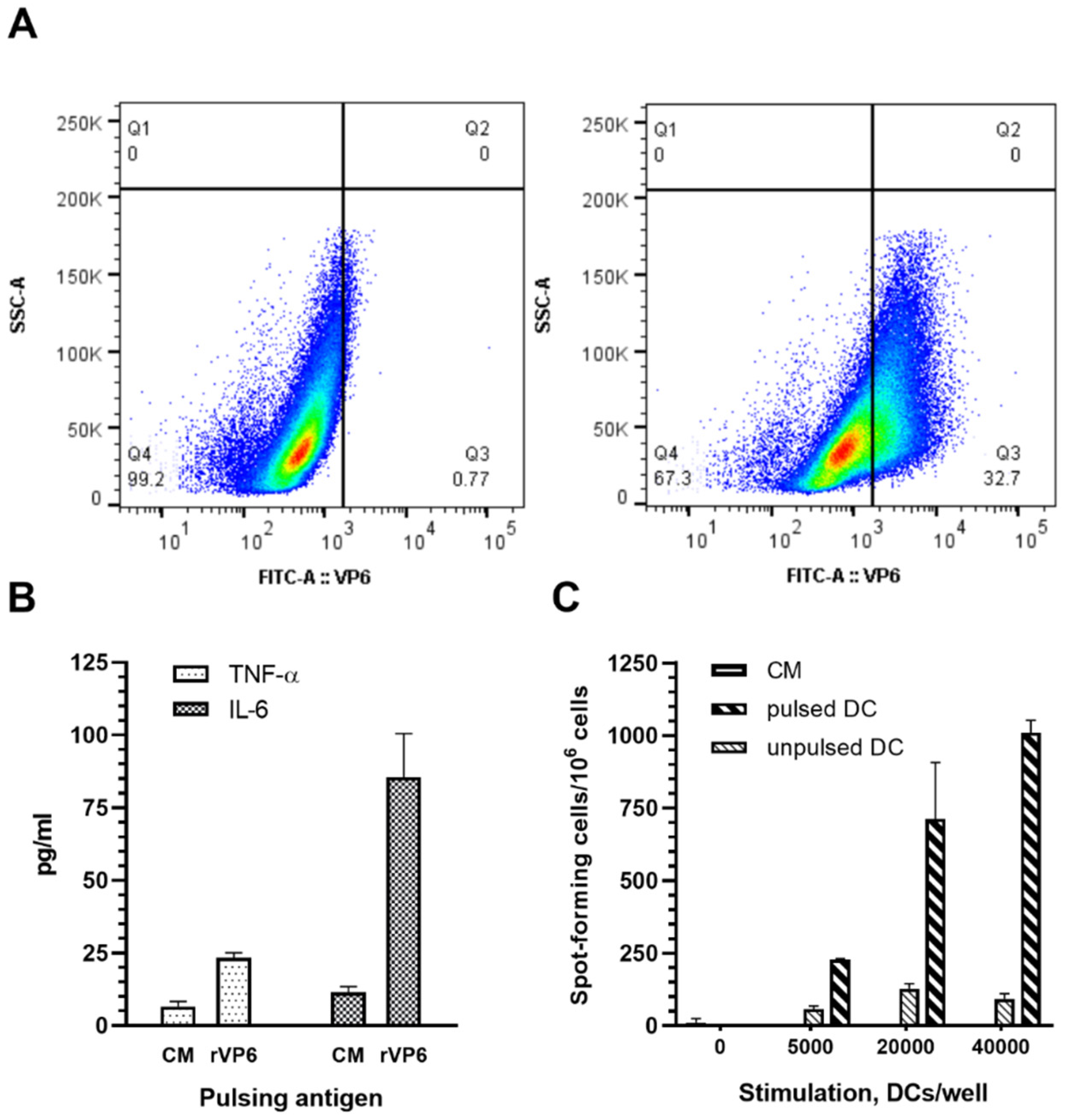
2.5. VP6 Internalization and Intracellular Staining
2.6. Cytokine Analysis
2.7. ELISPOT IFN-γ Assay
2.8. Statistics
3. Results
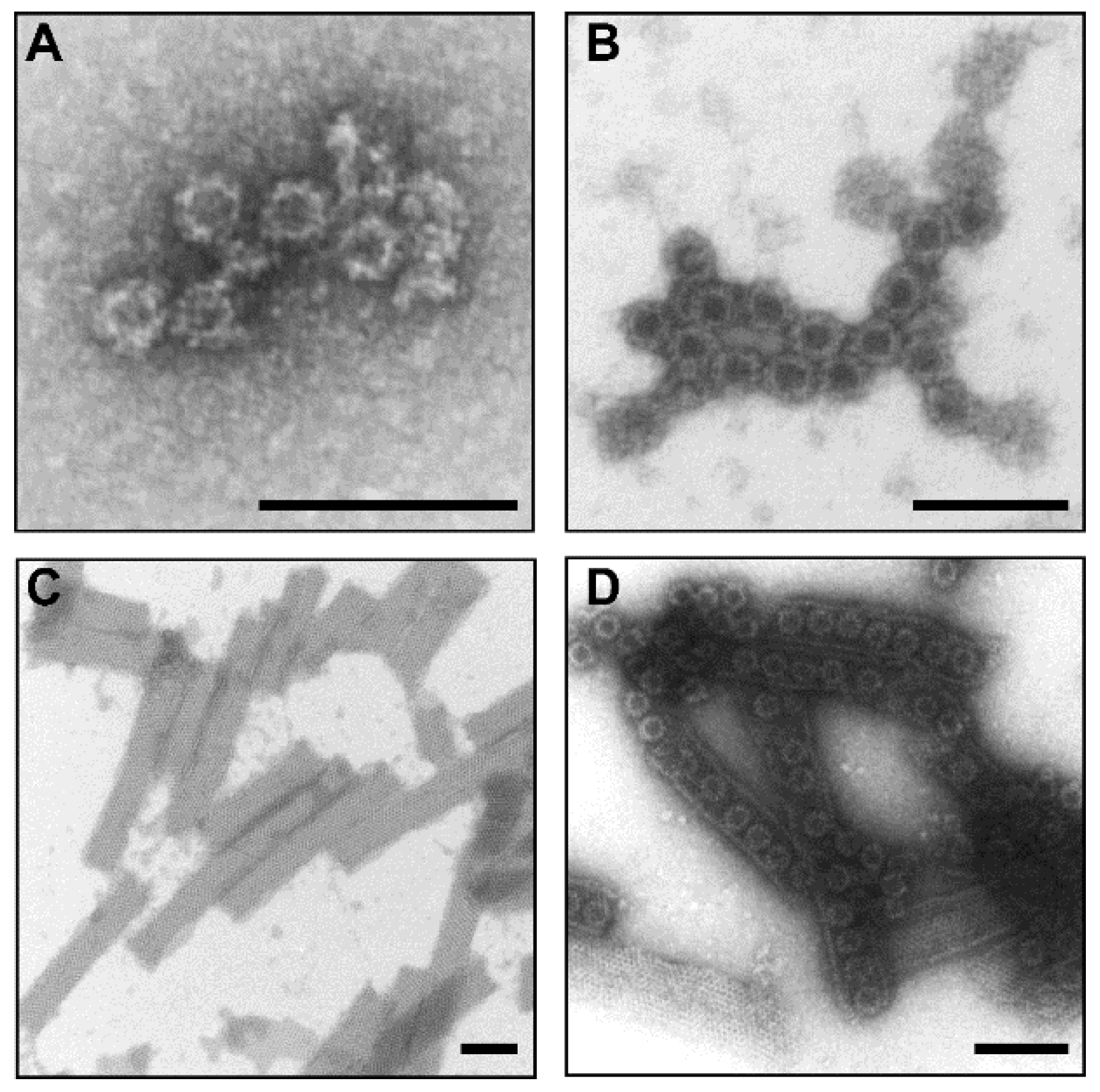
3.1. Protein Expression and Morphology
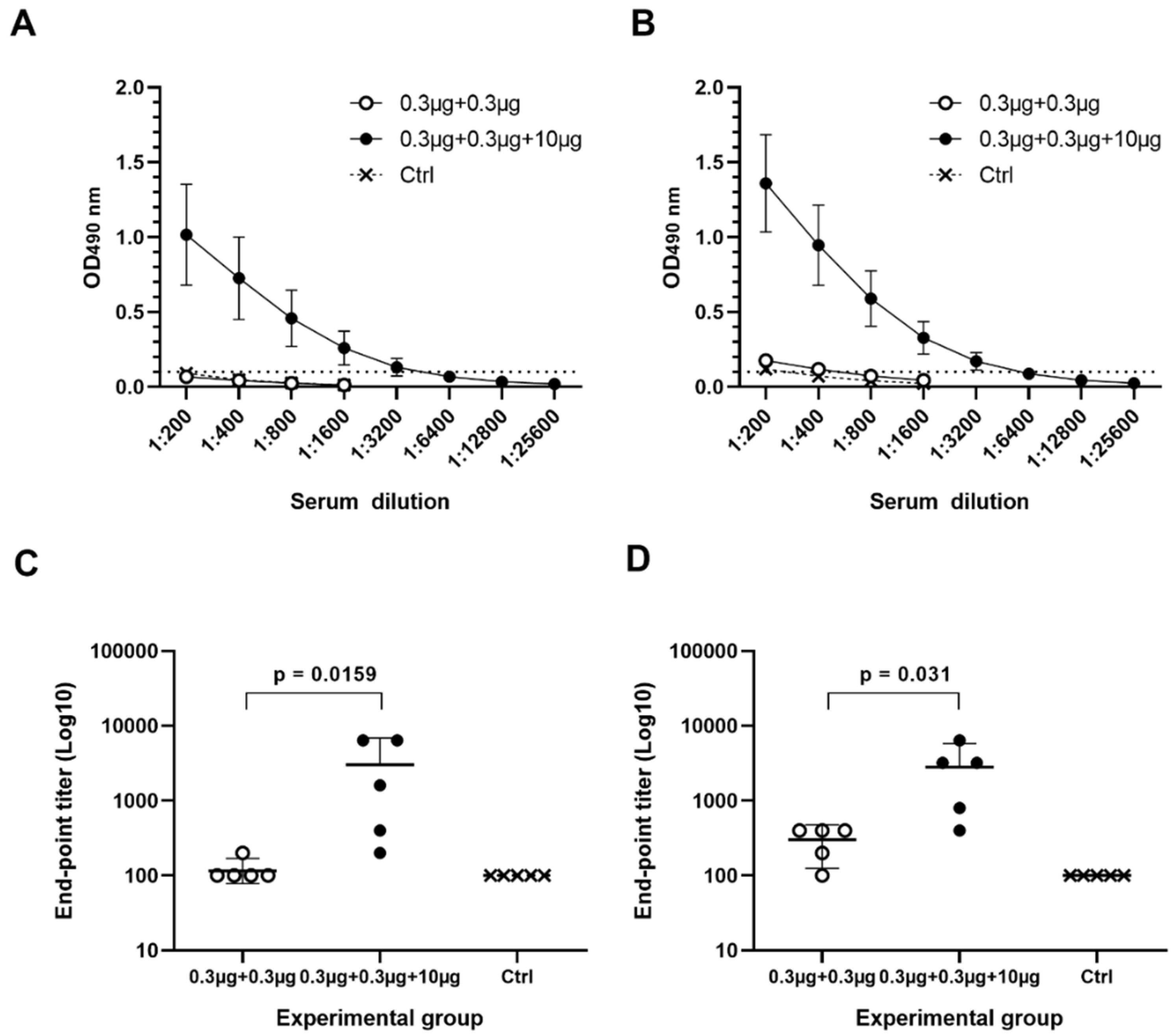
3.2. Immune Responses Induced with Suboptimal Doses of Bivalent Nov VLP Alone or Combined with RV VP6 in Excess
3.2.1. Serum NoV Genotype-Specific IgG, IgG1 and IgG2a
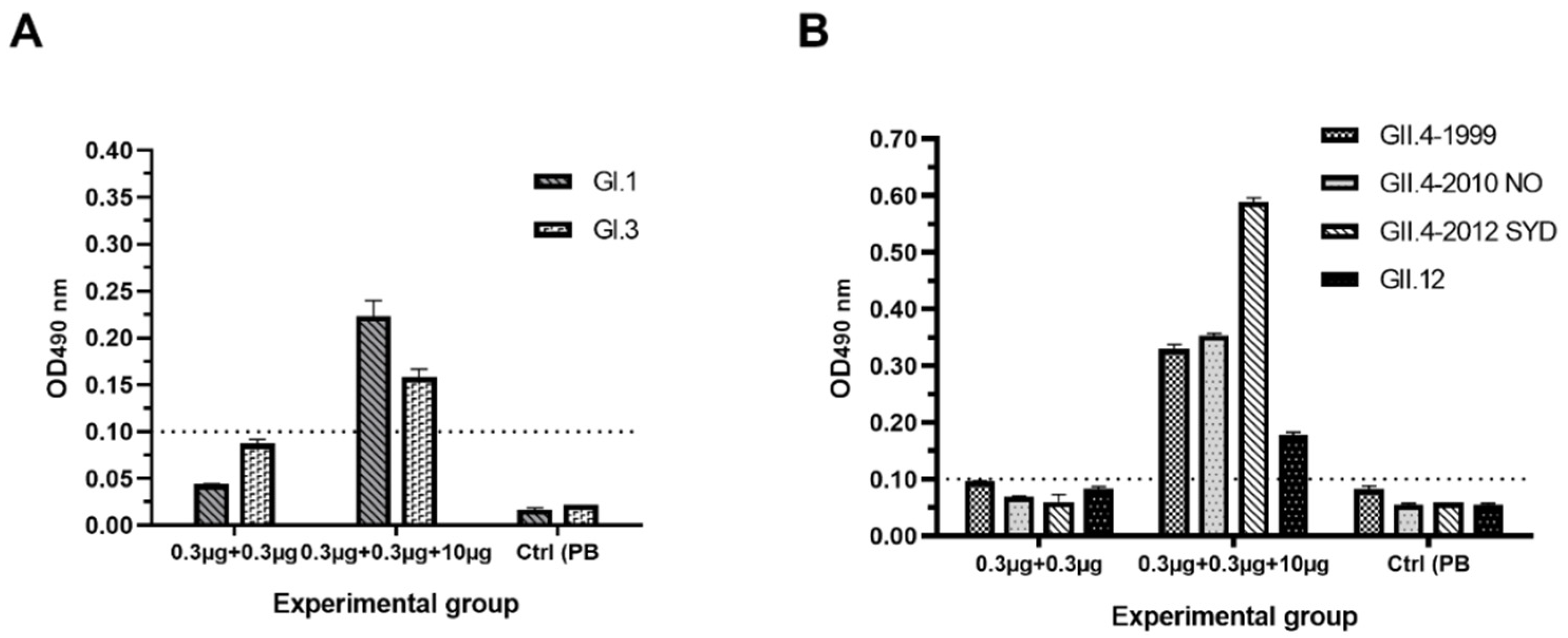
3.2.2. Serum NoV-Specific Cross-Reactive IgG Antibodies
3.2.3. Antibodies Block NoV VLP Binding to PGM HBGAs
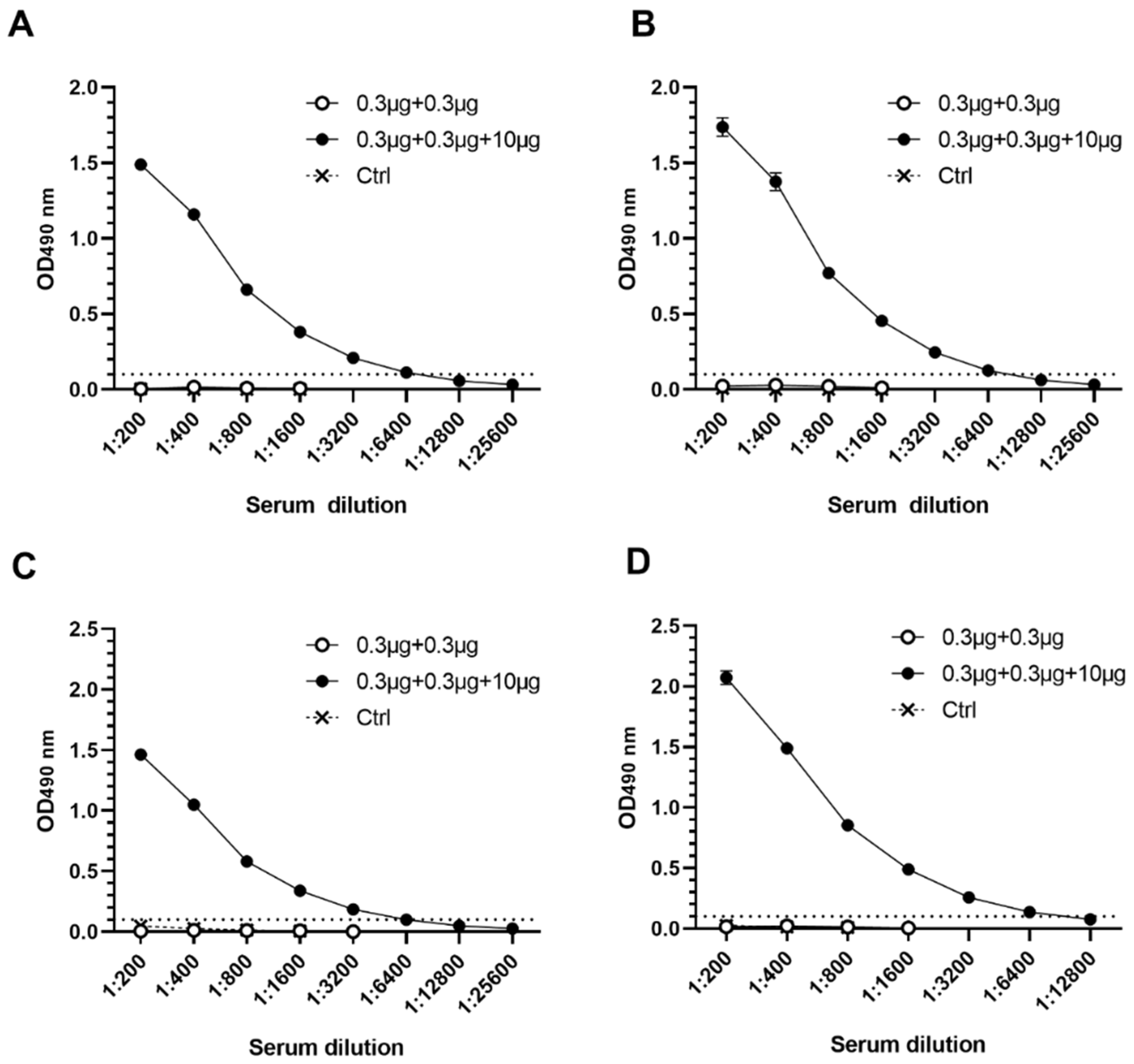
3.3. RV VP6 Adjuvant Effect on Bivalent NoV VLP Immune Responses with Equal Doses of Antigens Used
3.4. Plant-Based RV rVP6 Is Taken Up and Activates APC
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Banyai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Monedero, V.; Buesa, J.; Rodriguez-Diaz, J. The Interactions between Host Glycobiology, Bacterial Microbiota, and Viruses in the Gut. Viruses 2018, 10, 96. [Google Scholar] [CrossRef]
- Cortes-Penfield, N.W.; Ramani, S.; Estes, M.K.; Atmar, R.L. Prospects and Challenges in the Development of a Norovirus Vaccine. Clin. Ther. 2017, 39, 1537–1549. [Google Scholar] [CrossRef]
- Lucero, Y.; Vidal, R.; O’Ryan, G.M. Norovirus vaccines under development. Vaccine 2017, 36, 5435–5441. [Google Scholar] [CrossRef] [PubMed]
- GBD Diarrhoeal Diseases Collaborators. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef]
- Glass, R.I.; Parashar, U.; Patel, M.; Gentsch, J.; Jiang, B. Rotavirus vaccines: Successes and challenges. J. Infect. 2014, 68 (Suppl. 1), S9–S18. [Google Scholar] [CrossRef]
- Blazevic, V.; Lappalainen, S.; Nurminen, K.; Huhti, L.; Vesikari, T. Norovirus VLPs and rotavirus VP6 protein as combined vaccine for childhood gastroenteritis. Vaccine 2011, 29, 8126–8133. [Google Scholar] [CrossRef] [PubMed]
- Tamminen, K.; Lappalainen, S.; Huhti, L.; Vesikari, T.; Blazevic, V. Trivalent combination vaccine induces broad heterologous immune responses to norovirus and rotavirus in mice. PLoS ONE 2013, 8, e70409. [Google Scholar] [CrossRef]
- Leroux-Roels, G.; Cramer, J.P.; Mendelman, P.M.; Sherwood, J.; Clemens, R.; Aerssens, A.; De Coster, I.; Borkowski, A.; Baehner, F.; Van Damme, P. Safety and Immunogenicity of Different Formulations of Norovirus Vaccine Candidate in Healthy Adults: A Randomized, Controlled, Double-Blind Clinical Trial. J. Infect. Dis. 2018, 217, 597–607. [Google Scholar] [CrossRef]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science (New York N.Y.) 2016, 353, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Malm, M.; Tamminen, K.; Lappalainen, S.; Uusi-Kerttula, H.; Vesikari, T.; Blazevic, V. Genotype considerations for virus-like particle-based bivalent norovirus vaccine composition. Clin. Vaccine Immunol. CVI 2015, 22, 656–663. [Google Scholar] [CrossRef] [PubMed]
- LoBue, A.D.; Lindesmith, L.; Yount, B.; Harrington, P.R.; Thompson, J.M.; Johnston, R.E.; Moe, C.L.; Baric, R.S. Multivalent norovirus vaccines induce strong mucosal and systemic blocking antibodies against multiple strains. Vaccine 2006, 24, 5220–5234. [Google Scholar] [CrossRef] [PubMed]
- Kapikian, A.Z.; Chanock, Y.H.; Reoviridae, R.M. Rotaviruses. In Fields Virology, 4th ed.; Fields, B.N., Knipe, D.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2001; Chapter 55; pp. 2719–2769. [Google Scholar]
- Lappalainen, S.; Vesikari, T.; Blazevic, V. Simple and efficient ultrafiltration method for purification of rotavirus VP6 oligomeric proteins. Arch. Virol. 2016, 161, 3219–3223. [Google Scholar] [CrossRef] [PubMed]
- Lappalainen, S.; Pastor, A.R.; Malm, M.; Lopez-Guerrero, V.; Esquivel-Guadarrama, F.; Palomares, L.A.; Vesikari, T.; Blazevic, V. Protection against live rotavirus challenge in mice induced by parenteral and mucosal delivery of VP6 subunit rotavirus vaccine. Arch. Virol. 2015, 160, 2075–2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor, A.R.; Rodriguez-Limas, W.A.; Contreras, M.A.; Esquivel, E.; Esquivel-Guadarrama, F.; Ramirez, O.T.; Palomares, L.A. The assembly conformation of rotavirus VP6 determines its protective efficacy against rotavirus challenge in mice. Vaccine 2014, 32, 2874–2877. [Google Scholar] [CrossRef] [PubMed]
- Lappalainen, S.; Pastor, A.R.; Tamminen, K.; Lopez-Guerrero, V.; Esquivel-Guadarrama, F.; Palomares, L.A.; Vesikari, T.; Blazevic, V. Immune responses elicited against rotavirus middle layer protein VP6 inhibit viral replication in vitro and in vivo. Hum. Vaccines Immunother. 2014, 10, 2039–2047. [Google Scholar] [CrossRef] [Green Version]
- Choi, A.H.; McNeal, M.M.; Basu, M.; Flint, J.A.; Stone, S.C.; Clements, J.D.; Bean, J.A.; Poe, S.A.; VanCott, J.L.; Ward, R.L. Intranasal or oral immunization of inbred and outbred mice with murine or human rotavirus VP6 proteins protects against viral shedding after challenge with murine rotaviruses. Vaccine 2002, 20, 3310–3321. [Google Scholar] [CrossRef]
- Cheuvart, B.; Neuzil, K.M.; Steele, A.D.; Cunliffe, N.; Madhi, S.A.; Karkada, N.; Han, H.H.; Vinals, C. Association of serum anti-rotavirus immunoglobulin A antibody seropositivity and protection against severe rotavirus gastroenteritis: Analysis of clinical trials of human rotavirus vaccine. Hum. Vaccines Immunother. 2014, 10, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Glass, R.I.; Jiang, B.; Santosham, M.; Lopman, B.; Parashar, U. A systematic review of anti-rotavirus serum IgA antibody titer as a potential correlate of rotavirus vaccine efficacy. J. Infect. Dis. 2013, 208, 284–294. [Google Scholar] [CrossRef]
- Lappalainen, S.; Blazevic, V.; Malm, M.; Vesikari, T. Rotavirus vaccination and infection induce VP6-specific IgA responses. J. Med. Virol. 2017, 89, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Blazevic, V.; Malm, M.; Arinobu, D.; Lappalainen, S.; Vesikari, T. Rotavirus capsid VP6 protein acts as an adjuvant in vivo for norovirus virus-like particles in a combination vaccine. Hum. Vaccines Immunother. 2015, 12, 740–748. [Google Scholar] [CrossRef] [Green Version]
- Malm, M.; Heinimäki, S.; Vesikari, T.; Blazevic, V. Rotavirus capsid VP6 tubular and spherical nanostructures act as local adjuvants when co-delivered with norovirus VLPs. Clin. Exp. Immunol. 2017, 189, 331–341. [Google Scholar] [CrossRef]
- Malm, M.; Tamminen, K.; Lappalainen, S.; Vesikari, T.; Blazevic, V. Rotavirus Recombinant VP6 Nanotubes Act as an Immunomodulator and Delivery Vehicle for Norovirus Virus-Like Particles. J. Immunol. Res. 2016, 2016, 9171632. [Google Scholar] [CrossRef] [PubMed]
- Redmond, M.J.; Ohmann, H.B.; Hughes, H.P.; Sabara, M.; Frenchick, P.J.; Poku, S.K.; Ijaz, M.K.; Parker, M.D.; Laarveld, B.; Babiuk, L.A. Rotavirus particles function as immunological carriers for the delivery of peptides from infectious agents and endogenous proteins. Mol. Immunol. 1991, 28, 269–278. [Google Scholar] [CrossRef]
- Peralta, A.; Molinari, P.; Taboga, O. Chimeric recombinant rotavirus-like particles as a vehicle for the display of heterologous epitopes. Virol. J. 2009, 6, 192. [Google Scholar] [CrossRef]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Magnifection—A new platform for expressing recombinant vaccines in plants. Vaccine 2005, 23, 2042–2048. [Google Scholar] [CrossRef] [PubMed]
- Klimyuk, V.; Pogue, G.; Herz, S.; Butler, J.; Haydon, H. Production of recombinant antigens and antibodies in Nicotiana benthamiana using ‘magnifection’ technology: GMP-compliant facilities for small- and large-scale manufacturing. Curr. Top. Microbiol. Immunol. 2014, 375, 127–154. [Google Scholar] [CrossRef]
- Marillonnet, S.; Giritch, A.; Gils, M.; Kandzia, R.; Klimyuk, V.; Gleba, Y. In planta engineering of viral RNA replicons: Efficient assembly by recombination of DNA modules delivered by Agrobacterium. Proc. Natl. Acad. Sci. USA 2004, 101, 6852–6857. [Google Scholar] [CrossRef] [Green Version]
- Marillonnet, S.; Thoeringer, C.; Kandzia, R.; Klimyuk, V.; Gleba, Y. Systemic Agrobacterium tumefaciens-mediated transfection of viral replicons for efficient transient expression in plants. Nat. Biotechnol. 2005, 23, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Huhti, L.; Blazevic, V.; Nurminen, K.; Koho, T.; Hytonen, V.P.; Vesikari, T. A comparison of methods for purification and concentration of norovirus GII-4 capsid virus-like particles. Arch. Virol. 2010, 155, 1855–1858. [Google Scholar] [CrossRef] [Green Version]
- Huhti, L.; Tamminen, K.; Vesikari, T.; Blazevic, V. Characterization and immunogenicity of norovirus capsid-derived virus-like particles purified by anion exchange chromatography. Arch. Virol. 2013, 158, 933–942. [Google Scholar] [CrossRef]
- Tamminen, K.; Huhti, L.; Koho, T.; Lappalainen, S.; Hytonen, V.P.; Vesikari, T.; Blazevic, V. A comparison of immunogenicity of norovirus GII-4 virus-like particles and P-particles. Immunology 2012, 135, 89–99. [Google Scholar] [CrossRef]
- Lindesmith, L.C.; Debbink, K.; Swanstrom, J.; Vinje, J.; Costantini, V.; Baric, R.S.; Donaldson, E.F. Monoclonal antibody-based antigenic mapping of norovirus GII.4-2002. J. Virol. 2012, 86, 873–883. [Google Scholar] [CrossRef]
- Tamminen, K.; Malm, M.; Vesikari, T.; Blazevic, V. Immunological Cross-Reactivity of an Ancestral and the Most Recent Pandemic Norovirus GII.4 Variant. Viruses 2019, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Ramani, S.; Atmar, R.L.; Estes, M.K. Epidemiology of human noroviruses and updates on vaccine development. Curr. Opin. Gastroenterol. 2014, 30, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardstaff, J.L.; Clough, H.E.; Lutje, V.; McIntyre, K.M.; Harris, J.P.; Garner, P.; O’Brien, S.J. Foodborne and Food-Handler Norovirus Outbreaks: A Systematic Review. Foodborne Pathog. Dis. 2018, 15, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Reeck, A.; Kavanagh, O.; Estes, M.K.; Opekun, A.R.; Gilger, M.A.; Graham, D.Y.; Atmar, R.L. Serological correlate of protection against norovirus-induced gastroenteritis. J. Infect. Dis. 2010, 202, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Malm, M.; Uusi-Kerttula, H.; Vesikari, T.; Blazevic, V. High serum levels of norovirus genotype-specific blocking antibodies correlate with protection from infection in children. J. Infect. Dis. 2014, 210, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Blazevic, V.; Malm, M.; Salminen, M.; Oikarinen, S.; Hyoty, H.; Veijola, R.; Vesikari, T. Multiple consecutive norovirus infections in the first 2 years of life. Eur. J. Pediatr. 2015, 174, 1679–1683. [Google Scholar] [CrossRef] [Green Version]
- Malm, M.; Tamminen, K.; Vesikari, T.; Blazevic, V. Norovirus-Specific Memory T Cell Responses in Adult Human Donors. Front. Microbiol. 2016, 7, 1570. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.C.; Donaldson, E.; Leon, J.; Moe, C.L.; Frelinger, J.A.; Johnston, R.E.; Weber, D.J.; Baric, R.S. Heterotypic humoral and cellular immune responses following Norwalk virus infection. J. Virol. 2010, 84, 1800–1815. [Google Scholar] [CrossRef]
- Lindesmith, L.; Moe, C.; Lependu, J.; Frelinger, J.A.; Treanor, J.; Baric, R.S. Cellular and humoral immunity following Snow Mountain virus challenge. J. Virol. 2005, 79, 2900–2909. [Google Scholar] [CrossRef]
- Tacket, C.O.; Sztein, M.B.; Losonsky, G.A.; Wasserman, S.S.; Estes, M.K. Humoral, mucosal, and cellular immune responses to oral Norwalk virus-like particles in volunteers. Clin. Immunol. (Orlandofla.) 2003, 108, 241–247. [Google Scholar] [CrossRef]
- Malm, M.; Hyoty, H.; Knip, M.; Vesikari, T.; Blazevic, V. Development of T cell immunity to norovirus and rotavirus in children under five years of age. Sci. Rep. 2019, 9, 3199. [Google Scholar] [CrossRef]
- Heinimaki, S.; Malm, M.; Vesikari, T.; Blazevic, V. Parenterally Administered Norovirus GII.4 Virus-Like Particle Vaccine Formulated with Aluminum Hydroxide or Monophosphoryl Lipid A Adjuvants Induces Systemic but Not Mucosal Immune Responses in Mice. J. Immunol. Res. 2018, 2018, 3487095. [Google Scholar] [CrossRef]
- Jiang, X.; Shen, C.; Rey-Ladino, J.; Yu, H.; Brunham, R.C. Characterization of murine dendritic cell line JAWS II and primary bone marrow-derived dendritic cells in Chlamydia muridarum antigen presentation and induction of protective immunity. Infect. Immun. 2008, 76, 2392–2401. [Google Scholar] [CrossRef]
- Lutz, M.B.; Rossner, S. Factors influencing the generation of murine dendritic cells from bone marrow: The special role of fetal calf serum. Immunobiology 2007, 212, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Heinimaki, S.; Tamminen, K.; Malm, M.; Vesikari, T.; Blazevic, V. Live baculovirus acts as a strong B and T cell adjuvant for monomeric and oligomeric protein antigens. Virology 2017, 511, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Takahashi, H.; Hamazaki, H.; Miyano-Kurosaki, N.; Matsuura, Y.; Takaku, H. Baculovirus induces an innate immune response and confers protection from lethal influenza virus infection in mice. J. Immunol. 2003, 171, 1133–1139. [Google Scholar] [CrossRef]
- Hervas-Stubbs, S.; Rueda, P.; Lopez, L.; Leclerc, C. Insect baculoviruses strongly potentiate adaptive immune responses by inducing type I IFN. J. Immunol. 2007, 178, 2361–2369. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dose GI.4 + GII.4 | Dose VP6 | Mice/Group |
|---|---|---|---|
| I | 0.3 µg + 0.3 µg | - | 5 |
| II | 0.3 µg + 0.3 µg | 10 µg | 5 |
| III | 1 µg + 1 µg | - | 5 |
| IV | 1 µg + 1 µg | 1 µg | 5 |
| V (Control) | - 1 | - | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malm, M.; Diessner, A.; Tamminen, K.; Liebscher, M.; Vesikari, T.; Blazevic, V. Rotavirus VP6 as an Adjuvant for Bivalent Norovirus Vaccine Produced in Nicotiana benthamiana. Pharmaceutics 2019, 11, 229. https://doi.org/10.3390/pharmaceutics11050229
Malm M, Diessner A, Tamminen K, Liebscher M, Vesikari T, Blazevic V. Rotavirus VP6 as an Adjuvant for Bivalent Norovirus Vaccine Produced in Nicotiana benthamiana. Pharmaceutics. 2019; 11(5):229. https://doi.org/10.3390/pharmaceutics11050229
Chicago/Turabian StyleMalm, Maria, André Diessner, Kirsi Tamminen, Markus Liebscher, Timo Vesikari, and Vesna Blazevic. 2019. "Rotavirus VP6 as an Adjuvant for Bivalent Norovirus Vaccine Produced in Nicotiana benthamiana" Pharmaceutics 11, no. 5: 229. https://doi.org/10.3390/pharmaceutics11050229
APA StyleMalm, M., Diessner, A., Tamminen, K., Liebscher, M., Vesikari, T., & Blazevic, V. (2019). Rotavirus VP6 as an Adjuvant for Bivalent Norovirus Vaccine Produced in Nicotiana benthamiana. Pharmaceutics, 11(5), 229. https://doi.org/10.3390/pharmaceutics11050229