Skin Barriers in Dermal Drug Delivery: Which Barriers Have to Be Overcome and How Can We Measure Them?


Abstract
:1. Introduction
2. Which Skin Barriers Have to Be Overcome?
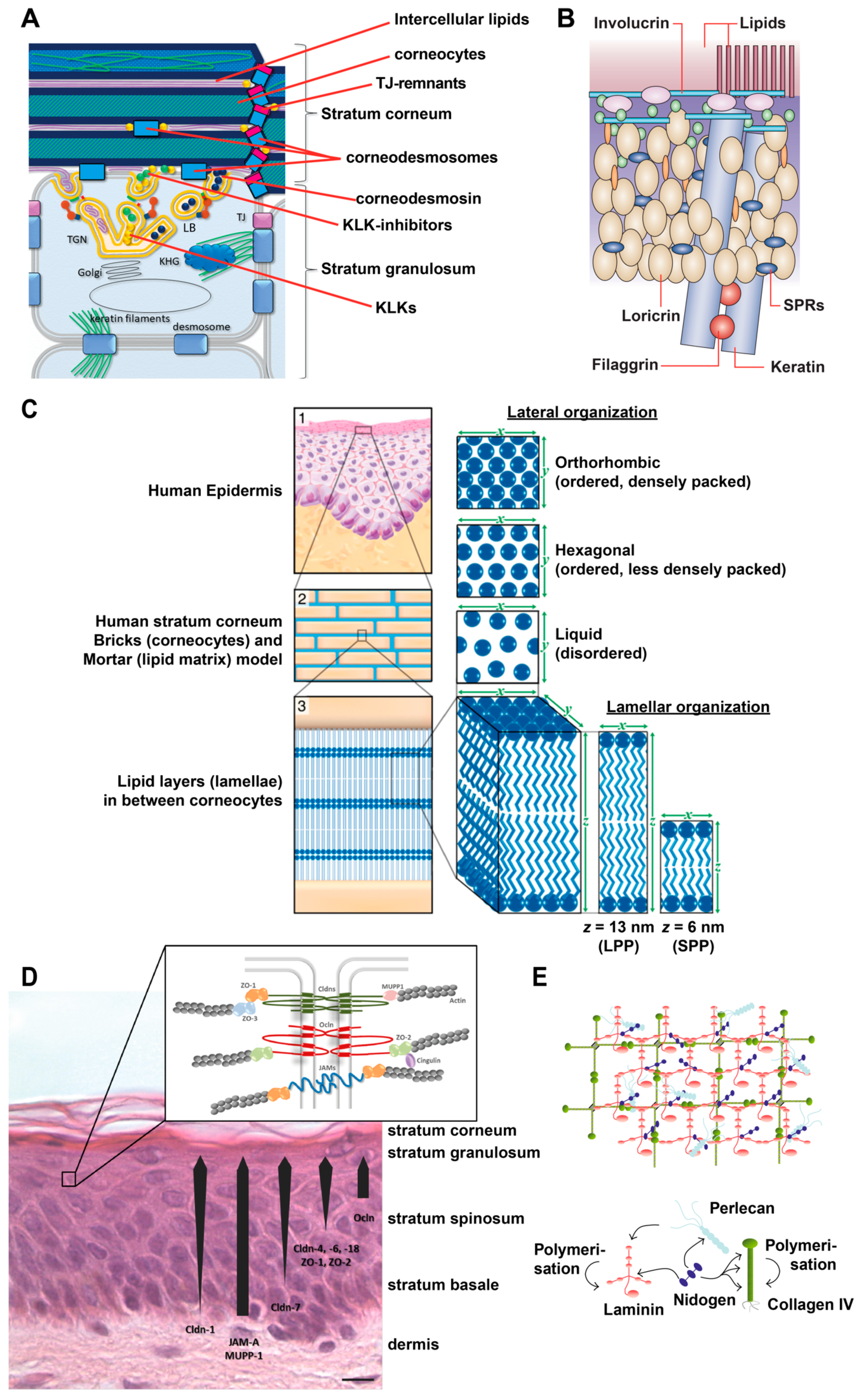
2.1. Interfollicular Epidermis
2.1.1. Stratum Corneum (SC)

2.1.2. Tight Junctions
2.1.3. Basement Membrane (Basal Lamina)
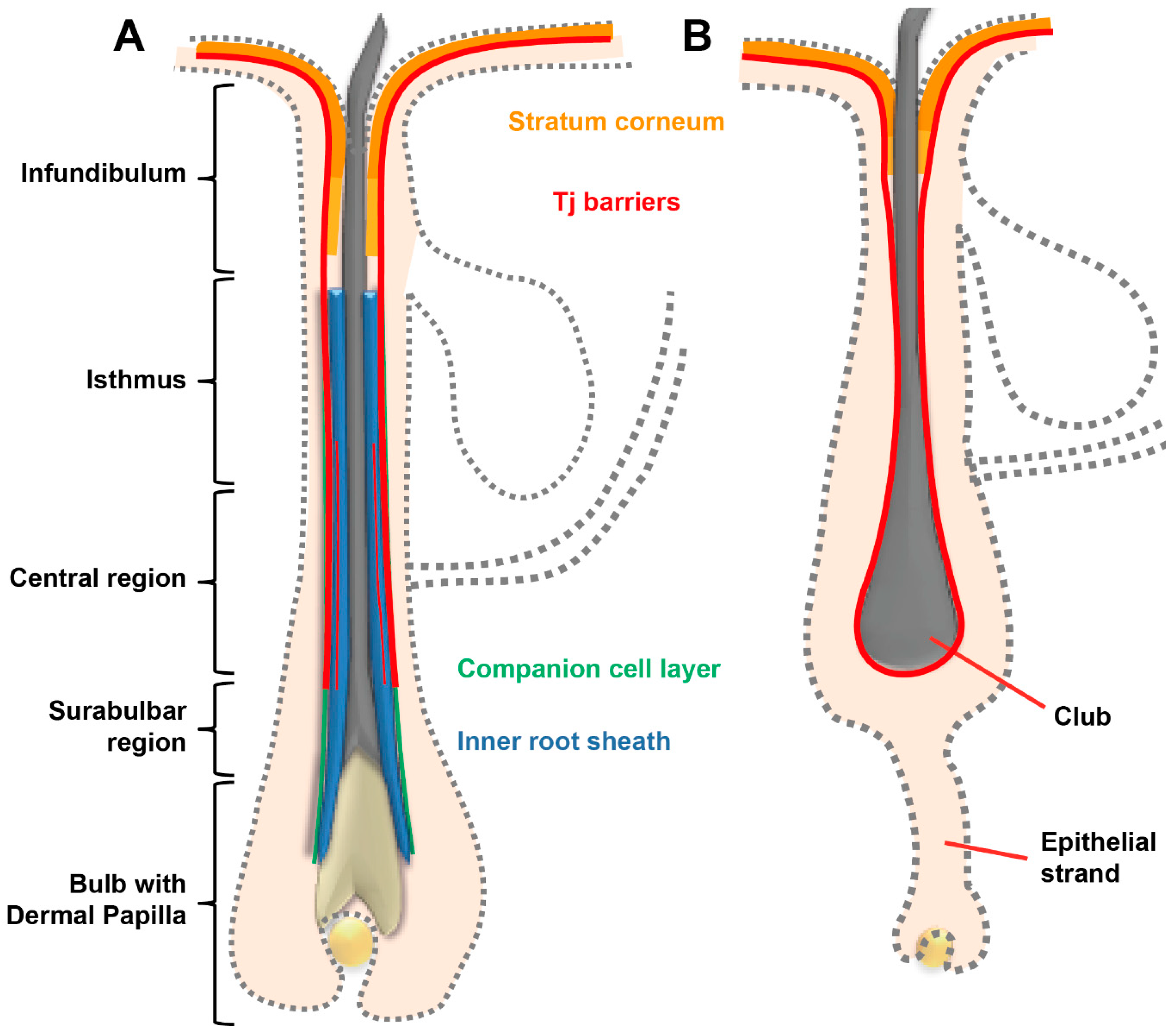
2.2. Hair Follicles
2.3. Glands
2.3.1. Sebaceous Glands
2.3.2. Sweat Glands
2.4. Blood Vessels
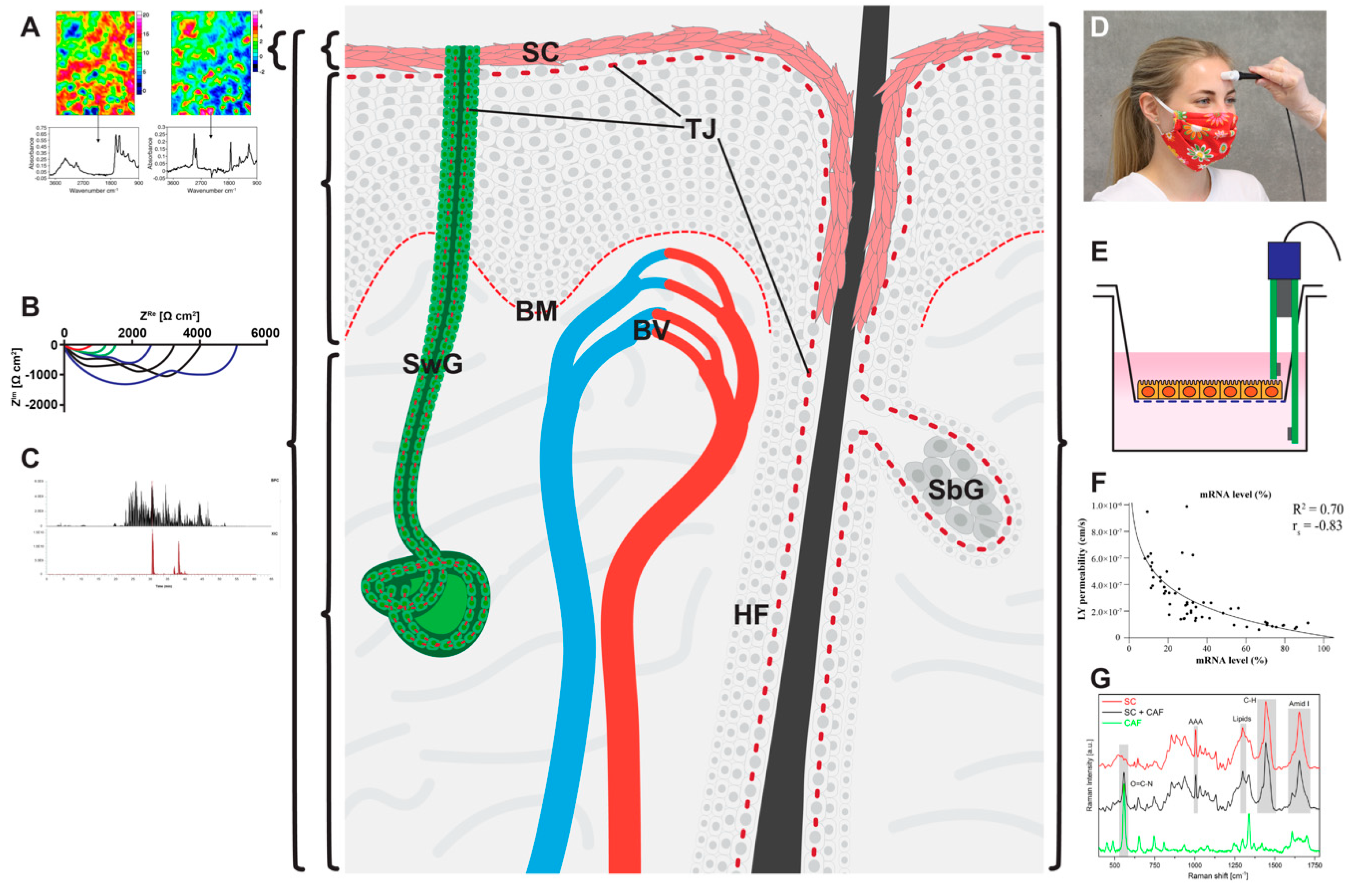
3. And How Can We Measure Them?
3.1. Physical/Chemical Methods
3.1.1. Transepidermal Water Loss (TEWL)

3.1.2. Transepithelial Electrical Resistance (TEER) and Electrical Impedance Spectroscopy (EIS)
3.1.3. Chemical and Radiochemical Analyses
3.1.4. Dye Permeation Analyses
3.1.5. Raman Spectroscopy
3.1.6. Fourier Transform Infrared (FTIR) Spectroscopy
3.2. Microscopical Methods
3.2.1. Immunohistochemical Analysis

3.2.2. Transmission Electron Microscopy
3.2.3. Optical Coherence Tomography (OCT) and Optoacoustic Imaging
3.2.4. Confocal Laser Scanning Microscopy (CLSM)
3.2.5. Multiphoton Microscopy (MPM)
4. Outlook/Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, A.Y. Molecular Mechanism of Epidermal Barrier Dysfunction as Primary Abnormalities. Int. J. Mol. Sci. 2020, 21, 1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basler, K.; Bergmann, S.; Heisig, M.; Naegel, A.; Zorn-Kruppa, M.; Brandner, J.M. The role of tight junctions in skin barrier function and dermal absorption. J. Control. Release 2016, 242, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Yokouchi, M.; Kubo, A. Maintenance of tight junction barrier integrity in cell turnover and skin diseases. Exp. Dermatol. 2018, 27, 876–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patzelt, A.; Lademann, J. Recent advances in follicular drug delivery of nanoparticles. Expert Opin. Drug Deliv. 2020, 17, 49–60. [Google Scholar] [CrossRef]
- Dragicevic, N.; Maibach, H. Percutaneous Penetration Enhancers Drug Penetration Into/Through the Skin: Methodology and General Considerations, 1st ed.; Springer: Berlin, Germany, 2017; p. 414. [Google Scholar]
- Ishida-Yamamoto, A.; Igawa, S.; Kishibe, M.; Honma, M. Clinical and molecular implications of structural changes to desmosomes and corneodesmosomes. J. Dermatol. 2018, 45, 385–389. [Google Scholar] [CrossRef]
- Matsui, T.; Amagai, M. Dissecting the formation, structure and barrier function of the stratum corneum. Int. Immunol. 2015, 27, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Jonca, N.; Leclerc, E.A.; Caubet, C.; Simon, M.; Guerrin, M.; Serre, G. Corneodesmosomes and corneodesmosin: From the stratum corneum cohesion to the pathophysiology of genodermatoses. Eur J. Dermatol. 2011, 21, 35–42. [Google Scholar] [CrossRef]
- Haftek, M. ‘Memory’ of the stratum corneum: Exploration of the epidermis’ past. Br. J. Dermatol. 2014, 171, 6–9. [Google Scholar] [CrossRef] [Green Version]
- Bouwstra, J.A.; Gooris, G.S.; van der Spek, J.A.; Bras, W. Structural investigations of human stratum corneum by small-angle X-ray scattering. J. Invest. Dermatol. 1991, 97, 1005–1012. [Google Scholar] [CrossRef] [Green Version]
- Bouwstra, J.A.; Gooris, G.S.; Bras, W.; Downing, D.T. Lipid organization in pig stratum corneum. J. Lipid Res. 1995, 36, 685–695. [Google Scholar]
- Bouwstra, J.A.; Thewalt, J.; Gooris, G.S.; Kitson, N. A model membrane approach to the epidermal permeability barrier: An X-ray diffraction study. Biochemistry 1997, 36, 7717–7725. [Google Scholar] [CrossRef] [PubMed]
- Boncheva, M.; Damien, F.; Normand, V. Molecular organization of the lipid matrix in intact Stratum corneum using ATR-FTIR spectroscopy. Biochim. Biophys. Acta (BBA) Biomembr. 2008, 1778, 1344–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojumdar, E.H.; Kariman, Z.; van Kerckhove, L.; Gooris, G.S.; Bouwstra, J.A. The role of ceramide chain length distribution on the barrier properties of the skin lipid membranes. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 2473–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Smeden, J.; Bouwstra, J.A. Stratum Corneum Lipids: Their Role for the Skin Barrier Function in Healthy Subjects and Atopic Dermatitis Patients. In Skin Barrier Function; Agner, T., Ed.; Karger: Basel, Switherland, 2016; Volume 49, pp. 8–26. [Google Scholar]
- Wertz, P.W. Lipids and the Permeability and Antimicrobial Barriers of the Skin. J. Lipids 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, A.; Guy, R. Infrared spectroscopic and differential scanning calorimetric investigations of the stratum corneum barrier function. Drugs Pharm. Sci. 1997, 83, 87–162. [Google Scholar]
- Ludovici, M.; Kozul, N.; Materazzi, S.; Risoluti, R.; Picardo, M.; Camera, E. Influence of the sebaceous gland density on the stratum corneum lipidome. Sci. Rep. 2018, 8, 11500. [Google Scholar] [CrossRef]
- Ishida-Yamamoto, A.; Igawa, S.; Kishibe, M. Molecular basis of the skin barrier structures revealed by electron microscopy. Exp. Dermatol. 2018, 27, 841–846. [Google Scholar] [CrossRef] [Green Version]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef]
- Janssens, M.; van Smeden, J.; Gooris, G.S.; Bras, W.; Portale, G.; Caspers, P.J.; Vreeken, R.J.; Hankemeier, T.; Kezic, S.; Wolterbeek, R.; et al. Increase in short-chain ceramides correlates with an altered lipid organization and decreased barrier function in atopic eczema patients. J. Lipid Res. 2012, 53, 2755–2766. [Google Scholar] [CrossRef] [Green Version]
- Breitkreutz, D.; Koxholt, I.; Thiemann, K.; Nischt, R. Skin basement membrane: The foundation of epidermal integrity--BM functions and diverse roles of bridging molecules nidogen and perlecan. Biomed. Res. Int. 2013, 2013, 179784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monash, S.; Blank, H. Location and reformation of the epithelial barrier to water vapor. AMA Arch. Derm. 1958, 78, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Rancan, F.; Giulbudagian, M.; Jurisch, J.; Blume-Peytavi, U.; Calderon, M.; Vogt, A. Drug delivery across intact and disrupted skin barrier: Identification of cell populations interacting with penetrated thermoresponsive nanogels. Eur. J. Pharm. Biopharm. 2017, 116, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Parhi, R.; Suresh, P.; Patnaik, S. Physical means of stratum corneum barrier manipulation to enhance transdermal drug delivery. Curr. Drug Deliv. 2015, 12, 122–138. [Google Scholar] [CrossRef] [PubMed]
- Sklar, L.R.; Burnett, C.T.; Waibel, J.S.; Moy, R.L.; Ozog, D.M. Laser assisted drug delivery: A review of an evolving technology. Lasers Surg. Med. 2014, 46, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Asfour, M.H. Advanced trends in protein and peptide drug delivery: A special emphasis on aquasomes and microneedles techniques. Drug Deliv. Transl. Res. 2020. [Google Scholar] [CrossRef]
- Del Rio-Sancho, S.; Lapteva, M.; Sonaje, K.; Bohler, C.; Ling, V.; Boehncke, W.H.; Kalia, Y.N. Targeted cutaneous delivery of etanercept using Er:YAG fractional laser ablation. Int. J. Pharm. 2020, 580, 119234. [Google Scholar] [CrossRef]
- Kezic, S.; Jakasa, I. Filaggrin and Skin Barrier Function. Curr. Probl. Dermatol. 2016, 49, 1–7. [Google Scholar] [CrossRef] [PubMed]
- McLean, W.H. Filaggrin failure-from ichthyosis vulgaris to atopic eczema and beyond. Br. J. Dermatol. 2016, 175, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, H.; Nagao, K.; Kubo, A.; Hata, T.; Shimizu, A.; Mizuno, H.; Yamada, T.; Amagai, M. Altered stratum corneum barrier and enhanced percutaneous immune responses in filaggrin-null mice. J. Allergy Clin. Immunol. 2012, 129, 1538–1546. [Google Scholar] [CrossRef] [Green Version]
- Thyssen, J.P.; Jakasa, I.; Riethmuller, C.; Schon, M.P.; Braun, A.; Haftek, M.; Fallon, P.G.; Wroblewski, J.; Jakubowski, H.; Eckhart, L.; et al. Filaggrin Expression and Processing Deficiencies Impair Corneocyte Surface Texture and Stiffness in Mice. J. Investig. Dermatol. 2020, 140, 615–623. [Google Scholar] [CrossRef]
- Niehues, H.; Schalkwijk, J.; van Vlijmen-Willems, I.; Rodijk-Olthuis, D.; van Rossum, M.M.; Wladykowski, E.; Brandner, J.M.; van den Bogaard, E.H.J.; Zeeuwen, P. Epidermal equivalents of filaggrin null keratinocytes do not show impaired skin barrier function. J. Allergy Clin. Immunol. 2017, 139, 1979–1981 e1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kezic, S. Loss-of-function mutations in filaggrin gene and malignant melanoma. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proksch, E. pH in nature, humans and skin. J. Dermatol 2018, 45, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kaur, T.; Malhotra, S.K.; Rai, J.; Chaudhari, S. Correlation of Vitamin D3 Levels and SCORAD Index in Atopic Dermatits: A Case Control Study. J. Clin. Diagn. Res. 2017, 11, WC01–WC03. [Google Scholar] [CrossRef] [PubMed]
- Marenholz, I.; Rivera, V.A.; Esparza-Gordillo, J.; Bauerfeind, A.; Lee-Kirsch, M.A.; Ciechanowicz, A.; Kurek, M.; Piskackova, T.; Macek, M.; Lee, Y.A. Association screening in the Epidermal Differentiation Complex (EDC) identifies an SPRR3 repeat number variant as a risk factor for eczema. J. Investig. Dermatol. 2011, 131, 1644–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catunda, R.; Rekhi, U.; Clark, D.; Levin, L.; Febbraio, M. Loricrin downregulation and epithelial-related disorders: A systematic review. J. Dtsch. Dermatol. Ges. 2019, 17, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Moosbrugger-Martinz, V.; Jalili, A.; Schossig, A.S.; Jahn-Bassler, K.; Zschocke, J.; Schmuth, M.; Stingl, G.; Eckl, K.M.; Hennies, H.C.; Gruber, R. Epidermal barrier abnormalities in exfoliative ichthyosis with a novel homozygous loss-of-function mutation in CSTA. Br. J. Dermatol. 2015, 172, 1628–1632. [Google Scholar] [CrossRef]
- Muttardi, K.; Nitoiu, D.; Kelsell, D.P.; O’Toole, E.A.; Batta, K. Acral peeling skin syndrome associated with a novel CSTA gene mutation. Clin. Exp. Dermatol. 2016, 41, 394–398. [Google Scholar] [CrossRef]
- Rice, R.H.; Durbin-Johnson, B.P.; Ishitsuka, Y.; Salemi, M.; Phinney, B.S.; Rocke, D.M.; Roop, D.R. Proteomic Analysis of Loricrin Knockout Mouse Epidermis. J. Proteome Res. 2016, 15, 2560–2566. [Google Scholar] [CrossRef]
- Boczonadi, V.; Maatta, A. Functional Analysis of Periplakin and Envoplakin, Cytoskeletal Linkers, and Cornified Envelope Precursor Proteins. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 569, pp. 309–329. [Google Scholar] [CrossRef]
- Cangkrama, M.; Darido, C.; Georgy, S.R.; Partridge, D.; Auden, A.; Srivastava, S.; Wilanowski, T.; Jane, S.M. Two Ancient Gene Families Are Critical for Maintenance of the Mammalian Skin Barrier in Postnatal Life. J. Investig. Dermatol. 2016, 136, 1438–1448. [Google Scholar] [CrossRef]
- van der Velden, J.J.; van Geel, M.; Nellen, R.G.; Jonkman, M.F.; McGrath, J.A.; Nanda, A.; Sprecher, E.; van Steensel, M.A.; McLean, W.H.; Cassidy, A.J. Novel TGM5 mutations in acral peeling skin syndrome. Exp. Dermatol. 2015, 24, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Karim, N.; Phinney, B.S.; Salemi, M.; Wu, P.W.; Naeem, M.; Rice, R.H. Human stratum corneum proteomics reveals cross-linking of a broad spectrum of proteins in cornified envelopes. Exp. Dermatol. 2019, 28, 618–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, M.; Nomura, T.; Sugiyama, T.; Miyauchi, T.; Suzuki, S.; Fujita, Y.; Shimizu, H. Compound heterozygous missense mutations p.Leu207Pro and p.Tyr544Cys in TGM1 cause a severe form of lamellar ichthyosis. J. Dermatol. 2018, 45, 1463–1467. [Google Scholar] [CrossRef]
- Has, C. Peeling Skin Disorders: A Paradigm for Skin Desquamation. J. Investig. Dermatol. 2018, 138, 1689–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclerc, E.A.; Huchenq, A.; Mattiuzzo, N.R.; Metzger, D.; Chambon, P.; Ghyselinck, N.B.; Serre, G.; Jonca, N.; Guerrin, M. Corneodesmosin gene ablation induces lethal skin-barrier disruption and hair-follicle degeneration related to desmosome dysfunction. J. Cell Sci. 2009, 122, 2699–2709. [Google Scholar] [CrossRef] [Green Version]
- Grubauer, G.; Elias, P.M.; Feingold, K.R. Transepidermal water loss: The signal for recovery of barrier structure and function. J. Lipid Res. 1989, 30, 323–333. [Google Scholar]
- Sajic, D.; Asiniwasis, R.; Skotnicki-Grant, S. A look at epidermal barrier function in atopic dermatitis: Physiologic lipid replacement and the role of ceramides. Skin Ther. Lett. 2012, 17, 6–9. [Google Scholar]
- Kihara, A. Synthesis and degradation pathways, functions, and pathology of ceramides and epidermal acylceramides. Prog. Lipid Res. 2016, 63, 50–69. [Google Scholar] [CrossRef]
- Bouwstra, J.A.; Honeywell-Nguyen, P.L.; Gooris, G.S.; Ponec, M. Structure of the skin barrier and its modulation by vesicular formulations. Prog. Lipid Res. 2003, 42, 1–36. [Google Scholar] [CrossRef]
- Williams, A.C.; Barry, B.W. Penetration enhancers. Adv. Drug Deliv. Rev. 2004, 56, 603–618. [Google Scholar] [CrossRef]
- Dragicevic, N.; Atkinson, J.P.; Maibach, H.I. Chemical penetration enhancers: Classification and mode of action. In Percutaneous Penetration Enhancers Chemical Methods in Penetration Enhancement; Springer: Berlin/Heidelberg, Germany, 2015; pp. 11–27. [Google Scholar]
- Hadgraft, J. Passive enhancement strategies in topical and transdermal drug delivery. Int. J. Pharm. 1999, 184, 1–6. [Google Scholar] [CrossRef]
- Shiohara, T.; Sato, Y.; Komatsu, Y.; Ushigome, Y.; Mizukawa, Y. Sweat as an Efficient Natural Moisturizer. Curr. Probl. Dermatol. 2016, 51, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Gorzelanny, C.; Goerge, T.; Schnaeker, E.M.; Thomas, K.; Luger, T.A.; Schneider, S.W. Atomic force microscopy as an innovative tool for nanoanalysis of native stratum corneum. Exp. Dermatol. 2006, 15, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Wohlrab, J.; Gebert, A.; Neubert, R.H.H. Lipids in the Skin and pH. Curr. Probl. Dermatol. 2018, 54, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Piontek, J.; Krug, S.M.; Protze, J.; Krause, G.; Fromm, M. Molecular architecture and assembly of the tight junction backbone. Biochim. Biophys. Acta (BBA) Biomembr. 2020, 1862, 183279. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, N.; Rosenthal, R.; Furuse, M.; Moll, I.; Fromm, M.; Brandner, J.M. Contribution of Tight Junction Proteins to Ion, Macromolecule, and Water Barrier in Keratinocytes. J. Investig. Dermatol. 2013, 133, 1161–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.P.; Mallet, D.G.; Pettet, G.J. Towards a quantitative theory of epidermal calcium profile formation in unwounded skin. PLoS ONE 2015, 10, e0116751. [Google Scholar] [CrossRef] [Green Version]
- Gunzel, D.; Yu, A.S. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [Green Version]
- Brandner, J.M.; Zorn-Kruppa, M.; Yoshida, T.; Moll, I.; Beck, L.A.; De Benedetto, A. Epidermal tight junctions in health and disease. Tissue Barriers 2015, 3, e974451. [Google Scholar] [CrossRef] [Green Version]
- Furuse, M.; Hata, M.; Furuse, K.; Yoshida, Y.; Haratake, A.; Sugitani, Y.; Noda, T.; Kubo, A.; Tsukita, S. Claudin-based tight junctions are crucial for the mammalian epidermal barrier: A lesson from claudin-1-deficient mice. J. Cell Biol. 2002, 156, 1099–1111. [Google Scholar] [CrossRef]
- Yokouchi, M.; Kubo, A.; Kawasaki, H.; Yoshida, K.; Ishii, K.; Furuse, M.; Amagai, M. Epidermal tight junction barrier function is altered by skin inflammation, but not by filaggrin-deficient stratum corneum. J. Dermatol. Sci. 2015, 77, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, T.; Iwamoto, N.; Akashi, M.; Kojima, T.; Hisatsune, J.; Sugai, M.; Furuse, M. Tight junction dysfunction in the stratum granulosum leads to aberrant stratum corneum barrier function in claudin-1-deficient mice. J. Dermatol. Sci. 2013, 70, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Yuki, T.; Komiya, A.; Kusaka, A.; Kuze, T.; Sugiyama, Y.; Inoue, S. Impaired tight junctions obstruct stratum corneum formation by altering polar lipid and profilaggrin processing. J. Dermatol. Sci. 2013, 69, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Hadj-Rabia, S.; Baala, L.; Vabres, P.; Hamel-Teillac, D.; Jacquemin, E.; Fabre, M.; Lyonnet, S.; De Prost, Y.; Munnich, A.; Hadchouel, M.; et al. Claudin-1 gene mutations in neonatal sclerosing cholangitis associated with ichthyosis: A tight junction disease. Gastroenterology 2004, 127, 1386–1390. [Google Scholar] [CrossRef]
- Feldmeyer, L.; Huber, M.; Fellmann, F.; Beckmann, J.S.; Frenk, E.; Hohl, D. Confirmation of the origin of NISCH syndrome. Hum. Mutat. 2006, 27, 408–410. [Google Scholar] [CrossRef]
- Nagtzaam, I.F.; van Geel, M.; Driessen, A.; Steijlen, P.M.; van Steensel, M.A. Bile duct paucity is part of the neonatal ichthyosis-sclerosing cholangitis phenotype. Br. J. Dermatol. 2010, 163, 205–207. [Google Scholar] [CrossRef]
- Paganelli, M.; Stephenne, X.; Gilis, A.; Jacquemin, E.; Henrion Caude, A.; Girard, M.; Gonzales, E.; Revencu, N.; Reding, R.; Wanty, C.; et al. Neonatal ichthyosis and sclerosing cholangitis syndrome: Extremely variable liver disease severity from claudin-1 deficiency. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 350–354. [Google Scholar] [CrossRef]
- Kirchmeier, P.; Sayar, E.; Hotz, A.; Hausser, I.; Islek, A.; Yilmaz, A.; Artan, R.; Fischer, J. Novel mutation in the CLDN1 gene in a Turkish family with neonatal ichthyosis sclerosing cholangitis (NISCH) syndrome. Br. J. Dermatol. 2014, 170, 976–978. [Google Scholar] [CrossRef]
- Batista, D.I.; Perez, L.; Orfali, R.L.; Zaniboni, M.C.; Samorano, L.P.; Pereira, N.V.; Sotto, M.N.; Ishizaki, A.S.; Oliveira, L.M.; Sato, M.N.; et al. Profile of skin barrier proteins (filaggrin, claudins 1 and 4) and Th1/Th2/Th17 cytokines in adults with atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 1091–1095. [Google Scholar] [CrossRef]
- Bergmann, S.; von Buenau, B.; Vidal, Y.S.S.; Haftek, M.; Wladykowski, E.; Houdek, P.; Lezius, S.; Duplan, H.; Basler, K.; Dahnhardt-Pfeiffer, S.; et al. Claudin-1 decrease impacts epidermal barrier function in atopic dermatitis lesions dose-dependently. Sci. Rep. 2020, 10, 2024. [Google Scholar] [CrossRef]
- Gruber, R.; Bornchen, C.; Rose, K.; Daubmann, A.; Volksdorf, T.; Wladykowski, E.; Vidal, Y.S.S.; Peters, E.M.; Danso, M.; Bouwstra, J.A.; et al. Diverse regulation of claudin-1 and claudin-4 in atopic dermatitis. Am. J. Pathol. 2015, 185, 2777–2789. [Google Scholar] [CrossRef]
- De Benedetto, A.; Slifka, M.K.; Rafaels, N.M.; Kuo, I.H.; Georas, S.N.; Boguniewicz, M.; Hata, T.; Schneider, L.C.; Hanifin, J.M.; Gallo, R.L.; et al. Reductions in claudin-1 may enhance susceptibility to herpes simplex virus 1 infections in atopic dermatitis. J. Allergy Clin. Immunol. 2011, 128, 242–246 e245. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Kubo, A.; Fujita, H.; Yokouchi, M.; Ishii, K.; Kawasaki, H.; Nomura, T.; Shimizu, H.; Kouyama, K.; Ebihara, T.; et al. Distinct behavior of human Langerhans cells and inflammatory dendritic epidermal cells at tight junctions in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2014, 134, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Tokumasu, R.; Yamaga, K.; Yamazaki, Y.; Murota, H.; Suzuki, K.; Tamura, A.; Bando, K.; Furuta, Y.; Katayama, I.; Tsukita, S. Dose-dependent role of claudin-1 in vivo in orchestrating features of atopic dermatitis. Proc. Natl. Acad. Sci. USA 2016, 113, E4061–E4068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, M.G.; Anderson, E.A.; Pandya, R.P.; De Benedetto, A.; Yoshida, T.; Hilimire, T.A.; Martinez-Sobrido, L.; Beck, L.A.; Miller, B.L. Peptides Derived from the Tight Junction Protein CLDN1 Disrupt the Skin Barrier and Promote Responsiveness to an Epicutaneous Vaccine. J. Investig. Dermatol. 2020, 140, 361–369 e363. [Google Scholar] [CrossRef] [PubMed]
- Beier, L.S.; Rossa, J.; Woodhouse, S.; Bergmann, S.; Kramer, H.B.; Protze, J.; Eichner, M.; Piontek, A.; Vidal, Y.S.S.; Brandner, J.M.; et al. Use of Modified Clostridium perfringens Enterotoxin Fragments for Claudin Targeting in Liver and Skin Cells. Int. J. Mol. Sci. 2019, 20, 4774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, M.; Nagase, S.; Iida, M.; Takeda, S.; Yamashita, M.; Watari, A.; Shirasago, Y.; Fukasawa, M.; Takeda, H.; Sawasaki, T.; et al. Claudin-1 Binder Enhances Epidermal Permeability in a Human Keratinocyte Model. J. Pharmacol. Exp. Ther. 2015, 354, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Uchida, T.; Kanazawa, T.; Takashima, Y.; Okada, H. Development of an efficient transdermal delivery system of small interfering RNA using functional peptides, Tat and AT-1002. Chem. Pharm. Bull. 2011, 59, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Ibaraki, H.; Kanazawa, T.; Kurano, T.; Oogi, C.; Takashima, Y.; Seta, Y. Anti-RelA siRNA-Encapsulated Flexible Liposome with Tight Junction-Opening Peptide as a Non-invasive Topical Therapeutic for Atopic Dermatitis. Biol. Pharm. Bull. 2019, 42, 1216–1225. [Google Scholar] [CrossRef] [Green Version]
- Abdayem, R.; Callejon, S.; Portes, P.; Kirilov, P.; Demarne, F.; Pirot, F.; Jannin, V.; Haftek, M. Modulation of transepithelial electric resistance (TEER) in reconstructed human epidermis by excipients known to permeate intestinal tight junctions. Exp. Dermatol. 2015, 24, 686–691. [Google Scholar] [CrossRef]
- Nystrom, A.; Bruckner-Tuderman, L. Matrix molecules and skin biology. In Seminars in Cell and Developmental Biology; Elesvier: Amsterdam, The Netherlands, 2019; Volume 89, pp. 136–146. [Google Scholar] [CrossRef]
- Witte, M.; Zillikens, D.; Schmidt, E. Diagnosis of Autoimmune Blistering Diseases. Front. Med. 2018, 5, 296. [Google Scholar] [CrossRef] [PubMed]
- Stemmler, S.; Parwez, Q.; Petrasch-Parwez, E.; Epplen, J.T.; Hoffjan, S. Association of variation in the LAMA3 gene, encoding the alpha-chain of laminin 5, with atopic dermatitis in a German case-control cohort. BMC Dermatol. 2014, 14, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.I.; Park, H.J.; Won, Y.Y.; Choi, H.; Jeong, K.H.; Sung, J.Y.; Shin, M.K. Basement Membrane Status Is Intact in Urticarial Dermatitis vs. Adult-Onset Atopic Dermatitis. Ann. Dermatol. 2018, 30, 258–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiner, E.; Wolff, K. The permeability of the intercellular space of the epidermis for low molecular weight protein. Electron microscopic cytochemical studies with peroxidase as a tracer substance. Archiv fur Klinische und Experimentelle Dermatologie 1969, 235, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Wolff, K.; Honigsmann, H. Permeability of the epidermis and the phagocytic activity of keratinocytes. Ultrastructural studies with thorotrast as a marker. J. Ultrastruct. Res. 1971, 36, 176–190. [Google Scholar] [CrossRef]
- Briggaman, R.A.; Wheeler, C.E., Jr. The epidermal-dermal junction. J. Investig. Dermatol. 1975, 65, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Weeks, B.S.; Ramchandran, R.S.; Hopkins, J.J.; Friedman, H.M. Herpes simplex virus type-1 and -2 pathogenesis is restricted by the epidermal basement membrane. Arch. Virol. 2000, 145, 385–396. [Google Scholar] [CrossRef]
- Kazama, T.; Yaoita, E.; Ito, M.; Sato, Y. Charge-selective permeability of dermo-epidermal junction: Tracer studies with cationic and anionic ferritins. J. Investig. Dermatol. 1988, 91, 560–565. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.W.; Kloepper, J.; Langan, E.A.; Kim, Y.; Yeo, J.; Kim, M.J.; Hsi, T.C.; Rose, C.; Yoon, G.S.; Lee, S.J.; et al. A Guide to Studying Human Hair Follicle Cycling In Vivo. J. Investig. Dermatol. 2016, 136, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Lademann, J.; Knorr, F.; Richter, H.; Blume-Peytavi, U.; Vogt, A.; Antoniou, C.; Sterry, W.; Patzelt, A. Hair follicles--an efficient storage and penetration pathway for topically applied substances. Skin Pharmacol. Physiol. 2008, 21, 150–155. [Google Scholar] [CrossRef]
- Vogt, A.; Blume-Peytavi, U. Selective hair therapy: Bringing science to the fiction. Exp. Dermatol. 2014, 23, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Zorn-Kruppa, M.; Vidal, Y.S.S.; Houdek, P.; Wladykowski, E.; Grzybowski, S.; Gruber, R.; Gorzelanny, C.; Harcup, J.; Schneider, S.W.; Majumdar, A.; et al. Tight Junction barriers in human hair follicles-role of claudin-1. Sci. Rep. 2018, 8, 12800. [Google Scholar] [CrossRef] [Green Version]
- Blume-Peytavi, U.; Vogt, A. Human hair follicle: Reservoir function and selective targeting. Br. J. Dermatol. 2011, 165, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Itoh, M.; Saitou, M.; Ando-Akatsuka, Y.; Furuse, M.; Yoneda, K.; Imamura, S.; Fujimoto, K.; Tsukita, S. Subcellular distribution of tight junction-associated proteins (occludin, ZO-1, ZO-2) in rodent skin. J. Investig. Dermatol. 1998, 110, 862–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebbe, B.; Mankertz, J.; Schwarz, C.; Amasheh, S.; Fromm, M.; Assaf, C.; Schultz-Ehrenburg, U.; Sanchez Ruderish, H.; Schulzke, J.D.; Orfanos, C.E. Tight junction proteins: A novel class of integral membrane proteins. Expression in human epidermis and in HaCaT keratinocytes. Arch. Dermatol. Res. 2002, 294, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Brandner, J.M.; McIntyre, M.; Kief, S.; Wladykowski, E.; Moll, I. Expression and localization of tight junction-associated proteins in human hair follicles. Arch. Dermatol. Res. 2003, 295, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Raymond, K.; Richter, A.; Kreft, M.; Frijns, E.; Janssen, H.; Slijper, M.; Praetzel-Wunder, S.; Langbein, L.; Sonnenberg, A. Expression of the orphan protein Plet-1 during trichilemmal differentiation of anagen hair follicles. J. Investig. Dermatol. 2010, 130, 1500–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsugi, T.; Yokouchi, M.; Hirano, T.; Hirabayashi, A.; Nagai, T.; Ohyama, M.; Abe, T.; Kaneko, M.; Zouboulis, C.C.; Amagai, M.; et al. Holocrine Secretion Occurs outside the Tight Junction Barrier in Multicellular Glands: Lessons from Claudin-1-Deficient Mice. J. Investig. Dermatol. 2020, 140, 298–308 e295. [Google Scholar] [CrossRef]
- Hashimoto, K. Demonostration of the intercellular spaces of the human eccrine sweat gland by lanthanum. I. The secretory coil. J. Ultrastruct. Res. 1971, 36, 249–262. [Google Scholar] [CrossRef]
- Hashimoto, K. Demonstration of the intercellular spaces of the human eccrine sweat gland by lanthanum. II. The duct. J. Ultrastruct. Res. 1971, 37, 504–520. [Google Scholar] [CrossRef]
- Briggman, J.V.; Bank, H.L.; Bigelow, J.B.; Graves, J.S.; Spicer, S.S. Structure of the tight junctions of the human eccrine sweat gland. Am. J. Anat. 1981, 162, 357–368. [Google Scholar] [CrossRef]
- Wilke, K.; Wepf, R.; Keil, F.J.; Wittern, K.P.; Wenck, H.; Biel, S.S. Are sweat glands an alternate penetration pathway? Understanding the morphological complexity of the axillary sweat gland apparatus. Skin Pharmacol. Physiol. 2006, 19, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Klar, J.; Piontek, J.; Milatz, S.; Tariq, M.; Jameel, M.; Breiderhoff, T.; Schuster, J.; Fatima, A.; Asif, M.; Sher, M.; et al. Altered paracellular cation permeability due to a rare CLDN10B variant causes anhidrosis and kidney damage. PLoS Genet. 2017, 13, e1006897. [Google Scholar] [CrossRef] [PubMed]
- Yamaga, K.; Murota, H.; Tamura, A.; Miyata, H.; Ohmi, M.; Kikuta, J.; Ishii, M.; Tsukita, S.; Katayama, I. Claudin-3 Loss Causes Leakage of Sweat from the Sweat Gland to Contribute to the Pathogenesis of Atopic Dermatitis. J. Investig. Dermatol. 2018, 138, 1279–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jajack, A.; Brothers, M.; Kasting, G.; Heikenfeld, J. Enhancing glucose flux into sweat by increasing paracellular permeability of the sweat gland. PLoS ONE 2018, 13, e0200009. [Google Scholar] [CrossRef]
- Petrofsky, J.S. Control of Skin Blood Flow. In Textbook of Aging Skin; Farage, M.A., Miller, K.W., Maibach, H.I., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1091–1104. [Google Scholar] [CrossRef]
- Kaiyala, K.J. Energy Homeostasis: Thermoregulation. In Encyclopedia of Neuroscience; Squire, L.R., Ed.; Elsevier: Oxford, UK, 2009; pp. 1043–1052. [Google Scholar] [CrossRef]
- Hao, J.; Ghosh, P.; Li, S.K.; Newman, B.; Kasting, G.B.; Raney, S.G. Heat effects on drug delivery across human skin. Expert Opin. Drug Deliv. 2016, 13, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Nanjwade, B.K. Novel Drug Delivery Systems and Regulatory Affairs; CBS PUBL & DIST PVT Limited I: Delhi, India, 2019. [Google Scholar]
- Shomaker, T.S.; Zhang, J.; Ashburn, M.A. Assessing the impact of heat on the systemic delivery of fentanyl through the transdermal fentanyl delivery system. Pain Med. 2000, 1, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, A.; Sasaki, M.; Harada, K.; Kumagai, Y.; Ohashi, K.; Ebihara, A. Influences of bathing and hot weather on the pharmacokinetics of a new transdermal clonidine, M-5041T. J. Clin. Pharmacol. 1996, 36, 892–896. [Google Scholar] [CrossRef]
- Shomaker, T.S.; Zhang, J.; Ashburn, M.A. A pilot study assessing the impact of heat on the transdermal delivery of testosterone. J. Clin. Pharmacol. 2001, 41, 677–682. [Google Scholar] [CrossRef]
- Petersen, K.K.; Rousing, M.L.; Jensen, C.; Arendt-Nielsen, L.; Gazerani, P. Effect of local controlled heat on transdermal delivery of nicotine. Int. J. Physiol. Pathophysiol. Pharmacol. 2011, 3, 236–242. [Google Scholar]
- Alexander, H.; Brown, S.; Danby, S.; Flohr, C. Research Techniques Made Simple: Transepidermal Water Loss Measurement as a Research Tool. J. Investig. Dermatol. 2018, 138, 2295–2300 e2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jungersted, J.M.; Scheer, H.; Mempel, M.; Baurecht, H.; Cifuentes, L.; Hogh, J.K.; Hellgren, L.I.; Jemec, G.B.; Agner, T.; Weidinger, S. Stratum corneum lipids, skin barrier function and filaggrin mutations in patients with atopic eczema. Allergy 2010, 65, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Kopecna, M.; Machacek, M.; Novackova, A.; Paraskevopoulos, G.; Roh, J.; Vavrova, K. Esters of terpene alcohols as highly potent, reversible, and low toxic skin penetration enhancers. Sci. Rep. 2019, 9, 14617. [Google Scholar] [CrossRef] [Green Version]
- Kazarian, S.G.; Chan, K.L. Applications of ATR-FTIR spectroscopic imaging to biomedical samples. Biochim. Biophys. Acta (BBA) Biomembr. 2006, 1758, 858–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiani, P.; Schlüter, H. Mass Spectrometric Investigation and Determination of Proteome Composition of Human Skin Tissues Ablated Using Picosecond Infrared Laser (PIRL) in a “layer by Layer” Approach. Ph.D. Thesis, University of Hamburg, Hamburg, Germany, 2019. [Google Scholar]
- Franzen, L.; Anderski, J.; Windbergs, M. Quantitative detection of caffeine in human skin by confocal Raman spectroscopy-A systematic in vitro validation study. Eur. J. Pharm. Biopharm. 2015, 95, 110–116. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, A.; Rafaels, N.M.; McGirt, L.Y.; Ivanov, A.I.; Georas, S.N.; Cheadle, C.; Berger, A.E.; Zhang, K.; Vidyasagar, S.; Yoshida, T.; et al. Tight junction defects in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2011, 127, 773–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celli, A.; Zhai, Y.; Jiang, Y.J.; Crumrine, D.; Elias, P.M.; Feingold, K.R.; Mauro, T.M. Tight junction properties change during epidermis development. Exp. Dermatol. 2012, 21, 798–801. [Google Scholar] [CrossRef] [Green Version]
- Schimek, K.; Hsu, H.H.; Boehme, M.; Kornet, J.J.; Marx, U.; Lauster, R.; Portner, R.; Lindner, G. Bioengineering of a Full-Thickness Skin Equivalent in a 96-Well Insert Format for Substance Permeation Studies and Organ-On-A-Chip Applications. Bioengineering 2018, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Kuo, I.H.; Carpenter-Mendini, A.; Yoshida, T.; McGirt, L.Y.; Ivanov, A.I.; Barnes, K.C.; Gallo, R.L.; Borkowski, A.W.; Yamasaki, K.; Leung, D.Y.; et al. Activation of epidermal toll-like receptor 2 enhances tight junction function: Implications for atopic dermatitis and skin barrier repair. J. Investig. Dermatol. 2013, 133, 988–998. [Google Scholar] [CrossRef] [Green Version]
- Giaever, I.; Keese, C.R. Micromotion of mammalian cells measured electrically. Proc. Natl. Acad. Sci. USA 1991, 88, 7896–7900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, M.; Pereira, S.; Pohl, L.; Ketelhut, S.; Kemper, B.; Gorzelanny, C.; Galla, H.J.; Moerschbacher, B.M.; Goycoolea, F.M. Chitosan encapsulation modulates the effect of capsaicin on the tight junctions of MDCK cells. Sci. Rep. 2015, 5, 10048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, S.M.; Amasheh, S.; Richter, J.F.; Milatz, S.; Gunzel, D.; Westphal, J.K.; Huber, O.; Schulzke, J.D.; Fromm, M. Tricellulin forms a barrier to macromolecules in tricellular tight junctions without affecting ion permeability. Mol. Biol. Cell 2009, 20, 3713–3724. [Google Scholar] [CrossRef] [Green Version]
- Rinaldi, A.O.; Morita, H.; Wawrzyniak, P.; Dreher, A.; Grant, S.; Svedenhag, P.; Akdis, C.A. Direct assessment of skin epithelial barrier by electrical impedance spectroscopy. Allergy 2019, 74, 1934–1944. [Google Scholar] [CrossRef] [PubMed]
- Gerasimenko, T.; Nikulin, S.; Zakharova, G.; Poloznikov, A.; Petrov, V.; Baranova, A.; Tonevitsky, A. Impedance Spectroscopy as a Tool for Monitoring Performance in 3D Models of Epithelial Tissues. Front. Bioeng. Biotechnol. 2019, 7, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannweiler, R.; Bergmann, S.; Vidal-y-Sy, S.; Brandner, J.M.; Günzel, D. Direct assessment of individual skin barrier components by electrical impedance spectroscopy. Biosens. Bioelectron. 2020. submitted. [Google Scholar]
- Kopecna, M.; Machacek, M.; Prchalova, E.; Stepanek, P.; Drasar, P.; Kotora, M.; Vavrova, K. Dodecyl Amino Glucoside Enhances Transdermal and Topical Drug Delivery via Reversible Interaction with Skin Barrier Lipids. Pharm. Res. 2017, 34, 640–653. [Google Scholar] [CrossRef]
- White, E.A.; Orazem, M.E.; Bunge, A.L. Characterization of damaged skin by impedance spectroscopy: Chemical damage by dimethyl sulfoxide. Pharm. Res. 2013, 30, 2607–2624. [Google Scholar] [CrossRef] [PubMed]
- Kandekar, S.G.; Singhal, M.; Sonaje, K.B.; Kalia, Y.N. Polymeric micelle nanocarriers for targeted epidermal delivery of the hedgehog pathway inhibitor vismodegib: Formulation development and cutaneous biodistribution in human skin. Expert Opin. Drug Deliv. 2019, 16, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Herbig, M.E.; Houdek, P.; Gorissen, S.; Zorn-Kruppa, M.; Wladykowski, E.; Volksdorf, T.; Grzybowski, S.; Kolios, G.; Willers, C.; Mallwitz, H.; et al. A custom tailored model to investigate skin penetration in porcine skin and its comparison with human skin. Eur. J. Pharm. Biopharm. 2015, 95, 99–109. [Google Scholar] [CrossRef]
- Rancan, F.; Contardi, M.; Jurisch, J.; Blume-Peytavi, U.; Vogt, A.; Bayer, I.S.; Schaudinn, C. Evaluation of Drug Delivery and Efficacy of Ciprofloxacin-Loaded Povidone Foils and Nanofiber Mats in a Wound-Infection Model Based on Ex Vivo Human Skin. Pharmaceutics 2019, 11, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausmann, C.; Hertz-Kleptow, D.; Zoschke, C.; Wanjiku, B.; Wentzien-Odenthal, A.; Kerscher, M.; Schafer-Korting, M. Reconstructed Human Epidermis Predicts Barrier-Improving Effects of Lactococcus lactis Emulsion in Humans. Skin Pharmacol. Physiol. 2019, 32, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Zoschke, C.; Schilrreff, P.; Romero, E.L.; Brandner, J.M.; Schafer-Korting, M. Dendritic nanoparticles for cutaneous drug delivery--testing in human skin and reconstructed human skin. Curr. Pharm. Des. 2015, 21, 2784–2800. [Google Scholar] [CrossRef] [PubMed]
- Beissner, N.; Bolea Albero, A.; Fuller, J.; Kellner, T.; Lauterboeck, L.; Liang, J.; Bol, M.; Glasmacher, B.; Muller-Goymann, C.C.; Reichl, S. Improved in vitro models for preclinical drug and formulation screening focusing on 2D and 3D skin and cornea constructs. Eur. J. Pharm. Biopharm. 2018, 126, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Bartosova, L.; Bajgar, J. Transdermal drug delivery in vitro using diffusion cells. Curr. Med. Chem. 2012, 19, 4671–4677. [Google Scholar] [CrossRef]
- Wagner, H.; Kostka, K.H.; Lehr, C.M.; Schaefer, U.F. Interrelation of permeation and penetration parameters obtained from in vitro experiments with human skin and skin equivalents. J. Control. Release 2001, 75, 283–295. [Google Scholar] [CrossRef]
- Herkenne, C.; Naik, A.; Kalia, Y.N.; Hadgraft, J.; Guy, R.H. Effect of propylene glycol on ibuprofen absorption into human skin in vivo. J. Pharm. Sci. 2008, 97, 185–197. [Google Scholar] [CrossRef]
- Rancan, F.; Volkmann, H.; Giulbudagian, M.; Schumacher, F.; Stanko, J.I.; Kleuser, B.; Blume-Peytavi, U.; Calderon, M.; Vogt, A. Dermal Delivery of the High-Molecular-Weight Drug Tacrolimus by Means of Polyglycerol-Based Nanogels. Pharmaceutics 2019, 11, 394. [Google Scholar] [CrossRef] [Green Version]
- Esim, O.; Savaser, A.; Karadurmus, L.; Bakirhan, N.K.; Ozkan, S.A.; Ozkan, Y. Current Status at Drug Delivery Approaches and Assay of Anti-Migraine Drugs. Curr. Drug Deliv. 2020. [Google Scholar] [CrossRef]
- Carrer, V.; Alonso, C.; Oliver, M.A.; Coderch, L. In vitro penetration through the skin layers of topically applied glucocorticoids. Drug Test. Anal. 2018, 10, 1528–1535. [Google Scholar] [CrossRef]
- Lau, W.M.; Ng, K.W. Finite and infinite dosing. In Percutaneous Penetration Enhancers Drug Penetration Into/Through the Skin; Springer: Berlin/Heidelberg, Germany, 2017; pp. 35–44. [Google Scholar]
- Kurasawa, M.; Kuroda, S.; Kida, N.; Murata, M.; Oba, A.; Yamamoto, T.; Sasaki, H. Regulation of tight junction permeability by sodium caprate in human keratinocytes and reconstructed epidermis. Biochem. Biophys. Res. Commun. 2009, 381, 171–175. [Google Scholar] [CrossRef]
- Auner, B.G.; Petzenhauser, E.; Valenta, C. Influence of 6-ketocholestanol on skin permeation of 5-aminolevulinic acid and evaluation of chemical stability. J. Pharm. Sci. 2004, 93, 2780–2787. [Google Scholar] [CrossRef] [PubMed]
- Volz, P.; Schilrreff, P.; Brodwolf, R.; Wolff, C.; Stellmacher, J.; Balke, J.; Morilla, M.J.; Zoschke, C.; Schafer-Korting, M.; Alexiev, U. Pitfalls in using fluorescence tagging of nanomaterials: Tecto-dendrimers in skin tissue as investigated by Cluster-FLIM. Ann. N. Y. Acad. Sci. 2017, 1405, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Falcone, D.; Uzunbajakava, N.E.; Varghese, B.; de Aquino Santos, G.R.; Richters, R.J.; van de Kerkhof, P.C.; van Erp, P.E. Microspectroscopic Confocal Raman and Macroscopic Biophysical Measurements in the in vivo Assessment of the Skin Barrier: Perspective for Dermatology and Cosmetic Sciences. Skin Pharmacol. Physiol. 2015, 28, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Franzen, L.; Windbergs, M. Applications of Raman spectroscopy in skin research--From skin physiology and diagnosis up to risk assessment and dermal drug delivery. Adv. Drug. Deliv. Rev. 2015, 89, 91–104. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Q.; Mao, G.; Conroy, O.; Pyatski, Y.; Fevola, M.J.; Cula, G.O.; Maitra, P.; Mendelsohn, R.; Flach, C.R. Novel confocal Raman microscopy method to investigate hydration mechanisms in human skin. Skin Res. Technol. 2019, 25, 653–661. [Google Scholar] [CrossRef]
- Choe, C.; Schleusener, J.; Choe, S.; Lademann, J.; Darvin, M.E. A modification for the calculation of water depth profiles in oil-treated skin by in vivo confocal Raman microscopy. J. Biophotonics 2020, 13, e201960106. [Google Scholar] [CrossRef]
- Mateus, R.; Abdalghafor, H.; Oliveira, G.; Hadgraft, J.; Lane, M.E. A new paradigm in dermatopharmacokinetics-Confocal Raman spectroscopy. Int. J. Pharm. 2013, 444, 106–108. [Google Scholar] [CrossRef]
- Bakonyi, M.; Gacsi, A.; Kovacs, A.; Szucs, M.B.; Berko, S.; Csanyi, E. Following-up skin penetration of lidocaine from different vehicles by Raman spectroscopic mapping. J. Pharm. Biomed. Anal. 2018, 154, 1–6. [Google Scholar] [CrossRef]
- Yew, Y.W.; Dinish, U.S.; Choi, E.C.E.; Bi, R.; Ho, C.J.H.; Dev, K.; Li, X.; Attia, A.B.E.; Wong, M.K.W.; Balasundaram, G.; et al. Investigation of morphological, vascular and biochemical changes in the skin of an atopic dermatitis (AD) patient in response to dupilumab using raster scanning optoacoustic mesoscopy (RSOM) and handheld confocal Raman spectroscopy (CRS). J. Dermatol. Sci. 2019, 95, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Choe, C.; Ri, J.; Schleusener, J.; Lademann, J.; Darvin, M.E. The non-homogenous distribution and aggregation of carotenoids in the stratum corneum correlates with the organization of intercellular lipids in vivo. Exp. Dermatol. 2019, 28, 1237–1243. [Google Scholar] [CrossRef]
- Darvin, M.E.; Choe, C.S.; Schleusener, J.; Lademann, J. Non-invasive depth profiling of the stratum corneum in vivo using confocal Raman microscopy considering the non-homogeneous distribution of keratin. Biomed. Opt. Express 2019, 10, 3092–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohan, S.B.; Saeidpour, S.; Solik, A.; Schanzer, S.; Richter, H.; Dong, P.; Darvin, M.E.; Bodmeier, R.; Patzelt, A.; Zoubari, G.; et al. Investigation of the cutaneous penetration behavior of dexamethasone loaded to nano-sized lipid particles by EPR spectroscopy, and confocal Raman and laser scanning microscopy. Eur. J. Pharm. Biopharm. 2017, 116, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Konig, K.; Breunig, H.G.; Batista, A.; Schindele, A.; Zieger, M.; Kaatz, M. Translation of two-photon microscopy to the clinic: Multimodal multiphoton CARS tomography of in vivo human skin. J. Biomed. Opt. 2020, 25, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, P.; Batista, A.; Zieger, M.; Kaatz, M.; Koenig, K. Artificial Intelligence in Multiphoton Tomography: Atopic Dermatitis Diagnosis. Sci. Rep. 2020, 10, 7968. [Google Scholar] [CrossRef]
- Griffiths, P.R.; De Haseth, J.A. Fourier Transform Infrared Spectrometry; John Wiley & Sons: New York, NY, USA, 2007; Volume 171. [Google Scholar]
- Perkins, W.D. Fourier-Transform Infrared-Spectroscopy 2. Advantages of Ft-Ir. J. Chem. Educ. 1987, 64, A269–A271. [Google Scholar] [CrossRef]
- Suss, B.; Ringleb, F.; Heberle, J. New ultrarapid-scanning interferometer for FT-IR spectroscopy with microsecond time-resolution. Rev. Sci. Instrum. 2016, 87, 063113. [Google Scholar] [CrossRef] [PubMed]
- Covi-Schwarz, J.; Klang, V.; Valenta, C. ATR-FTIR Spectroscopy and the Skin Barrier: Evaluation of Penetration-Enhancement Effects. In Percutaneous Penetration Enhancers Drug Penetration Into/Through the Skin; Springer: Berlin/Heidelberg, Germany, 2017; pp. 247–254. [Google Scholar]
- Kirschner, N.; Houdek, P.; Fromm, M.; Moll, I.; Brandner, J.M. Tight junctions form a barrier in human epidermis. Eur. J. Cell Biol. 2010, 89, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Yokouchi, M.; Nagao, K.; Ishii, K.; Amagai, M.; Kubo, A. Functional tight junction barrier localizes in the second layer of the stratum granulosum of human epidermis. J. Dermatol. Sci. 2013, 71, 89–99. [Google Scholar] [CrossRef]
- Mathes, C.; Brandner, J.M.; Laue, M.; Raesch, S.S.; Hansen, S.; Failla, A.V.; Vidal, S.; Moll, I.; Schaefer, U.F.; Lehr, C.M. Tight junctions form a barrier in porcine hair follicles. Eur. J. Cell Biol. 2016, 95, 89–99. [Google Scholar] [CrossRef]
- Gruber, R.; Elias, P.M.; Crumrine, D.; Lin, T.K.; Brandner, J.M.; Hachem, J.P.; Presland, R.B.; Fleckman, P.; Janecke, A.R.; Sandilands, A.; et al. Filaggrin genotype in ichthyosis vulgaris predicts abnormalities in epidermal structure and function. Am. J. Pathol. 2011, 178, 2252–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaytsev, S.M.; Svenskaya, Y.I.; Lengert, E.V.; Terentyuk, G.S.; Bashkatov, A.N.; Tuchin, V.V.; Genina, E.A. Optimized skin optical clearing for optical coherence tomography monitoring of encapsulated drug delivery through the hair follicles. J. Biophotonics 2020, 13, e201960020. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. Intercellular spaces of the human epidermis as demonstrated with lanthanum. J. Investig. Dermatol. 1971, 57, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Baroli, B.; Ennas, M.G.; Loffredo, F.; Isola, M.; Pinna, R.; Lopez-Quintela, M.A. Penetration of metallic nanoparticles in human full-thickness skin. J. Investig. Dermatol. 2007, 127, 1701–1712. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, P.F.; Peng, S.; Tang, T.C.; Lin, S.Y.; Tsai, H.C. Enhancing the in vivo transdermal delivery of gold nanoparticles using poly(ethylene glycol) and its oleylamine conjugate. Int. J. Nanomed. 2016, 11, 1867–1878. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Engelhardt, R.; Rosperich-Palm, J.; Huettmann, G.; Birngruber, R. Measurement of optical-transport coefficients of Intralipid in visible and NIR range. In Laser-Tissue Interaction V; and Ultraviolet Radiation Hazards; International Society for Optics and Photonics: Bellingham, WA, USA, 1994; Volume 2134, pp. 354–363. [Google Scholar]
- Welzel, J.; Lankenau, E.; Birngruber, R.; Engelhardt, R. Optical coherence tomography of the human skin. J. Am. Acad. Dermatol. 1997, 37, 958–963. [Google Scholar] [CrossRef]
- Cao, T.; Tey, H.L. High-definition optical coherence tomography-an aid to clinical practice and research in dermatology. J. Dtsch. Dermatol. Ges. 2015, 13, 886–890. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, C.S.K.; Ortner, V.K.; Mogensen, M.; Philipsen, P.A.; Haedersdal, M. Transfollicular delivery of gold microparticles in healthy skin and acne vulgaris, assessed by in vivo reflectance confocal microscopy and optical coherence tomography. Lasers Surg. Med. 2019. [Google Scholar] [CrossRef]
- Xu, Q.; Jalilian, E.; Fakhoury, J.W.; Manwar, R.; Michniak-Kohn, B.; Elkin, K.B.; Avanaki, K. Monitoring the topical delivery of ultrasmall gold nanoparticles using optical coherence tomography. Skin Res. Technol. 2020, 26, 263–268. [Google Scholar] [CrossRef]
- Ghosn, M.G.; Sudheendran, N.; Wendt, M.; Glasser, A.; Tuchin, V.V.; Larin, K.V. Monitoring of glucose permeability in monkey skin in vivo using Optical Coherence Tomography. J. Biophotonics 2010, 3, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Shyu, V.B.; Chen, C.T. Dissolving Microneedle Patches for Transdermal Insulin Delivery in Diabetic Mice: Potential for Clinical Applications. Materials 2018, 11, 1625. [Google Scholar] [CrossRef] [Green Version]
- Carcamo-Martinez, A.; Anjani, Q.K.; Permana, A.D.; Cordeiro, A.S.; Larraneta, E.; Donnelly, R.F. Coated polymeric needles for rapid and deep intradermal delivery. Int. J. Pharm. 2020, 2, 100048. [Google Scholar] [CrossRef]
- Hermsmeier, M.; Sawant, T.; Chowdhury, K.; Nagavarapu, U.; Chan, K.F. First Use of Optical Coherence Tomography on In Vivo Inflammatory Acne-Like Lesions: A Murine Model. Lasers Surg. Med. 2020, 52, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Larsen, H.F.; Ahlstrom, M.G.; Gjerdrum, L.M.R.; Mogensen, M.; Ghathian, K.; Calum, H.; Sorensen, A.L.; Forman, J.L.; Vandeven, M.; Holerca, M.N.; et al. Noninvasive measurement of reepithelialization and microvascularity of suction-blister wounds with benchmarking to histology. Wound Repair Regen. 2017, 25, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Odorici, G.; Losi, A.; Ciardo, S.; Pellacani, G.; Conti, A. Non-invasive evaluation of Secukinumab efficacy in severe plaque psoriasis with confocal microscopy and optical coherence tomography: A case report. Skin Res. Technol. 2018, 24, 160–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, M.; Buehler, A.; Aguirre, J.; Ntziachristos, V. Three-dimensional multispectral optoacoustic mesoscopy reveals melanin and blood oxygenation in human skin in vivo. J. Biophotonics 2016, 9, 55–60. [Google Scholar] [CrossRef]
- Moustakidis, S.; Omar, M.; Aguirre, J.; Mohajerani, P.; Ntziachristos, V. Fully automated identification of skin morphology in raster-scan optoacoustic mesoscopy using artificial intelligence. Med. Phys. 2019, 46, 4046–4056. [Google Scholar] [CrossRef]
- Chuah, S.Y.; Attia, A.B.; Long, V.; Ho, C.J.; Malempati, P.; Fu, C.Y.; Ford, S.J.; Lee, J.S.; Tan, W.P.; Razansky, D.; et al. Structural and functional 3D mapping of skin tumours with non-invasive multispectral optoacoustic tomography. Skin Res. Technol. 2017, 23, 221–226. [Google Scholar] [CrossRef]
- Attia, A.B.E.; Chuah, S.Y.; Razansky, D.; Ho, C.J.H.; Malempati, P.; Dinish, U.S.; Bi, R.; Fu, C.Y.; Ford, S.J.; Lee, J.S.; et al. Noninvasive real-time characterization of non-melanoma skin cancers with handheld optoacoustic probes. Photoacoustics 2017, 7, 20–26. [Google Scholar] [CrossRef]
- Garza-Morales, R.; Rendon, B.E.; Malik, M.T.; Garza-Cabrales, J.E.; Aucouturier, A.; Bermudez-Humaran, L.G.; McMasters, K.M.; McNally, L.R.; Gomez-Gutierrez, J.G. Targeting Melanoma Hypoxia with the Food-Grade Lactic Acid Bacterium Lactococcus Lactis. Cancers 2020, 12, 438. [Google Scholar] [CrossRef] [Green Version]
- Ghazaryan, A.; Omar, M.; Tserevelakis, G.J.; Ntziachristos, V. Optoacoustic detection of tissue glycation. Biomed. Opt. Express 2015, 6, 3149–3156. [Google Scholar] [CrossRef] [Green Version]
- Anani, T.; Brannen, A.; Panizzi, P.; Duin, E.C.; David, A.E. Quantitative, real-time in vivo tracking of magnetic nanoparticles using multispectral optoacoustic tomography (MSOT) imaging. J. Pharm. Biomed. Anal. 2020, 178, 112951. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Dean-Ben, X.L.; Liu, N.; Gujrati, V.; Gottschalk, S.; Ntziachristos, V.; Razansky, D. Concurrent fluorescence and volumetric optoacoustic tomography of nanoagent perfusion and bio-distribution in solid tumors. Biomed. Opt. Express 2019, 10, 5093–5102. [Google Scholar] [CrossRef] [PubMed]
- Beziere, N.; Lozano, N.; Nunes, A.; Salichs, J.; Queiros, D.; Kostarelos, K.; Ntziachristos, V. Dynamic imaging of PEGylated indocyanine green (ICG) liposomes within the tumor microenvironment using multi-spectral optoacoustic tomography (MSOT). Biomaterials 2015, 37, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Sa, G.F.; Serpa, C.; Arnaut, L.G. Stratum corneum permeabilization with photoacoustic waves generated by piezophotonic materials. J. Control. Release 2013, 167, 290–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristina, F.; Vieira, L.; Badra Bentley, M.V.L. Confocal Laser Scanning Microscopy as a Tool for the Investigation of Skin Drug Delivery Systems and Diagnosis of Skin Disorders. In Confocal Laser Microscopy-Principles and Applications in Medicine, Biology, and the Food Sciences; Lagali, N., Ed.; Intech: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Roman, R.; Naik, A.; Kalia, Y.N.; Fessi, H.; Guy, R.H. Visualization of skin penetration using confocal laser scanning microscopy. Eur. J. Pharm. Biopharm. 2004, 58, 301–316. [Google Scholar] [CrossRef]
- Huzaira, M.; Rius, F.; Rajadhyaksha, M.; Anderson, R.R.; Gonzalez, S. Topographic variations in normal skin, as viewed by in vivo reflectance confocal microscopy. J. Investig. Dermatol. 2001, 116, 846–852. [Google Scholar] [CrossRef] [Green Version]
- Denk, W.; Strickler, J.H.; Webb, W.W. Two-photon laser scanning fluorescence microscopy. Science 1990, 248, 73–76. [Google Scholar] [CrossRef] [Green Version]
- Yew, E.; Rowlands, C.; So, P.T. Application of Multiphoton Microscopy in Dermatological Studies: A Mini-Review. J. Innov. Opt. Health Sci. 2014, 7, 1330010. [Google Scholar] [CrossRef]
- Patalay, R.; Talbot, C.; Alexandrov, Y.; Lenz, M.O.; Kumar, S.; Warren, S.; Munro, I.; Neil, M.A.; Konig, K.; French, P.M.; et al. Multiphoton multispectral fluorescence lifetime tomography for the evaluation of basal cell carcinomas. PLoS ONE 2012, 7, e43460. [Google Scholar] [CrossRef] [Green Version]
- Obeidy, P.; Tong, P.L.; Weninger, W. Research Techniques Made Simple: Two-Photon Intravital Imaging of the Skin. J. Investig. Dermatol 2018, 138, 720–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Digabel, J.; Houriez-Gombaud-Saintonge, S.; Filiol, J.; Lauze, C.; Josse, G. Dermal fiber structures and photoaging. J. Biomed. Opt. 2018, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Puschmann, S.; Rahn, C.D.; Wenck, H.; Gallinat, S.; Fischer, F. Approach to quantify human dermal skin aging using multiphoton laser scanning microscopy. J. Biomed. Opt. 2012, 17, 036005. [Google Scholar] [CrossRef] [PubMed]
- Fischer, F.; Volkmer, B.; Puschmann, S.; Greinert, R.; Breitbart, E.; Kiefer, J.; Wepf, R. Assessing the risk of skin damage due to femtosecond laser irradiation. J. Biophotonics 2008, 1, 470–477. [Google Scholar] [CrossRef]
- Kiss, N.; Fesus, L.; Bozsanyi, S.; Szeri, F.; Van Gils, M.; Szabo, V.; Nagy, A.I.; Hidvegi, B.; Szipocs, R.; Martin, L.; et al. Nonlinear optical microscopy is a novel tool for the analysis of cutaneous alterations in pseudoxanthoma elasticum. Lasers Med. Sci. 2020. [Google Scholar] [CrossRef]
- Springer, S.; Zieger, M.; Hipler, U.C.; Lademann, J.; Albrecht, V.; Bueckle, R.; Mess, C.; Kaatz, M.; Huck, V. Multiphotonic staging of chronic wounds and evaluation of sterile, optical transparent bacterial nanocellulose covering: A diagnostic window into human skin. Skin Res. Technol. 2019, 25, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.L.; Hu, P.S.; Ghazaryan, A.; Chen, S.J.; Tsai, T.H.; Dong, C.Y. Quantitative analysis of multiphoton excitation autofluorescence and second harmonic generation imaging for medical diagnosis. Comput. Med. Imaging Graph. 2012, 36, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Sugata, K.; Osanai, O.; Sano, T.; Takema, Y. Evaluation of photoaging in facial skin by multiphoton laser scanning microscopy. Skin Res. Technol. 2011, 17, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Pena, A.M.; Chen, X.; Pence, I.J.; Bornschlogl, T.; Jeong, S.; Gregoire, S.; Luengo, G.S.; Hallegot, P.; Obeidy, P.; Feizpour, A.; et al. Imaging and quantifying drug delivery in skin-Part 2: Fluorescence andvibrational spectroscopic imaging methods. Adv. Drug Deliv. Rev. 2020. [Google Scholar] [CrossRef]
- Becker, W.; Bergmann, A.; Hink, M.A.; Konig, K.; Benndorf, K.; Biskup, C. Fluorescence lifetime imaging by time-correlated single-photon counting. Microsc. Res. Tech. 2004, 63, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R.; Szmacinski, H.; Nowaczyk, K.; Johnson, M.L. Fluorescence lifetime imaging of free and protein-bound NADH. Proc. Natl. Acad. Sci. USA 1992, 89, 1271–1275. [Google Scholar] [CrossRef] [Green Version]
- Krasieva, T.B.; Stringari, C.; Liu, F.; Sun, C.H.; Kong, Y.; Balu, M.; Meyskens, F.L.; Gratton, E.; Tromberg, B.J. Two-photon excited fluorescence lifetime imaging and spectroscopy of melanins in vitro and in vivo. J. Biomed. Opt. 2013, 18, 31107. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.; Greenfield, D.A.; Hermsmeier, M.; Yamamoto, A.; Chen, X.; Chan, K.F.; Evans, C.L. Time-resolved fluorescence microscopy with phasor analysis for visualizing multicomponent topical drug distribution within human skin. Sci. Rep. 2020, 10, 5360. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, P.M.; Kalinina, S.; Rueck, A.; von Arnim, C.A.F.; von Einem, B. NADH Autofluorescence-A Marker on its Way to Boost Bioenergetic Research. Cytometry A 2019, 95, 34–46. [Google Scholar] [CrossRef] [Green Version]
- Huck, V.; Gorzelanny, C.; Thomas, K.; Getova, V.; Niemeyer, V.; Zens, K.; Unnerstall, T.R.; Feger, J.S.; Fallah, M.A.; Metze, D.; et al. From morphology to biochemical state-intravital multiphoton fluorescence lifetime imaging of inflamed human skin. Sci. Rep. 2016, 6, 22789. [Google Scholar] [CrossRef]
- Seidenari, S.; Arginelli, F.; Bassoli, S.; Cautela, J.; French, P.M.; Guanti, M.; Guardoli, D.; Konig, K.; Talbot, C.; Dunsby, C. Multiphoton laser microscopy and fluorescence lifetime imaging for the evaluation of the skin. Dermatol. Res. Pract. 2012, 2012, 810749. [Google Scholar] [CrossRef]
- Ogawa-Fuse, C.; Morisaki, N.; Shima, K.; Hotta, M.; Sugata, K.; Ichihashi, T.; Oguri, M.; Yoshida, O.; Fujimura, T. Impact of water exposure on skin barrier permeability and ultrastructure. Contact Dermat. 2019, 80, 228–233. [Google Scholar] [CrossRef]
- Lademann, J.; Meinke, M.C.; Schanzer, S.; Richter, H.; Darvin, M.E.; Haag, S.F.; Fluhr, J.W.; Weigmann, H.J.; Sterry, W.; Patzelt, A. In vivo methods for the analysis of the penetration of topically applied substances in and through the skin barrier. Int. J. Cosmet. Sci. 2012, 34, 551–559. [Google Scholar] [CrossRef]
- Mohammed, Y.H.; Holmes, A.; Haridass, I.N.; Sanchez, W.Y.; Studier, H.; Grice, J.E.; Benson, H.A.E.; Roberts, M.S. Support for the Safe Use of Zinc Oxide Nanoparticle Sunscreens: Lack of Skin Penetration or Cellular Toxicity after Repeated Application in Volunteers. J. Investig. Dermatol. 2019, 139, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Brodwolf, R.; Volz-Rakebrand, P.; Stellmacher, J.; Wolff, C.; Unbehauen, M.; Haag, R.; Schafer-Korting, M.; Zoschke, C.; Alexiev, U. Faster, sharper, more precise: Automated Cluster-FLIM in preclinical testing directly identifies the intracellular fate of theranostics in live cells and tissue. Theranostics 2020, 10, 6322–6336. [Google Scholar] [CrossRef]
- Wanjiku, B.; Yamamoto, K.; Klossek, A.; Schumacher, F.; Pischon, H.; Mundhenk, L.; Rancan, F.; Judd, M.M.; Ahmed, M.; Zoschke, C.; et al. Qualifying X-ray and Stimulated Raman Spectromicroscopy for Mapping Cutaneous Drug Penetration. Anal. Chem. 2019, 91, 7208–7214. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Can Distinguish between Interfollicular Epidermis, Hair-Follicles and Glands | Can Distinguish between Barriers in SC, Viable Epidermis, Dermis | Can Specifically Address Tjs in the Viable Epidermis/Hair Follicles/Glands | Measurement of Inside-Out or Outside-In Flux? | Flux Can Be Quantified? | Measurement at Different Time Points Possible [Permeation-Penetration Kinetics]? | Human In Vivo 1 Application Possible? |
|---|---|---|---|---|---|---|---|
| TEWL | − | − | − | Inside-out | + | ++ | ++ |
| TEER | − | − | − | No directionality | ++ | ++ | − |
| Advanced electrical impedance spectroscopy | − | + | − | No directionality | ++ | + | (+) |
| (Radio)Chemical analyses of drugs/tracers applied onto the skin in acceptor compartment/blood (e.g., by UPLC, UHPLC-MS/MS etc.) | − | − | − | Outside-in | +++ | ++ | + |
| Chemical analysis of drugs/tracers applied onto the skin in different skin layers after separation 2 and subsequent extraction (analyses e.g., by UPLC, UHPLC-MS/MS etc.) | − | (+) (with certain limitations) | − | Outside-in | +++ | − [++] 3 | − |
| Dye permeation analysis | − | − | − | Outside-in | ++ | ++ | − |
| Raman Spectroscopy/microscopy | + | + 4/++ 5 | − | Outside-in and inside-out 6 | + | ++ | ++ |
| (ATR-)FTIR spectroscopy | (+) 7 | (+) 7 | − | Outside-in and inside-out 6 | + | ++ | ++ |
| Transmission electron microscopy combined with tracers | +++ | +++ | +++ | Outside-in and inside-out 6 | − | − | − |
| Immuno-histochemistry combined with tracers (e.g., biotinylation assay) (Fluorescence microscopy or CLSM) | ++ | ++ | ++ | Outside-in and inside-out 6 | (+) | − | − |
| OCT/optoacoustic imaging | + | + | − | Outside-in and inside-out 6 | + | ++ | ++ |
| In vivo CLSM/reflectance confocal microscopy (RCM) | + (HF) − (Glands) | − | − | Outside-in and inside-out 6 | − | ++ | + |
| MPM/FLIM | + | ++ | (−) except for intrinsic fluorescent TJs | Outside-in and inside-out 6 | (+) | ++ | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorzelanny, C.; Mess, C.; Schneider, S.W.; Huck, V.; Brandner, J.M. Skin Barriers in Dermal Drug Delivery: Which Barriers Have to Be Overcome and How Can We Measure Them? Pharmaceutics 2020, 12, 684. https://doi.org/10.3390/pharmaceutics12070684
Gorzelanny C, Mess C, Schneider SW, Huck V, Brandner JM. Skin Barriers in Dermal Drug Delivery: Which Barriers Have to Be Overcome and How Can We Measure Them? Pharmaceutics. 2020; 12(7):684. https://doi.org/10.3390/pharmaceutics12070684
Chicago/Turabian StyleGorzelanny, Christian, Christian Mess, Stefan W. Schneider, Volker Huck, and Johanna M. Brandner. 2020. "Skin Barriers in Dermal Drug Delivery: Which Barriers Have to Be Overcome and How Can We Measure Them?" Pharmaceutics 12, no. 7: 684. https://doi.org/10.3390/pharmaceutics12070684
APA StyleGorzelanny, C., Mess, C., Schneider, S. W., Huck, V., & Brandner, J. M. (2020). Skin Barriers in Dermal Drug Delivery: Which Barriers Have to Be Overcome and How Can We Measure Them? Pharmaceutics, 12(7), 684. https://doi.org/10.3390/pharmaceutics12070684